










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.4 Pretoria Apr. 2008
SHORT COMMUNICATION
Physical impact of grazing by sheep in the Nama Karoo subshrub/grass rangeland of South Africa on litter and dung distribution
G. van N. du Toit*; H.A. Snyman; P.J. Malan
Animal, Wildlife & Grassland Sciences, University of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa
ABSTRACT
The direct short-term impact of three rates of stocking (4, 8 and 16 Small Stock Units/ha) on arid Nama Karoo vegetation (shrub/grass) was quantified in terms of dung distribution and trampling. Grazing, by mature Merino wethers took place for one month at a time over a period of two years (1995 and 1996). As expected, the quantity of litter and dung significantly increased with an increase in stocking rate. When litter and dung are expressed in terms of loss and excretion per animal, both decreased with increased stocking rate. On average 80% of the dung was excreted on only 10% of the camp area, which mostly took place at the watering and sleeping places. These findings demonstrate the complex interaction between grazing and soil parameters.
Keywords: Karoo bushes, Merino wethers, soil parameters, stocking rate, trampling
Rangeland management focused more on the effects of various management practices on forage production and animal responses but less attention to the impact of grazing on the nutrient dynamics of soils (Snyman, 1998; Bisigato et al., 2008). Grazing has been reported to accelerate nutrient cycling in African savannas (McNaughton et al., 1988), North American semi-arid grasslands (Shariff et al., 1994) and Patagonian Monte shrublands of Argentina (Bisigato et al., 2008). No comparable studies have been undertaken on Karoo subshrub/grass vegetation of South Africa; however Whitford (1986) suggested that sheep grazing might tie up organic matter in resistant pellets, reducing rates of nutrient return to the soil in arid ecosystems. This is supported by findings of Milton & Dean (1996) in the South African Nama Karoo vegetation, that dung pellets of sheep and springbok disintegrated slowly, particularly on the soil surface.
Assessment of the spatial and temporal distributions of herbivores on the basis of dung counts is also based on assumptions about the rates of dung decay (Wigley & Johnson, 1981). Despite dung applications to rangeland conservation and rehabilitation, ecosystem processes, including decomposition and nutrient cycling, have as yet received little attention in arid South African ecosystems (Milton & Dean, 1996). Information on site-specific decay rates for dung and other plant materials has applications in rangeland management for sustainable functioning of the ecosystem (McAuliffe, 1988). In general, most studies do not indicate any single or consistent response of organic matter to grazing. It is clear that many factors determine the response of soil organic matter to livestock grazing (Manley et al., 1995). This study therefore aimed at the quantification of the short-term direct impact of different sheep stocking rates on litter and dung distribution of arid subshrub/grass vegetation.
The research was conducted on the Research Station of the Grootfontein Agricultural Development Institute, 10 km north-east of Middelburg, Eastern Cape Province (25° 06' E; 31° 40' S, altitude 1 400 m), which is situated in the arid region of South Africa. Rain falls almost exclusively during spring and autumn, with a mean annual average of 366 mm. The study area is subject to temperature extremes. Mean monthly maximum and minimum temperatures range from 38 °C in January to -12 °C in June, with a mean of 150 frost days per annum (Schulze, 1979).
The study site was situated in the Eastern Mixed Nama Karoo (Nama Karoo biome) (Hoffman, 1996). A complex mix of grass- and subshrub-dominated vegetation types, which is subject to dynamic changes in species composition dependent on seasonal rainfall events, occurs within this vegetation type. Common shrubs or Karoo bushes include Pentzia incana, Eriocephalus ericoides and Hermannia spp., while grasses, such as Aristida spp., Eragrostis spp. and Themeda triandra may dominate the landscape after good summer rains. This vegetation type has the highest cover of herbs of all the Nama Karoo types, as well as numerous geophytes (Hoffman, 1996). Beaufort Group sandstones and shales dominate the landscape with the flat-topped landscape shaped by many dolerite dykes and sills (Hoffman, 1996). Soils in the study area are mostly clay loam soils of the Valsrivier Form (Zuiderzee series) (Soil Classification Working Group, 1991). The depth of the A horizon varies from 90 to 120 mm.
The research was carried out on eight camps of one ha each (108 x 92 m) with a western aspect and a 1% slope. The camps were situated in two blocks of four, adjacent to each other. The four treatments consisted of three different stocking rates and a control. The stocking rates consisted of 4, 8 and 16 Small Stock Units (SSU)/ha. Two grazing periods of one month each were applied. The long-term grazing capacity of the study area is 2.6 ha/SSU/year or 0.39 SSU/ha/year, which is equal to a stocking rate of 4.6 SSU/ha/month (Du Toit, 2001). Mature Merino wethers were used as trial animals. The body mass of the animals, randomly allocated to camps, varied between 49.4 and 51.5 kg. The first grazing period lasted from 1 May 1995 to 30 May 1995 and the second from 1 May 1996 to 30 May 1996. This period forms the end of the seasonal growth activity for both grasses and Karoo bushes (herbaceous layer). The objective of the study was not to look at weight changes and therefore the animals were not weighed at the end of each season or grazing period. For the season preceding the trial, no grazing was applied in the camps.
Trampling losses were determined by gathering all litter, except for dung, in nine plots of 1 m2, randomly distributed in a camp. Only material not attached to plants was collected. The material was dried at 80 °C for 24 hours before determining mass. The results for every camp were expressed in both gram per m2 and gram per animal. The distribution of the droppings was determined by gathering all the droppings in the same nine plots used for trampling loss determinations. These were then dried for 24 hours at 80 °C and expressed in gram per m2 and gram per animal for each camp. The trampling losses of plants (litter) and dung excretion were carried out only after each grazing period was applied. This was done due to the fact that there was obviously no pre-grazing data available for trampling and dung excretion before the grazing trial started.
The experimental layout consisted of split-plots with four treatments and two replications. The treatments and repetitions were randomly allocated. All data were analyzed using a one-way analysis of variance technique (Mendenhall & Sincich, 1996). The programme of BMDP 2 V.9 (Dixon, 1985), set up for analysis of split-plots, was used. Data from different sampling periods and/or years were analysed separately. The trampling losses of plants (litter) and dung excretions were only analysed for stocking rate effects and not before and after grazing. The reason for this being that no preparation data was available. Tukey's test was used to determine the differences (Mendenhall & Sincich, 1996).
Litter production within a stocking density differed non-significantly (P >0.05) between the two seasons or grazing periods. The mean litter production for stocking rates 0, 4, 8 and 16 SSU/ha, over the two seasons, was 99.6, 139.3, 209.7 and 353.4 kg/ha (F332 = 6.14, P <0.01), respectively. Litter production in the 0 SSU/ha stocking density can be ascribed to the normal dying and decomposing of plant material. When litter is expressed in terms of loss per animal, the loss decreased significantly (F332 = 5.34, P <0.01) for the different stocking rates, from 179.2, 31.3, 23.6 and 19.9 g/animal for no grazing, 4, 8 and 16 SSU/ha, respectively.
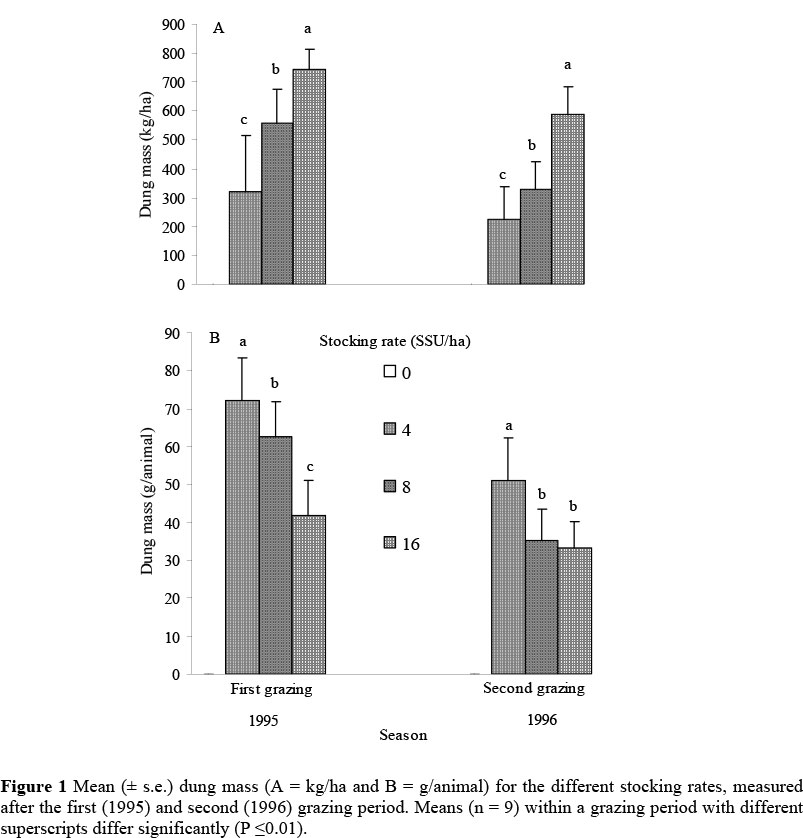
As expected, in both seasons dung excretion increased (F332 = 5.55, P <0.01) with increased stocking density (Figure 1). Interestingly, in all stocking rates there was a considerably lower dung excretion with the second grazing (1996). For the 1996 season for example, the dung excretion for the 8 SSU stocking rate was 41% lower than in the 1995 season. The reason for this is unclear. When dung excretion is expressed as dung excretion per animal, it shows a decrease with increased stocking rate. An important aspect was that about 80% of the dung was excreted on 10% of the camp area. Dung excretion took place at the watering and sleeping places.
Most researchers are concerned about the effects of heavy grazing on soil quality and on sustainability of rangeland resources (Manley et al., 1995; Snyman & Du Preez, 2005; Bisigato et al., 2008). The litter and dung excretion results as obtained in this study indicate that a low stocking rate has the least detrimental effect on vegetation over the long-term. This does not mean that a high stocking rate should be avoided. A high stocking rate can play an important role in renewal of the vegetation and an increase in production over the short-term (Flemmer et al., 2002; 2003). On the other hand, high intensity and/or early season grazing have been found to have a greater negative impact on litter and live plant biomass than light intensity grazing of mixed grass prairie of North Dakota (Shariff et al., 1994) and/or late season grazing of mixed prairie pastures in Canada (Neath et al., 1991).
Considering the soil aspects in this study, trampling was too uneven to really be of value. It was especially localised at watering points and sleeping places. In larger camps, trampling may be even more uneven. If the expenses incurred to fence off smaller camps are compared to the few advantages accompanying high stocking densities, it is probably not feasible making these camps. On the same Karoo vegetation as in this study, Beukes et al. (2002) showed that 60 camps or more allow time for forage reserves to build up, but that more than 150 camps become too costly. Due to the extensive nature of Karoo subshrub/grass vegetation and its sparse cover, it is questionable whether a greater percentage litter on soil with a higher stocking rate might have any real long-term benefits. This must also be compared to the fact that those areas with the highest percentage litter could also be those where most soil compaction took place (Snyman & Du Preez, 2005). Furthermore, high stocking rates lead to a greater decrease in canopy and canopy-spread cover, which is detrimental for the vegetation in total (O'Connor & Roux, 1995). According to Milton & Dean (1996) the dead shrubs lying on the soil surface in an arid shrub land in the southern Karoo have half-lives of nine to 18 years depending on wood density which varies among species.
The increase in litter as found in this study with greater stocking rates, can well be beneficial to the functioning of the rangeland ecosystem over the short-term by better covering the soil, lowering the soil temperatures (Snyman & Du Preez, 2005) and increasing infiltration (Snyman, 2005). Also, many studies reported an increase in soil nutrients and C in grazed plots which have usually been attributed to litter inputs with reduced C : N ratios (Whitehead, 1995), increased litter turnover and incorporation via trampling and redistribution of nutrients by grazing and excretion (Dormaar et al., 1997). In contrast, Thurow et al. (1988) reported that on semi-arid rangelands, high stocking rates resulted in less litter accumulation than in low stocking rates. According to Bisigato et al. (2008) the conservation of soil resources is not linearly related to grazing. Unfortunately, the C and N content of the soil were not determined in our study.
The benefits of dung and urine added to soil surfaces have been known for millennia (Wood et al., 2008). It is apparent from the fact that about 80% of the dung is excreted on 10% of the camp area and that dung excretions are very localised even in small camps, which is in agreement with the findings of Marten & Donker (1966). Though not measured, the decrease in dung excretion with increase in stocking rate can possibly be ascribed to a decrease in feed intake due to higher competition between the animals for the available food. If this hypothesis is correct, it would lead to poorer animal production; but unfortunately no weight changes of the animals were recorded in this study. According to Milton & Dean (1996), dung pellets of sheep can remain intact on the soil surface in Karoo subshrub/grass vegetation for five years or more.
The study clearly showed that the quantity of litter and dung on the soil is mainly determined by the level of grazing intensity. As expected, the dung quantity increased with higher stocking rates. Unfortunately no direct conclusions on the effects of management and weather on nutrient cycling in these dry rangelands could be made from these results. In future the relation between rates of physical disintegration of organic matter and nutrient release must be determined for a clearer understanding of the functioning of the ecosystem.
References
Beukes, P.C., Cowling, R.M. & Higgins, S.I., 2002. An ecological economic simulation model of a nonselective grazing system in the Nama Karoo, South Africa. Ecol. Econ. 42, 221-242. [ Links ]
Bisigato, A.J., Laphitz, R.M.L. & Carrera, A.L., 2008. Non-linear relationships between grazing pressure and conservation of soil resources in Patagonian Monte shrublands. J. Arid Environ. 72, 1464-1475. [ Links ]
Dixon, W.J., 1985. BMDP Statistical Software Manual. University of California Press. Berkley, Los Angeles, London. [ Links ]
Dormaar, J.F., Adams, B.W. & Williams, W.D., 1997. Impacts of rotational grazing on mixed prairies' soil and vegetation. J. Range Mgmt. 50, 647-651. [ Links ]
Du Toit, P.C.V., 2001. A study of animal production under different grazing regimes in the False Upper Karoo at Grootfontein. Afr. J. Range For. Sci. 18, 137-141. [ Links ]
Flemmer, A.C., Busso, C.A. & Fernandez, O.A., 2002. Bud viability in perennial grasses: Water stress and defoliation effects. J. Range Mgmt. 55, 150-163. [ Links ]
Flemmer, A.C. Busso, C.A., Fernandez, O.A. & Montani, T., 2003. Effects of defoliation at varying soil water regimes on aboveground biomass of perennial grasses. Arid Land Res. Mgmt. 17, 139-152. [ Links ]
Hoffman, M.T., 1996. Eastern Mixed Nama Karoo. In: Vegetation of South Africa, Lesotho and Swaziland. Eds Low, A.B. & Rebelo, A.G., Department of Environmental Affairs and Tourism, Pretoria. pp. 85. [ Links ]
Manley, J.T., Schuman, G.E., Reeder, J.D. & Hart, R.H., 1995. Rangeland soil carbon and nitrogen responses to grazing. J. Soil Water Conservation 50, 294-298. [ Links ]
Marten, G.C. & Donker, J.D., 1966. Animal excrement as a factor influencing acceptability of grazed forage. Proceedings 10th International Grassland Congress, Helsinki. pp. 359-363. [ Links ]
McAuliffe, J.R., 1988. Markovian dynamics of simple and complex desert plant communities. The American Naturalist 131, 459-490. [ Links ]
McNaughton, S.J., Ruess, R.W. & Seagle, S.W., 1988. Large mammals and process dynamics in African ecosystems. BioScience 38, 794-800. [ Links ]
Mendenhall, W. & Sincich, T., 1996. Principles of Experimental Design. A second course in Statistics. 5th Ed. Prentice-Hall Inc., New Jersey. pp. 582-595. [ Links ]
Milton, S.J. & Dean, W.R.J., 1996. Rates of wood and dung disintegration in arid South African rangelands. Afr. J. Range Forage Sci. 13, 89-93. [ Links ]
Neath, M.A. , Baily, A.W. , Pluth, D.J, Chanask, D.S. & Hardon, R.T., 1991. Grazing impacts on litter and soil organic matter in mixed prairie and fescue grassland ecosystems in Alberta. J. Range Mgmt. 44, 7-12. [ Links ]
O'Connor, T.G. & Roux, P.W., 1995. Vegetation changes (1949-71) in a semi-arid, grassy dwarf shrubland in the Karoo, South Africa: Influence of rainfall variability and grazing by sheep. J. Appl. Ecol. 32, 612-626. [ Links ]
Schulze, E.R., 1979. Climate of South Africa. Part 8. General Survey, Weather Bureay, Pretoria. 158 pp. [ Links ]
Shariff, A.R., Biondini, M.E. & Grygiel, C.E., 1994. Grazing intensity effects on litter decomposition and soil nitrogen mineralization. J. Range Mgmt. 47, 444-449. [ Links ]
Snyman, H.A., 1998. Dynamics and sustainable utilization of the rangeland ecosystem in arid and semi-arid climates of southern Africa. J. Arid Environ. 39, 645-666. [ Links ]
Snyman, H.A., 2005. Rangeland degradation in a semi-arid South Africa, I: influence on seasonal root distribution, root/shoot ratios and water-use efficiency. J. Arid Environ. 60, 457-481. [ Links ]
Snyman, H.A. & Du Preez, C.C., 2005. Rangeland degradation in a semi-arid South Africa, II: influence on soil quality. J. Arid Environ. 60, 483-507. [ Links ]
Soil Classification Working Group, 1991. Soil Classification: A Taxonomic System for South Africa. Department of Agriculture Development, Pretoria South Africa. 262 pp. [ Links ]
Thurow, T.L., Blackburn, W.H. & Taylor, C.A., 1988. Some vegetation responses to selected livestock grazing strategies. J. Range Mgmt. 41, 108-114. [ Links ]
Whitehead, D.C., 1995. Grassland Nitrogen. CAB International, Wallingford. pp. 397. [ Links ]
Whitford, W.G., 1986. Decomposition and nutrient cycling in deserts. In: Pattern and process in desert ecosystems. Ed. Whitford, W.G., University of New Mexico Press. New Mexico. pp. 93-117. [ Links ]
Wigley, T.B. & Johnson, M.K., 1981. Disappearance rate for deer pellets in the southeast. J. Wildlife Mgmt. 45,251-253. [ Links ]
Wood, K., Rubio, H. & Wood, C., 2008. Rangeland Management and Hydrology. Proc. XXI International Grassland Congress and VII International Rangeland Congress, Hohhot, China. 1, 809-812. [ Links ]
 Correspondence:
Correspondence:
E-mail: snymanha.sci@ufs.ac.za
* Present address: P.O. Box 94, Hanover 7005, South Africa


{kind=link}