










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 no.4 Pretoria Abr. 2008
Carcass characteristics and meat quality of progeny of five merino dam lines, crossed with dormer and suffolk sires
J.J.E. CloeteI, II; L.C. HoffmanI; S.W.P. CloeteI, III
IDepartment of Animal Sciences, University of Stellenbosch, Private Bag XI, Matieland 7602, South Africa
IICape Institute for Agricultural Training: Elsenburg, Private Bag X1, Elsenburg 7607, South Africa
IIIInstitute for Animal Production: Elsenburg, Private Bag X1, Elsenburg 7607, South Africa
ABSTRACT
This study details the slaughter traits of lambs born from a terminal crossbreeding experiment that involved five Merino type dam lines crossed with Dormer and Suffolk sires. Dam lines included dual purpose types; South African Mutton Merino (SAMM), Dohne Merino, SAMM rams crossed to commercial Merino ewes (SAMM cross) and specialist Merino lines selected either for clean fleece weight (FW+) or for an increased fertility (Rep+). Data include between 228 and 483 individual records, depending on the trait. The unadjusted fat depth 25 mm from the midline at the 13th rib of lambs from dual-purpose ewes was between 22 and 32% greater than that of lambs from FW+ Merino ewes. Progeny from Rep+ Merino ewes showed a closer resemblance to the progeny of dual-purpose breeds than to those of FW+ Merino ewes in this instance. Adjustment for slaughter weight eliminated these differences. The initial pH of meat from progeny of FW+ Merino ewes was lower than that from progeny of dual-purpose ewes, and Rep+ Merino ewes. No differences in Warner-Bratzler shear values of the meat were found between the different crosses. Crossbred progeny of the Merino lines performed satisfactorily for all the traits considered, and will not be discriminated against in the market. No conclusive differences in favour of either sire breed were found.
Keywords: Dohne Merino, meat quality, SA Mutton Merino, terminal crossbreeding
Introduction
At the present wool : meat price ratio, most of the financial income acquired in a sheep enterprise is derived from meat, and particularly from the marketing of lambs. Therefore, many strategies for the improvement of lamb production and the growth performance of the lambs have been implemented. One such strategy is the production of commercial crossbred slaughter lambs. The crossbred lambs should have a higher birth weight, and better growth performance (relative to the mean of the parent breed) up to the age of weaning and should reach slaughter weight in an intensive fattening system faster (Teixeira et al., 1996; Doloksaribu et al., 2000; Freking et al, 2000; Snowder & Duckett, 2003; Fogarty, 2006).
Crossbreeding is a powerful tool for the animal breeder and is of critical importance to the meat sheep industry. Crossbred individuals are expected to exhibit heterosis. The percentage increase in performance relative to the mid-parent value of the pure breeds is given as 10% for lamb survival, while it ranges from 3 - 10% for growth traits (Fogarty, 2006). A good crossbreeding system aims to use breeding ewes that are small as well as prolific (Piper & Ruvinsky, 1997).
However, within the South African small stock industry, very little research has been conducted on crossbreeding systems utilizing local breeds. The objective of this study was therefore to compare the carcass composition and meat quality of the progeny of Dormer and Suffolk sires. The Dormer breed was originally developed to provide a terminal crossbreeding sire breed for use on Merino ewes in the Western Cape (De Villiers & Cloete, 1984). The Suffolk sire is used widely in the world for heavy slaughter lamb production (Milton et al., 1994). Ellis et al. (1997) found that Suffolk sired lambs were heavier, but with similar carcass weights and killing-out proportions compared to Charollais and Texel rams. The rams were crossed with five dam lines (Cloete et al., 2004b). The dam lines were South African Mutton Merino (SAMM), a first cross ewe line called SAMM cross (SAMM rams crossed to commercial Merino ewes), Dohne Merino, as well as two specialist Merino lines. The first of these was selected for clean fleece weight (FW+) and the second was selected for an increased reproduction (Rep+). Both lines originated from commercial Merino stock.
The carcass characteristics and measurements of carcass quality of progeny of the respective ewe lines mated to either Dormer or Suffolk rams were compared in this paper.
Materials and Methods
The experiment was conducted at Langgewens experimental farm in the Swartland region of South Africa from 1999 to 2002. The climate at the site is Mediterranean, with 78% of the total long-term annual precipitation of 395 mm being recorded during winter (April to September). Merino type ewes, belonging to five distinct lines, represented the female breeding animals. These ewes were born during 1997 and 1998, and were not selected as replacements for breeding purpose in their flocks of origin. Approximately 20 ewes within each birth year groups (thus 40 ewes in total) represented each line. The maternal lines were described in detail by Cloete et al. (2003; 2004b). Dormer or Suffolk rams were used as terminal sire breeds on the ewe flock. Two rams of each sire breed were used in single sire groups during 1999, when only the ewes born in 1997 were available. Three rams of each breed were used in 2000 and 2001, while only two rams were used in 2002. In each year, one ram of each breed was retained for breeding, to provide sire links across years. In total, eight rams were used per sire breed.
The ewes lambed during June and July. All the lambs were slaughtered at a target live weight of 40 kg (Cloete et al., 2004b/ Full body live weight was determined 24 h prior to slaughtering (Hopkins et al., 1996). The sheep were slaughtered at a commercial abattoir using standard South African slaughter techniques, which included electrical stunning (4 sec at 200 Volts) before being exsanguinated. The carcasses were subsequently hung to bleed out before they were skinned. The carcasses were kept in a cooler at 4 °C for 24 h, after which the dressed carcasses were transported (carcasses were transported in a refrigerated truck, and the duration of the trip was approximately 45 min) from the abattoir to a deboning facility. Here they were kept for another 24 h at 4 °C, prior to deboning and sampling.
Carcass components that were weighed included the head, trotters and skin. Measurements on the carcass included the measurement of fat depth at two sites. The first site was for fat measurements at the 13th rib 25 mm from the midline and the second site between the 3rd and 4th lumbar vertebrae 25 mm from the midline on the left side of the carcass (Bruwer et al., 1987). The carcass length was measured on the hanging carcass from the Pubis ischium to the cranial edge of the first rib. The hindleg circumference was measured at the maximum circumference of a line passing over the distal end of the iliac wings of the pelvis and the most caudal point on the median line between the legs (Stanford et al., 1997). Hindleg length was measured from the inner edge of the proximal end of the tibia to the anterior tip of the pubis (Enright et al., 1990). The carcass depth was measured at the first rib cranial of the sternum, and the carcass width was taken at the widest place of the carcass at the forequarter.
After 48 h in cold storage (4 °C) the carcasses were weighed to determine cold carcass weight before they were partitioned into South African retail cuts, which were weighed separately (Cloete et al., 2004a). These cuts consisted of the neck, shoulder, chuck, flatrib, primerib, loin and hindquarters. The neck was removed at the 7th cervical vertebrae (the point where the neck starts bending), the cut being made at right angles to the spine. Thereafter the hind legs were removed. This consisted of loosening the flanks on the inside of the legs (following the curve of the leg muscle) to an imaginary line perpendicular to the ilium (seen from the inside of the carcass). The leg was then removed by cutting along this line, just missing the ilium (through the last lumbar vertebrae). The rest of the carcass was then halved along the spinal column, prior to being separated into retail cuts (shoulder, chuck, flatrib, primerib and loin). The shoulder was removed by sawing along an imaginary line from the elbow joint to a point below the spinal column, between the 5th and 6th ribs. The carcass was then swivelled so that the spinal column was sawn through at right angles. The flank was removed by sawing from the obliquus abdominis internus muscle parallel to the spine. The loin and rib were separated perpendicularly to the spinal column at the junction of the thoracic and lumbar vertebrae. All commercial cuts were weighed to the nearest gram without any trimming of fat.
Instrumental measurements of meat quality were made on the M. longissimus lumborum. A CRISON pH meter (507) fitted with a CRISON electrode (Cat. 52-32) was inserted directly into the meat, to measure pH 45 min, 24 h and 48 h post mortem. The pH meter was equipped with a thermometer that allowed automatic adjustment for temperature, and was calibrated with standard buffers at pH 4.0 and pH 7.0. Recalibration was done after every 10 animals measured. The pH was measured on the right side of each animal in the M. longissimus between the 1st and the 2nd lumbar vertebrae.
The longissimus lumborum muscle from the left side was dissected from the 1st to the 6th lumbar vertebrae and used for meat quality analyses (Schönfeldt et al., 1993). Two loin 1.5 cm sub-samples of approximately 35 g (the first taken at the first lumbar vertebrae and the second, caudally adjacent to it) were taken for the determination of cooking loss, drip loss, colour and meat tenderness (Honikel, 1998). For drip loss determination one of the loin sub-samples was weighed, placed in netting and suspended in an inflated plastic bag. After a storage period of 24 h at 4 °C, the samples were weighed again after removal from the bags, and the drip loss was calculated as weight loss expressed as a percentage of the original weight of the sample (Honikel, 1998).
Colour was evaluated by using a Color-guide 45°/0° colorimeter (BYK-Gardner, USA) to determine L*, a*, b*, chroma and hue angle (Commission International de l'Eclairage, 1976), with L* indicating brightness, a* the red-green range, and b* the blue-yellow range. The Color-guide 45°/0° colorimeter was calibrated using the green, black and white colour samples provided for this purpose. The meat samples were bloomed for 30 min at room temperature and three measurements per sample were taken.
For the cooking loss determination the other loin sub-sample was weighed and placed in a thin-walled plastic bag in a water-bath at 75 °C. After one hour the samples were removed from the water-bath, cooled in cold water, removed from the plastic bag, blotted dry and weighed. Cooking loss was calculated as the difference in sample weight before and after cooking, expressed as a percentage of the initial sample weight (Honikel, 1998).
Tenderness of the meat (same sample as used for cooking loss) was measured with the Warner-Bratzler shear force test using 1.27 cm diameter samples in triplicate (the loin sub-samples were too small to allow more measurements) (Honikel, 1998). Samples were randomly removed from the centre of each cooled (4 °C) M. longissimus lumborum sample. Maximum shear force values (N) required to shear a cylindrical core of 1.27 cm diameter, perpendicular to the grain (at a crosshead speed of 200.0 mm/min), were recorded for each sample. The results were then transformed (by multiplying by a factor of 0.62) to give readings for a 1 cm diameter core and the mean was calculated for each muscle.
The ASREML statistical package (Gilmour et al., 1999) is suitable for the estimation of various random effects in animal breeding, while also allowing for the prediction of least squares means for selected fixed effects, the latter being applied in this investigation. Fixed effects that were considered in the analyses of lamb slaughter traits (carcass measurements, retail cuts, pH measured 45 min, 24 h and 48 h post-slaughter, shear values and colour measurements of the meat) were birth year, sex, birth type, dam line and sire breed as well as relevant interactions. Since interactions were not significant in the vast majority of cases, only main effect means are tabulated. Information is supplied in the text in those cases where significant interactions were found. Age of dam was confounded with production years and was thus not considered. Random sire within ram breed and maternal permanent environmental effects were also computed for the respective traits. Back pedigrees were not included for sires and dams, since all sires and dams were derived from different flocks without genetic linkages. It was thus not attempted to fit maternal genetic effects. It would also be of limited application in a terminal crossbreeding system, where all female progeny are slaughtered. Carcass measurements and retail cut weights were adjusted for carcass weight as there were slight, but significant, differences in carcass weights between the different groups of crossbred lambs (Cloete et al., 2004b). Non-carcass components (head and trotters) were accordingly adjusted for slaughter weight. Carcass temperature had an effect on meat pH (P <0.05), and was included as a covariate in these analyses.
The random effects that were estimated could have been used for the calculation of genetic parameters, but since exhaustive parameter estimates on much more comprehensive data sets are available for most traits in sheep (Safari & Fogarty, 2003; Safari et al., 2005), estimates of these parameters from the current data are not reported. The random effects of sire and dam were, however, retained in the final analyses.
Results
Significant differences between birth years, birth type and gender classes were obtained for some of the traits considered. However, differences between singles and multiples, as well as between male and female lambs were generally consistent with results from the literature. Year effects are transient and not repeatable. These fixed effects were thus not tabulated, except when it was pertinent to the outcomes of the study.
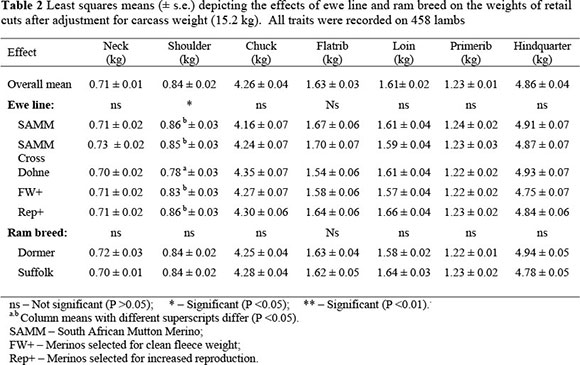
FW+ and Rep+ ewes bred lambs had heavier (P <0.05) heads than Dohne and SAMM ewes (Table 1). There were significant differences (P <0.01) in skin weight between lambs from the two Merino lines and lambs from the dual-purpose ewes, with the progeny of the two Merino lines having heavier skins. No differences in trotter weight were found between lambs from the different ewe lines, but Dormer-sired lambs had heavier trotters than Suffolk sired lambs. The only difference in carcass depth was between the two sire breeds, where the Dormer-sired lambs had an increased carcass depth compared to the progeny of the Suffolk. Carcass width was increased in the crossbred progeny of dual-purpose ewes breeds compared to the progeny of the two purebred Merino lines (P <0.05). The mean leg circumferences of crossbred lambs produced by the dual-purpose ewes and the Rep+ Merino ewes were greater than that lambs from the FW+ Merino ewes (P <0.05). Progeny from Rep+ Merino ewes were intermediate, and not different from either group (P >0.05)
When retail cut weights were considered after adjustment for carcass weight, it was clear that male lambs also had heavier (P <0.01) necks than female lambs (0.73 ± 0.02 vs. 0.69 ± 0.02 kg). The shoulder retail cut weight of Dohne Merino crossbred lambs was lighter (P <0.05) than that of the other types of crossbred lambs. Singles had higher (P <0.01) chuck retail cut weights than multiples (4.29 ± 0.06 vs. 4.23 ± 0.04 kg). Primerib and hindquarter weights were independent of ewe line and ram breed.
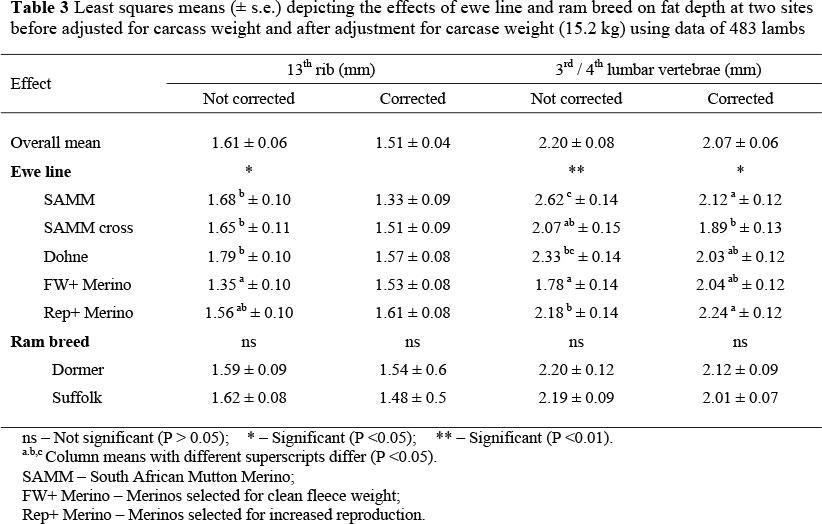
Ewe lambs had a greater (P <0.05) fat cover than rams at the 13th rib, with a corresponding tendency between the 3rd and 4th lumbar vertebra. Multiple lambs were generally leaner (P <0.01) than singles. At the 13th rib, the fat depth of progeny from the dual-purpose breeds (SAMM, SAMM Cross and Dohne Merino) was between 22 and 32% thicker (P <0.05) than that of lambs of the same weight reared by FW+ Merino ewes (Table 3). The progeny of Rep+ Merino ewes resembled those of the dual-purpose ewe lines more closely than their contemporaries reared by FW+ ewes. Adjustment for carcass weight resulted in these ewe line differences being eliminated. Fat depth between the 3rd and 4th lumbar vertebra was affected by a significant interaction between ewe line and birth type. In the case of progeny from purebred SAMM ewes and Dohne ewes, averages for fat depth of singles were respectively 78 and 69% thicker (P <0.05) than that of multiples (3.35 ± 0.24 vs. 1.88 ± 0.14 mm and 2.92 ± 0.20 vs. 1.73 ± 0.17 mm, respectively). The corresponding difference (P <0.05) for single and multiple progeny of FW+ Merino ewes amounted to 38% (2.07 ± 0.21 vs. 1.50 ± 0.18 mm, respectively). No conclusive birth type differences could be demonstrated for the progeny of SAMM cross ewes (2.32 ± 0.27 vs. 1.82 ± 0.14 mm for singles and multiples, respectively) and the progeny of Rep + Merino ewes (2.35 ± 0.22 vs. 2.01 ± 0.15 mm, respectively). Fat depth between the 3rd and 4th lumbar vertebra was also affected by an interaction (P <0.05) between ram breed and birth year. During 2000, Dormer-sired progeny had a thicker (P <0.05) average fat depth at this site than contemporaries sired by Suffolk rams (2.33 ± 0.13 vs. 2.05 ± 0.10 mm, respectively). No sire breed differences were obtained during 2001 (1.65 ± 0.14 vs. 1.74 ± 0.12 mm, respectively) and 2002 (2.38 ± 0.18 vs. 2.24 ± 0.15 mm, respectively).
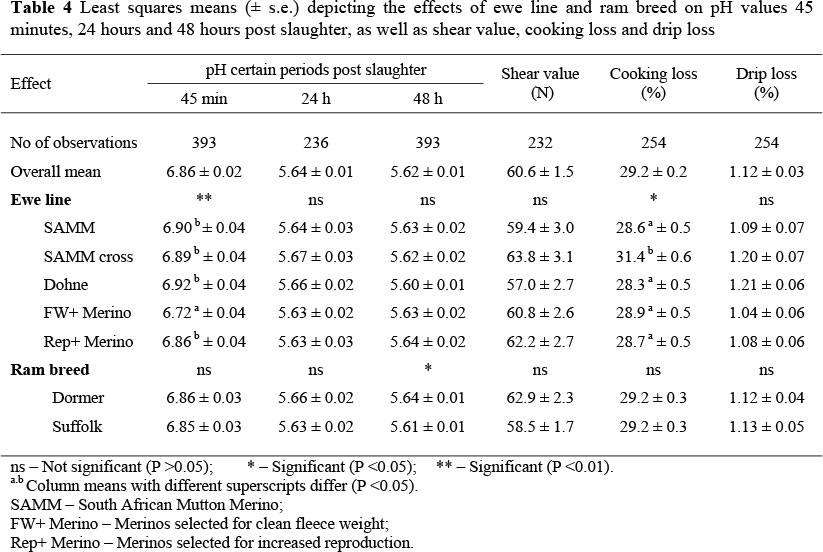
Ram lambs had higher (P <0.05) initial pH values (measured after 45 min) than ewes (6.92 ± 0.03 vs. 6.80 ± 0.03). Multiples also had higher (P <0.05) initial pH values than singles (6.90 ± 0.03 vs. 6.82 ± 0.03). Initial pH values of lambs from the dual-purpose ewe lines were between 0.19 and 0.20 units higher than those of progeny of FW+ Merino ewes (Table 4). Progeny from the Rep+ Merino ewes resembled the dualpurpose ewe lines in this regard, with a 0.14 units higher initial pH than FW+ progeny (P <0.05). Year differences in pH measured at 24 and 48 h post-slaughter were also significant (P <0.05), although the birth years were not ranked similarly throughout (not tabulated). Overall, measurements of pH 24 h post-slaughter were independent (P >0.25) of ewe line and sire breed. Measurements of pH obtained 48 h post-slaughter were affected by sire breed where progeny from the Dormer rams had a higher overall pH value than progeny from Suffolk rams (Table 4). This comparison was complicated by a sire breed x birth year interaction (P <0.05). During 2000, there was no breed difference in ultimate pH between the progeny of Dormer and Suffolk sires (5.97 ± 0.05 vs. 5.95 ± 0.05, respectively).
During 2001 the mean pH measurement made on progeny from Dormer sires was higher than that of progeny sired by Suffolk's (5.78 ± 0.04 vs. 5.52 ± 0.05). No breed differences were again obtained during 2002 (5.98 ± 0.05 vs. 5.87 ± 0.06), but the absolute tendency was in the same direction as for 2001.
Gender did not influence cooking loss, but multiples had a higher (P <0.01) cooking loss than singles (29.7 ± 0.03 vs. 28.7 ± 0.03%). Birth type was also involved in an interaction (P <0.05) with ewe line. Single and multiple born progeny of SAMM ewes and Dohne ewes did not differ with regard to cooking loss (28.2 ± 0.9 vs. 29.1 ± 0.5% and 28.8 ± 0.7 vs. 27.7 ± 0.6% respectively). In the two Merino lines cooking loss differences were generally in favour of singles, namely 27.4 ± 0.7 vs. 30.4 ± 0.7% for the progeny of FW+ Merino ewes, and 27.8 ± 0.5 vs. 29.6 ± 0.6% for the progeny of Rep+ Merino ewes. Results obtained from the progeny of SAMM cross ewes followed an opposite direction, singles having higher cooking loss values than multiples (32.4 ± 1.0 vs. 30.2 ± 0.5%, respectively). Ram breed and year were also involved in a significant (P <0.05) interaction. During 2000 Dormer-sired lambs had higher cooking loss values than Suffolk-sired lambs (23.2 ± 0.5 vs. 21.7 ± 0.5%, respectively). An opposite result was obtained during 2001, Dormer-sired progeny having a lower average cooking loss than contemporaries sired by Suffolk rams (31.4 ± 0.5 vs. 33.4 ± 0.5%, respectively). No ram breed differences were found during 2002 (32.9 ± 0.5 vs. 32.5 ± 0.5% for progeny of Dormer and Suffolk rams, respectively). A significant (P <0.05) interaction between breed and birth year was also found for drip loss. During 2000 and 2002, Dormer-sired lambs tended to have higher drip loss values than Suffolk-sired lambs (1.09 ± 0.06 vs. 1.03 ± 0.07% and 1.06 ± 0.06 vs. 0.93 ± 0.08% respectively). An opposite result was obtained during 2001, Dormer-sired progeny having a lower (P <0.05) average drip loss than contemporaries sired by Suffolk rams (1.20 ± 0.06 vs. 1.42 ± 0.08%, respectively). Shear strength was unaffected by ewe line and sire breed.
When meat colour was considered, it was clear that meat from male lambs had generally higher (P <0.05) L* values (37.7 ± 0.2 vs. 36.6 ± 0.2) than ewe lambs. Suffolk-sired progeny also had generally higher (P <0.05) L* values than Dormer-sired progeny (Table 5). The derived L* values were independent of ewe line. Progeny of FW+ Merino ewes had meat that was situated more towards saturated red on the green-red scale than progeny of purebred SAMM ewes and SAMM cross ewes. There was also a suggestion of values closer to saturated yellow in the case of Suffolk-sired progeny (P <0.25), but it did not reach significance. Hue angle was not affected by ewe line (P >0.05). Suffolk-sired lambs tended to have a higher (P <0.10) hue angle than Dormer-sired lambs. Chroma was not affected by either ewe line or sire breed. No significant interactions were found in these analyses.
Discussion
All the animals were slaughtered at the same abattoir using mostly the same staff throughout the experiment. It was impossible to slaughter all the lambs on the same day under the same environmental conditions, since they were slaughtered as they reached a target weight. However, it was assumed that potential environmental effects would have been evened out. No abnormal environmental conditions (e.g. excessive heat, humidity, long transport, etc.) were noted on the days that the lambs were slaughtered.
Rams had heavier head and neck weights than ewes in the present study, as was also found by Jeremiah et al. (1997). The phenomenon of heavier forequarters in male sheep is typical of sexual dimorphism. Lambs from the two Merino lines also had heavier head weights than those of the dual-purpose ewes. This could be related to the fact that Merinos have horns, which are transferred to some of their crossbred progeny (Campher et al., 1998). As Merinos are specialist wool-producing sheep, it could be expected that crossbred lambs from the two Merino lines were also likely to have heavier skin weights than the crossbred progeny of the dual-purpose ewes. Female lambs took longer to reach slaughter weight (Cloete et al., 2004b), resulting in a longer wool growth period and consequently heavier skin weights.
A gender influence on subcutaneous fat depth is consistent with trends generally found in the literature, with ewe carcasses being fatter and with a higher dressing percentage (Kirton et al., 1982; Cloete et al., 2005). The significant effect of birth type on fat depth was consistent with the study of Greeff et al. (2003) where singles were fatter than multiples at slaughter, based on data for 829 Merino ram hoggets.
Lambs produced by the dual purpose and Rep+ Merino ewes had a higher meat pH 45 min post slaughter than the progeny of the FW+ Merino ewes. This finding may be related to a greater susceptibility to pre-slaughter stress, since a low pH 45 min post slaughter is regarded as an indicator of acute, short-term stress (Lawrie, 1998). These results could be related to a previous study where purebred Rep+ line progeny had higher pH values within 45 min after slaughter than contemporaries from a line that was selected against reproduction (Cloete et al., 2005). The contention that selection resulted in reduced levels of fear from humans was supported by behavioural observations that Rep+ line progeny allowed shorter distances between them and a human operator when unaccustomed to handling, while they were also less likely to defecate during the test. Although there were differences in pH measured 45 min after slaughter between dam lines there were no differences in final pH values (48 h after slaughter) between the lambs from the different ewe lines. The viewpoint that purebred Merino lambs have a higher ultimate meat pH than other breeds is often expressed (Greeff et al., 2003). In contrast, no differences in meat pH were found between lambs of five diverse genotypes (two specialist meat sheep breeds crossed to Border Leicester x Merino and pure Merino ewes) and purebred Merino lambs in the study of Safari et al. (2001). Hopkins & Fogarty (1998), Fogarty et al. (2000) and Purchas et al. (2001) also found no differences in the ultimate pH between lambs sired by different rams. The fact that there were no differences in final pH in the present study could also indicate that the lambs were not really put under severe stress (Lawrie, 1998). Limited genetic variation was found in pH measured 24 h after slaughter in the study of Greeff et al. (2003), supporting the lack of genetic differences between ewe lines and sire breeds in the present study. In contrast, Fogarty et al. (2003) found a higher heritability estimate for muscle pH of 0.27 ± 0.09. More recently, Safari et al. (2005) derived an average h2 of 0.18 for meat pH from two literature values, with a very low coefficient of variation of 6% for the trait. It thus appears that, although some genetic variation may exist for pH, it may not be substantial.
Juiciness of meat is directly related to the intramuscular lipids and moisture content of meat. In combination with water, the melted lipids constitute a "broth" that is retained in meat (Schönfeldt et al., 1993). Overall, cooking loss from fatter meat tends to be less than from lean meat. Although fatter meat loses more fat (which is expected in view of a higher fat content), it loses less moisture, possibly because the structural changes caused by the presence of the fat enhance the water-holding capacity (Lawrie, 1998). This reasoning could explain why the cooking loss of the meat from multiples was generally higher than that of meat from the singles. The thicker average fat cover of singles compared to multiples provides further support to this line of reasoning. Ewe line interacted with birth type for fat thickness at the 3rd and 4th lumbar vertebra, with no birth type differences in the progeny of SAMM cross and Rep+ Merino ewes. No apparent reason could be provided for this interaction, except that the Rep+ line was selected for their ability to rear multiples. This selection resulted in an improved lamb survival of multiples in particular (Cloete & Scholtz, 1998). It is also generally accepted that improved maternal care contributes to the better overall reproduction of crossbred ewes. It is of interest to note that the latter two ewe lines had the best overall reproduction in absolute terms (Cloete et al., 2003; 2004b).
In accordance with the results from the present study, Hoffman et al. (2003) found no overall ewe line or ram breed differences in cooking loss and drip loss between lambs sampled from the same experiment. It needs to be stated that only a small number of lambs born in 2000 were used. Interactions of sire breed with birth year for fat depth, pH, cooking loss and drip loss could possibly be associated with the sampling of sires for usage in specific years.
No significant differences in shear values were found between the ewe lines or sire breeds. Hopkins & Fogarty (1998) and Safari et al. (2001) accordingly did not find any differences in the shear values of five diverse crossbred genotypes and purebred Merinos. Young & Dobbie (1994) compared the inter-muscular collagen content of purebred Romney lambs and lambs sired by a terminal sire breed (Texel). They found that there was no effect of breed on collagen content between slaughter ages of 100 to 215 days. The mean slaughter age of progeny from the respective ewe lines in the present experiment ranged from 134 to 165 days (Cloete et al., 2004b). These results suggest that the use of terminal sires is unlikely to affect the tenderness of lamb slaughtered in this range. In support of this argument, Hoffman et al. (2003) found no differences in tenderness between lambs from different genotypes originating from the same experiment in a taste panel. It has to be conceded that only a small number of lambs born in 2000 were used for the latter investigation. The absolute shear values reported in this investigation are lower compared to that reported for South African sheep breeds in previous investigations where the same techniques were used in the same laboratory (Sheridan et al., 2003; Cloete et al., 2005). This difference stems from the adjustment to shear values described earlier. However, the overall mean of ~60 N was about twice as high as means of 20 - 33 N that were reported for Australian lamb (Hopkins & Fogarty, 1998; Safari et al., 2001). Other studies reported intermediate overall means for tenderness, ranging from 36 - 46 N (Perlo et al., 2008; Toohey et al., 2008). The possibility could exist that the methodology employed in the present investigation differs from that of the other reports and this may explain these high values. It is well known that cooking temperature (and duration) will have an influence on the gelatinisation of the collagen. This phenomenon always makes comparisons between different laboratories difficult.
Priolo et al. (2000) and Sanudo et al. (1998) found that muscle with a higher initial pH tended more towards the saturated red end of the colour scale. Previous observations on the pH and a* value of meat produced by the progeny of Merino ewes divergently selected for reproduction also supports this line of reasoning (Cloete et al., 2005). Although there were some differences in colour between years and sexes, there were no substantial differences between lambs from the different dam lines. Hopkins & Fogarty (1998) accordingly did not find major differences in colour measurements between cryptorchid lambs of five crossbred genotypes and purebred Merinos. Greeff et al. (2003) reported limited genetic variation for meat colour measurements. Respective h2 estimates subsequently derived from the literature were 0.16 for L*, 0.04 for a* and 0.05 for b*, as based on two previous estimates from the literature (Safari et al., 2005). Values for pH and colour measurements after 24 h of cooling were also unaffected by birth type in the study of Greeff et al. (2003), which conforms to the general trend found in the present study.
Conclusion
Terminal crossbreeding of five Merino-type ewe lines with Dormer and Suffolk sires did not have adverse effects on the meat quality and retail cut weights of the lamb produced. The utilization of heterosis in terminal crossbreeding systems with specialist meat breeds may thus be promoted to improve farm income in South Africa, as elsewhere (Fogarty, 2006). Differences that were found were mostly between the progeny of dual-purpose ewes and the Merino ewes selected for clean fleece weight (FW+). Progeny from the latter line had a lower initial pH and a reduced fat depth. Relatively inexpensive components of the carcass, viz. the skins and heads of progeny of both Merino dam lines were heavier than those of the dual-purpose lines. Crossbred progeny of the Merino lines, however, still performed satisfactorily for all the traits considered. Decisions pertaining to the choice of ewe line could thus be based on other concerns, such as the relative economic return of the genotype (Cloete et al., 2004b). It was interesting to note that the crossbred progeny of Rep+ Merino ewes resembled the progeny of dual-purpose ewes rather than that of FW+ Merino ewes for traits such as initial pH and fat depth. Previous work has also suggested that selection for reproduction in the Rep+ line may have improved some slaughter traits (Cloete et al., 2004a; 2005).
No clear advantage in favour of either one of the ram breeds was found. The choice of rams between the two breeds could thus be based on other factors like the availability of rams, and considerations such as the probability of contamination with coloured fibres in the case of the Suffolk breed (Erasmus et al., 1983).
Acknowledgement
We gratefully acknowledge the support of the staff at the Langgewens experimental farm during the experiment. We also wish to thank the personnel of Roelcor abattoir for their friendly co-operation. The experiment was supported financially by the South African Red Meat Research and Development Trust. The South African Wool Industry supported the development of the Rep+ line financially. We are also grateful to the Western Cape Dohne Merino club, for the donation of the 1997 born progeny from the Kromme Rhee nucleus flock. We are also indebted to A. Kotzé of the South African Suffolk Breeder's Association, who provided Suffolk rams for the experiment.
References
Bruwer, G.G., Naude, R.T. & Vosloo, W.A., 1987. An evaluation of the lamb and mutton carcass grading system in the Republic of South Africa. 1. A survey of carcass characteristics of the different grades. S. Afr. J. Anim. Sci. 17, 79-84. [ Links ]
Cloete, J.J.E., Cloete, S.W.P. & Hoffman, L.C., 2005. Behaviour of Merinos divergently selected for multiple rearing ability in response to external stimuli. Small Rumin. Res. 60, 227-236. [ Links ]
Cloete, J.J.E., Cloete, S.W.P., Hoffman, L.C. & Fourie, J.E., 2004a. Slaughter traits of Merino sheep divergently selected for multiple rearing ability. S. Afr. J. Anim. Sci. 34, 189-196. [ Links ]
Cloete, S.W.P. & Scholtz, A.J., 1998. Lamb survival in relation to lambing and neonatal behaviour in medium wool Merino lines divergently selected for multiple rearing ability. Aust. J. Exp. Agric. 38, 801-811. [ Links ]
Cloete, S.W.P., Cloete, J.J.E., Durand, A. & Hoffman, L.C., 2003. Production of five Merino type lines in a terminal crossbreeding system with Dormer or Suffolk sires. S. Afr. J. Anim. Sci. 33, 223-232. [ Links ]
Cloete, S.W.P., Cloete, J.J.E., Herselman, M.J. & Hoffman, L.C., 2004b. Relative performance and efficiency of five Merino dam lines in a terminal crossbreeding system with Dormer or Suffolk sires. S. Afr. J. Anim. Sci. 34, 135-143. [ Links ]
Campher, J.P., Hunlun, C. & Van Zyl, G.J., 1998. South African Livestock Breeding. South African Stud Book and Livestock Improvement Association. PO Box 270, Bloemfontein 9300, South Africa. [ Links ]
Commision International De L' Eclairage, 2nd, 1976. Commision Internationale De L' Eclairage, 18th session, London, England. September 1975. CIE publication no. 36. [ Links ]
De Villers, T.T. & Cloete, S.W.P., 1984. The Dormer sheep breed. Proc. 2nd World Cong. Sheep Beef Cattle Breed. 16 - 19 April 1984, Pretoria. South African Stud Book and Livestock Improvement Association, Bloemfontein (1984), 695-698. [ Links ]
Doloksaribu, M., Gatenby, R.M., Subandriyo, S.T. & Bradford, G.E., 2000. Comparison of Sumatra sheep crossbreds. III. Reproductive performance of F2 ewes and weights of lambs. Small Rumin. Res. 38, 115-121. [ Links ]
Ellis, M., Webster, G.M., Merrell, B.G. & Brown, I., 1997. The influence of terminal sire breed on carcass composition and eating quality of crossbred lambs. Anim. Sci. 64, 77-86. [ Links ]
Enright, W.J., Quirke, J.F., Gluckman, P.D., Breier, B.H., Kennedy, L.G., Hart, I.C., Roche, J.F., Coert, A. & Allen, P., 1990. Effects of long-term administration of pituitary-derived bovine growth hormone and estradiol on growth in steers. J. Anim. Sci. 68, 2345-2356. [ Links ]
Erasmus, L.S., De Kock, J.A. & Grobler, J.W., 1983. Slaglamproduksie in die Suid-Kaap. Elsenburg J. 7, 13-32. [ Links ]
Fogarty, N.M., 2006. Utilization of breed resources for sheep production. 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, MG, Brasil. [ Links ]
Fogarty, N.M., Hopkins, D.L. & Van der Ven, R., 2000. Lamb production from diverse genotypes. 2. Carcass characteristics. Anim. Sci. 70, 147-156. [ Links ]
Fogarty, N.M., Safari, E., Taylor, P.J. & Murray, W., 2003. Genetic parameters for meat quality and carcass traits and their correlation with wool traits in Australian Merino sheep. Aust. J. Agric. Res. 54, 715-722. [ Links ]
Freking, B.A., Leymaster, K.A. & Young, L.D., 2000. Evaluation of Dorset, Finnsheep, Romanov, Texel, and Montadale breeds of sheep: I. Effects of ram breed on productivity of ewes of two crossbred populations. J. Anim. Sci. 78, 1422-1429. [ Links ]
Gilmour, A.R., Cullis, B.R., Welham, S.J. & Thompson, R., 1999. ASREML - Reference manual. NSW Agricultural Biometric Bulletin No. 3. NSW Agriculture, Orange Agricultural Institute, Forest Road, Orange 2800, NSW, Australia. [ Links ]
Greeff, J.C., Davidson, R. & Skerritt, J.W., 2003. Genetic relationship between carcass quality and wool production traits in Australian Merino rams. Proc. Assoc. Advmt. Anim. Breed. Gen. 15, 330-333. [ Links ]
Hoffman, L.C., Muller, M.M., Cloete, S.W.P. & Schmidt, D., 2003. Comparison of six crossbred lamb types: sensory, physical and nutritional meat quality characteristics. Meat Sci. 65, 1265-1274. [ Links ]
Honikel, K.O., 1998. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 49, 447-457. [ Links ]
Hopkins, D.L. & Fogarty, N.M., 1998. Diverse lamb genotypes. 2. Meat pH, colour and tenderness. Meat Sci. 49, 477-488. [ Links ]
Hopkins, D.L., Gilbert, K.D., Pirlot, K.L. & Roberts, A.H.K., 1996. Elliottdale and crossbred lambs: growth rate, wool production, fat depth, saleable meat yield, carcass composition and muscle content of selected cuts. Aust. J. Exp. Agric. 32, 429-434. [ Links ]
Jeremiah, L.E., Jones, S.D.M., Tong, A.K.W., Robertson, W.M. & Gibson, L.L., 1997. The influence of lamb chronological age, slaughter weight and gender on yield and cutability. Int. Sheep Goat Res. J. 13, 39-46. [ Links ]
Kirton, A.H., Clarke, J.N. & Hickey, S.M., 1982. A comparison of the composition and carcass quality of Kelly and Russian castrate, ram, wether and ewe lambs. Proc. N. Z. Soc. Anim. Prod. 42, 117-118. [ Links ]
Lawrie, R.A., 1998. Lawrie's Meat Science. 6th ed. Woodhead Publ. Ltd. Cambridge, England. [ Links ]
Milton, J.T.B., Banks, R.B., Hepworth, G.W. & Dorman, S.A., 1994. Genetic evaluation of American Suffolks through progeny testing. Proc. Aust. Soc. Anim. Prod. 20, 167-177. [ Links ]
Perlo, F., Bonato, P., Teira, G., Tisocco, O., Vicentin, J., Pueyo, J. & Mansilla, A., 2008. Meat quality of lambs produced in the Mesopotamia region of Argentina finished on different diets. Meat Sci. 79, 521-528. [ Links ]
Piper, L. & Ruvinsky, A., 1997. The genetics of sheep. CAB international 198 Madison Avenue New York, NY 10016-4314 USA. [ Links ]
Priolo, A., Waghorn, G.C., Lanza, M., Biondi, L. & Pennisi, P., 2000. Poleythylene glycol as a means for reducing the impact of condensed tannins in carob pulp: Effects on lamb growth performance and meat quality. J. Anim. Sci. 78, 810-816. [ Links ]
Purchas, R.W., Silva Sobrinho, A.G., Garrick, D.J. & Lowe, K.I., 2001. Effects of age at slaughter and sire genotype on fatness, muscularity, and the quality of meat from ram lambs born to Romney ewes. N. Z. J. Agric. Res. 45, 77-86. [ Links ]
Safari, E. & Fogarty, N.M., 2003. Genetic parameters for sheep production traits: Estimates from the literature. Tech. Bull. 49. New South Wales Agriculture, Orange, Australia. Document available at: http://www.sheep.crc.org.au/articles.php?rc=145. [ Links ]
Safari, E., Fogarty, N.M. & Gilmour, A.R., 2005. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest. Prod. Sci. 92, 271-289. [ Links ]
Safari, E., Fogarty, N.M., Ferrier, G.R., Hopkins, L.D. & Gilmour, A., 2001. Diverse lamb genotypes. 3. Eating quality and the relationship between its objective measurement and sensory assessment. Meat Sci. 57, 153-159. [ Links ]
Sañudo, C., Sierra, I., Olleta, J.L., Martin, L., Campo, M.M., Santolaria, P., Wood, J.D. & Nute, G.R., 1998. Influence of weaning on carcass quality, fatty acid composition and meat quality in intensive lamb production systems. Anim. Sci. 66, 175-187. [ Links ]
Schönfeldt, H.C., Naudè, R.T., Bok, W., Van Heerden, S.M. & Smit, R., 1993. Flavour- and tenderness-related quality characteristics of goat and sheep meat. Meat Sci. 34, 363-379. [ Links ]
Sheridan, R., Hoffman, L.C. & Ferreria, A.V., 2003. Meat quality of Boar goat kids and Mutton Merino lambs. 2. Sensory meat evaluation. Anim. Sci. 76, 73-79. [ Links ]
Snowder, G.D. & Duckett S.K., 2003. Evaluation of the South African Dorper as a terminal sire breed for growth, carcass, and palatability characteristics. J. Anim. Sci. 81, 368-375. [ Links ]
Stanford, K., Woloschuk, C.M., McClelland, L.A., Jones, S.D.M. & Price, M.A., 1997. Comparison of objective external carcass scores for prediction of lamb carcass quality. Can. J. Anim. Sci. 77, 217-223. [ Links ]
Teixeira, A., Delfa, R. & Treacher, T., 1996. Carcass composition and body fat depots of Galego Bragançano and crossbred lambs by Suffolk and Merino Precoce sire breeds. Anim. Sci. 63, 389-394. [ Links ]
Toohey, E.S., Hopkins, D.L., Stanley, D.F. & Nielsen, S.G., 2008. The impact of new generation pre-dressing medium-voltage electrical stimulation on tenderness and colour stability of lamb meat. Meat Sci. 79, 683-691. [ Links ]
Young, O.A. & Dobbie, J.L., 1994. Characteristics of intermuscular collagen. N. Z. J. Agric. Res. 37, 97-105. [ Links ]
 Correspondence:
Correspondence:
E-mail: jasperc@elsenburg.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}