










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.4 Pretoria Apr. 2008
Systematic factors that affect ostrich egg incubation traits
Z. BrandI, II; S.W.P. CloeteI, III; C.R. BrownIV; I.A. MaleckiV
IDepartment of Animal Sciences, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africa
IIInstitute for Animal Production: Oudtshoorn, PO Box 313, Oudtshoorn 6620, South Africa
IIIInstitute for Animal Production: Elsenburg, Private Bag X1, Elsenburg 7607, South Africa
IVInstitute of Science & the Environment, University of Worcester, Henwick Grove, Worcester, WR2 6AJ, UK
VSchool of Animal Biology, Faculty of Natural and Agricultural Science, University of Western Australia, Crawley WA, 6009, Australia
ABSTRACT
Data obtained from a pair-mated ostrich flock maintained at Oudtshoorn, South Africa, were used to estimate environmental and genetic parameters for egg weight (EWT), water loss of incubated eggs up to 21 days (WL21), water loss up to 35 days (WL35), pipping time (PT) and weight of day-old chicks (CWT). Between 13806 and 19913 artificially incubated ostrich eggs during the 2003 to 2006 production years were used. Systematic factors affecting these traits such as production year, breeding season, female age, incubator type, storage time and ostrich breed, were initially assessed in single-trait-analyses, using ASREML. Eggs and chicks produced by Zimbabwean Blue (ZB) females were 5 and 7% heavier, respectively, than those produced by South African Black (SAB) females. WL21 and WL35 were not significantly different between ZB and SAB birds. There were trends for within-season effects on EWT and CWT, but no general, robust trend applicable to all years could be discerned. Season had a significant effect on WL21, WL35 and PT. An increase was apparent in EWT, CWT and PT with an increase in female age. There was a linear increase in pipping time as egg storage time prior to incubation increased. Incubator type had an effect on WL21 and WL35. Systematic factors affect traits such as WL21, WL35 and PT and should be accounted for before the estimation of genetic parameters. These factors should be considered when planning commercial ostrich husbandry and artificial incubation operations.
Keywords: Embryo development, eggshell, albumen, egg water loss, hatchability
Introduction
Artificial incubation has become an integral part of any commercial ostrich enterprise. Successful artificial incubation is, however, affected by a number of factors including storage conditions of eggs prior to setting in the incubator, water loss from the egg during incubation, season, and female age and genetic makeup (Blood et al., 1998; Van Schalkwyk et al., 1999; Brand et al., 2007). Peak production for ostriches in the southern hemisphere occurs between winter (July) and summer (January) (Lambrechts, 2004). Ostrich females begin laying at 2 to 2.5 years old and peak egg and chick production are achieved at 8 to 9 years of age. Female age, however, is known to influence the number of eggs laid as well as egg weight and, consequently, chick weight at hatching (Bunter & Cloete, 2004; Ipek & Sahan, 2004; Lambrechts, 2004; Cloete et al., 2006).
Pre-incubation storage leads to morphological changes in the blastoderm and to a reduced growth rate of the embryos of chickens (Fasenko et al., 1992; Meijerhof, 1992) and ostriches (Malecki et al., 2005). Albumen quality is compromised by a prolonged storage time (Bradley, 1997), which can lead to a proportionate increase in early embryonic mortality in duck and quail eggs (Narahari et al., 1991; Yildirim, 2005). Storage of ostrich eggs for periods longer than seven days results in an increase in embryonic mortality (Brand et al., 2007). Because the storage time and storage temperature of ostrich eggs are usually variables that can easily be managed, it is important to get an indication of possible factors predisposing ostrich eggs that were stored for longer periods to a higher incidence of embryonic deaths.
Avian eggs lose water during incubation and the amount of water lost is important for successful hatching. Achieving the appropriate water loss during artificial incubation is one of the problems regularly encountered because water loss is influenced by a number of factors including physical characteristics of the eggshell, incubator conditions and heat production of the developing embryo (Ar, 1991). Swart et al. (1987) determined that the total water loss from eggs in natural ostrich nests amounts to about 13% of the initial egg weight; the main driving force behind incubation water loss being the gradient in water vapour pressure across the incubating eggshell. Results from studies by Deeming (1995), Ar (1996) and Blood et al. (1998) showed that the optimal water loss for artificially incubated ostrich eggs amounts to approximately 15% but, like other birds, ostriches show some latitude in the amount of water loss at which eggs will still hatch successfully. Eggs which lost less than 10% or more than 20% of their initial weight were less likely to hatch. Horbaňczuk et al. (1999) found a higher incidence of malpositioned chicks and chicks with unabsorbed yolk sacs if incubator humidity was too high. Relative humidity and thus vapour pressure inside the incubator can, to a large extent, be controlled during artificial incubation.
Genetic make-up is one of the factors influencing the performance of individuals (Petitte & Davis, 1999). Egg quality is reported to have significant genetic components (Stewart, 1995). Shell deaths were accordingly influenced by breed combination in the study of Brand et al. (2007), involving the South African Black and Zimbabwean Blue breeds. At present there is no indication of how these differences are related to evaporative water loss of the eggs produced by the different breed combinations. No quantitative information is available for the water loss from eggs of different ostrich lines or strains.
A better understanding of how systematic factors influence the successful artificial incubation of ostrich eggs is essential (Cloete et al., 2002). The aim of this study was thus to quantify the effects of environmental factors such as production year, season, female age, the incubator type used and storage time on egg weight, chick weight, water loss and pipping time of ostrich eggs. The effect of breed combination was also assessed as a systematic effect.
Material and Methods
The experimental population used for the study (2003 to 2006) was the commercial, pair-bred ostrich flock at the Oudtshoorn Research Farm in the Klein Karoo region of South Africa. The origin of the flock and the general management procedures implemented have been described previously (Van Schalkwyk et al. , 1998; Bunter & Cloete, 2004). During 2003, Zimbabwean Blue (ZB) breeders (21 females; 33 males) were introduced to the flock and mated in various combinations with South African Black (SAB) males and females (Brand et al., 2005). Data that were recorded for 2003 to 2006 thus involved various combinations of the two purebred bloodlines (SAB and ZB), as well as the reciprocal crosses between them. The flock consisted of 188 breeding pairs. Birds in the flock ranged between 2 and 11 years of age, and the annual breeding season usually lasted for about eight months followed by a four month rest period. A total of 20740 eggs were available for analyses. After excluding eggs with broken or cracked shells and eggs used in other experiments, a total of 19913 eggs were analysed. The number of records analysed ranged from 13248 for chick weight to 19913 for egg weight. Methods of collection, sanitation and storage of eggs at the research farm are well documented (Van Schalkwyk et al., 1998; Van Schalkwyk et al., 1999; Bunter & Cloete, 2004). Briefly, eggs were collected daily, weighed and identified by date and paddock (female) of origin. The surface of each egg was sterilized by 20 min of ultraviolet exposure and labelled with a permanent marker. With the exception of the first two weeks of the breeding season, eggs were stored for no more than six days at a temperature of 17 °C and relative humidity (RH) of 75%. During the first two weeks of the breeding season, egg production is still very low and eggs were stored for 14 days to accumulate enough eggs to put in setters. Eggs were then artificially incubated at 36 °C and 24% RH in Buckeye®, Prohatch®, African International® or Natureform® incubators. The capacity and operation of the incubators, with the exception of the African International® incubator are described by Cloete et al. (2001). Information regarding the African International® incubator can be obtained from the paper by Brand et al. (2007). Details of the genotype, female age (only known explicitly for all individuals in the SAB breeds), year, season, and specific incubator used were known for individual eggs. Traits that were considered were egg weight at collection (EWT) and at candling after 21 or 35 days of incubation. These weights were used to derive water loss (% of fresh egg weight) up to 21 (WL21) and 35 days (WL35) of incubation. Eggs were transferred to the hatcher at day 35 of incubation and were inspected twice daily (at 08:00 and 16:00) for external pipping from day 39 of incubation. The pipping time of the eggs was recorded. These data were used to derive the number of days from the commencement of incubation to the recorded external pipping time (PT). Day-old chick weight was recorded after the chicks were allowed to dry off for 24 hours.
The data were subjected to a genetic analysis, using ASREML software (Gilmour et al., 1999). The software is suitable for fitting a wide range of random effects in animal breeding, while least-squares means for selected systematic effects are predicted simultaneously. Such effects are tested for significance, using an F-test in the analysis of variance table. Fixed effects that were considered included sire line (SAB or ZB), dam line (SAB or ZB), year of production (2003 - 2006), laying season (winter, spring or summer), female age (2 to 11 years), incubator (as defined above) and storage time (0 days to 7+ days). Various two-factor interactions were considered initially, but only the year x season interaction was significant and retained in the final analyses. The sire line x dam line interaction was also estimated, although it only approached significance in the analysis on WL35 (P = 0.09). This paper only includes information on the fixed effects that were considered. Random effects and genetic parameters are reported in a subsequent paper (Brand et al., 2008).
Results and Discussion
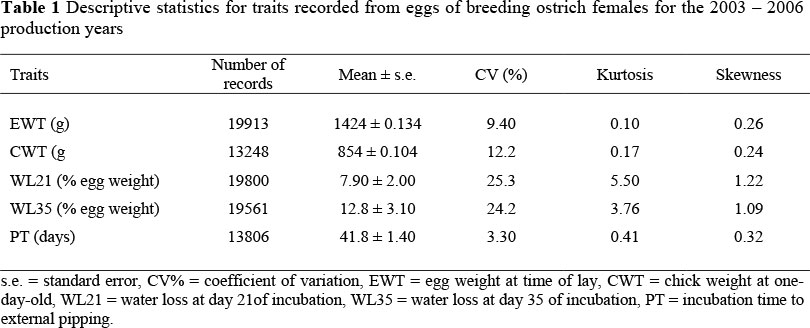
The average weight of over 19000 ostrich eggs was 1424 g with a coefficient of variation (CV) of 9.4%, and the average weight of day-old chicks was 854 g with a coefficient of variation of 12.2% (Table 1). These are consistent with previous results from the same breeding population (Cloete et al., 1998; Bunter et al., 1999; Bunter & Cloete, 2004). The average chick weight represented about 60% of fresh egg weight, which corresponds with other avian species (Wilson, 1991a). Incubation time to external pipping averaged 42 days and had a very low CV of 3.3% (Table 1). No comparable estimate could be found in the literature. In contrast, CV's of WL21 and WL35 were higher at 25.5% and 24.2%, respectively. The change in egg weight at 35 days, expressed as a percentage of initial egg weight, is an indication of an evaporative water loss of approximately 13% of fresh egg weight during incubation, which is characteristic of ostriches (Swart & Rahn, 1988; Blood et al., 1998). Deviations from normality for WL21 and WL35 involved kurtosis rather than skewness, and the interpretation of the results was thus continued without attempting to transform the data to obtain a better distribution (Glass et al., 1972).
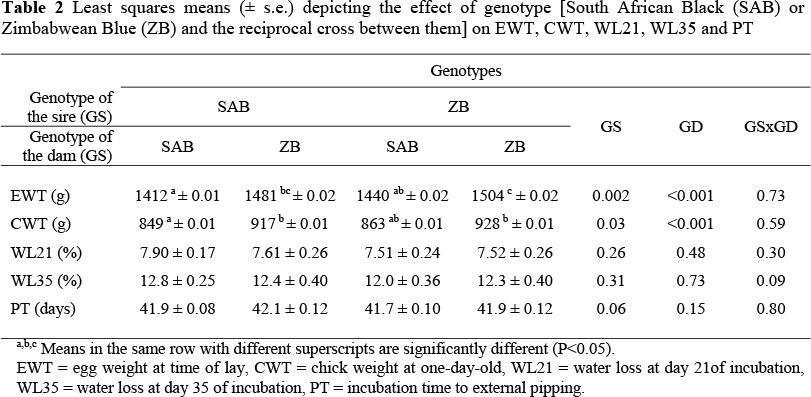
Zimbabwean Blue females laid significantly heavier eggs (5%), which resulted in their chicks being 7% heavier than those of SAB females (P <0.05; Table 2). Crossing ZB males with SAB females also resulted in heavier eggs being laid by their SAB mates compared to pure-bred SAB birds, although the difference only approached significance (P <0.10). Eggs produced by the reciprocal cross (SAB males x ZB females) were substantially heavier than those produced by SAB females subjected to pure breeding. On average, ZB breeding birds are approximately 10% heavier than the SAB (Jarvis, 1998; Brand et al., 2005), which can be a contributing factor to the heavier eggs produced by ZB birds. Water loss over the periods up to 21 and 35 days of incubation was independent of sire line, dam line or the sire line x dam line interaction.
Incubation time to external pipping (PT) was not different (P >0.05) between the different breeds and breed combinations (Table 2). Incubation water loss was generally independent of sire bloodline and/or dam bloodline, although there was a tendency for an interaction between these main effects for WL35 (P = 0.09).
All traits were affected by an interaction between year of production and season (winter, spring or summer) of production (P <0.05). Trends within seasons could be discerned, but no general robust trend applicable to all years was observed. The effects of the year x season interaction are thus detailed in Figures 1 to 3 for CWT, WL35 and PT. There appears to be a more or less linear decline in CWT as the season progressed during 2003 and 2005. Day-old weight of chicks hatched during spring resembled winter values during 2004 and 2006, with a subsequent decline towards the summer.
During 2004, WL35 increased more or less linearly as the season progressed from winter to summer (Figure 2). Seasonal effects were not as obvious during 2003 and 2005, although the absolute values tended to increase between winter and summer. Conversely, WL35 increased from winter to spring in 2005, with no further change to summer. The reasons for these interactions are unclear, but slight differences or changes between year-season ambient climate and between incubators in the micro environment of the incubators could contribute. Although the incubators were set at 24% RH and 36 °C, the absolute humidity or vapour pressure and temperature surrounding the eggshell could differ from these settings because of incubator design and ambient conditions inherent to incubators and year-season combinations. It is generally accepted that year-season effects are transient and unpredictable and unlikely to be repeated. All these factors could contribute to the observed findings. Another possible contributing factor is changes in eggshell structure or egg composition between year-season combinations. More research is required to determine whether seasonal changes in eggshell structure (as determined by the female) contributed to this variation between year and season for water loss. Previous research suggested that eggshell characteristics of females were adapted to mirror changes in the ambient climate (Cloete et al., 2006). However, the data used by Cloete et al. (2006) were obtained in a single production year, and should be subjected to further investigation. The hatching of lighter chicks from eggs with a higher water loss can be expected. The overall outcome of the trends for CWT, i.e. general decline from winter to summer, and for WL35, i.e. general increase from winter to summer, is therefore reasonable.
In general, ostrich chicks hatching from eggs laid during the summer had a shorter PT compared to those hatching from eggs laid in winter (Figure 3). During spring, average PT more closely resembled means for the winter in 2004-2006. The opposite was true during 2003, when the PT of chicks hatching during spring was significantly lower than those hatching in the winter, and was closer to summer pipping times. We did not find comparable results in the literature on this subject.
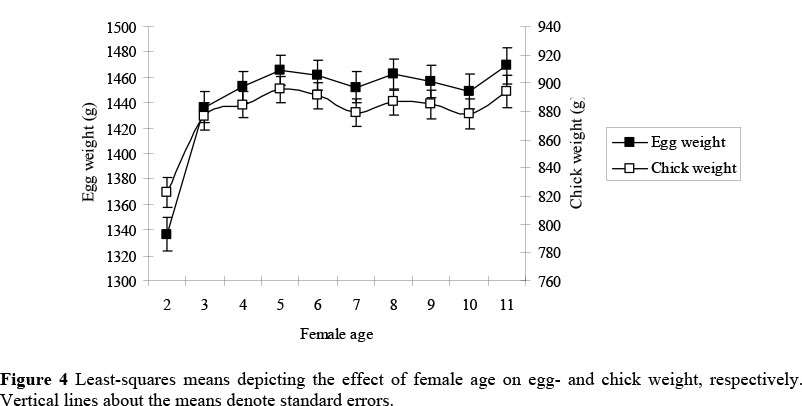
Ostrich females have a markedly longer productive life compared to other poultry species (Ipek & Sahan, 2004), making it difficult to compare ostriches to the smaller domestic poultry species traditionally used for egg and chick production. In our study, female age significantly influenced all the traits under consideration. Ostrich EWT and CWT increased by about 7% between the second and third years of production of individual females (P <0.05). These traits reached their peak at five years of age (P <0.05; Figure 4), as previously noted by Bunter & Cloete (2004) and Ipek & Sahan (2004). Production was then effectively constant until 11 years-old. Bunter & Cloete (2004) reported that both EWT and CWT decreased at older ages, but this decrease was only evident in the eggs or chicks of females of 11 - 12 years and older. These age groups were not represented in the data set used for the present study, as the population was allowed to become substantially younger (Cloete et al., 2006).
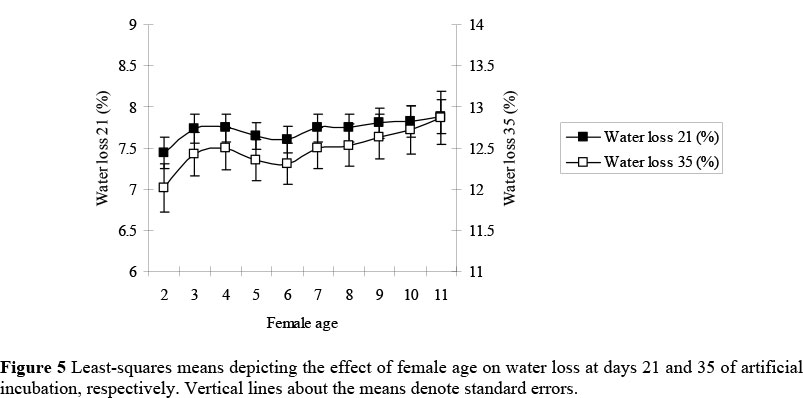
Water loss to 21 and 35 days of incubation was largely independent of female age (Figure 5), although some evidence of an incline was present (P = 0.16 for WL21 and P = 0.06 for WL35). There is strong evidence to suggest that embryonic mortality of ostrich chicks increases with age of ostrich females (Brand et al., 2007). The present results, however, are inconclusive whether this increase could be attributed to age-related changes in water loss. More research is required to better understand the mechanism involved in a reduced embryonic survival of eggs produced by older females. Although speculative, it could possibly be attributed to a decrease in albumen quality thickness with age. In a study by Benton & Brake (1996) it was found that the albumen quality at oviposition in breeder broilers decreased with an increase in flock age.
Pipping time of ostrich chicks gradually increased as females got older (P <0.05), and reached a peak for eggs laid by females at 8 to 9 years of age (Figure 6). The overall magnitude of this increase is very small (0.3 days) and it is uncertain whether this would be of any biological relevance. It is, however, worthwhile noting that our findings in this study differ from reports for both broilers and quails, in that the eggs from older hens were found to require less time to hatch than eggs of younger flock mates (Shanawany, 1984; Suarez et al., 1997; Yildirim, 2005). Butler (1991) found that the embryos of eggs laid by older broiler hens were more developed and at a more advanced stage compared with embryos from eggs produced by younger birds. For ostriches, there is only an indication that incubation time may decrease for eggs laid by females 10 years and older. For females of four years and younger, this can possibly be attributed to the fact that egg weight increases with female age, possibly resulting in an increased incubation time (Figure 4), since incubation time is positively correlated with egg weight (Wilson, 1991a).
A linear increase (P <0.050, Figure 7) occurred in pipping time from eggs that were set immediately (fresh eggs - 41.7 ± 0.1 d) to eggs stored for longer than seven days (42.2 ± 0.1 days). Wilson (1991b) and Yildirim (2005) also reported that ostrich eggs set on the day of laying required less time to incubate than eggs stored for longer periods. Accordingly, Tona et al. (2003) found that 18-day pre-incubation storage prolonged the incubation of broiler eggs by at least 15 h compared to eggs stored for only three days. In previous research it was shown that the embryonic survival of ostrich chicks was impaired in those eggs that were stored for seven days and longer (Brand et al., 2007). Thus prolonged egg storage appears to affect embryonic development that can result either in embryonic mortality or a delayed pipping time. A previous study by Brand et al. (2007) indicated that embryonic survival was compromised both in eggs set directly and in those stored for >7 days.
Ostrich eggs lost between 7.5 and 7.8% of their fresh weight up to 21 days of incubation (Table 3). By day 35 of incubation the average egg had lost between 12.2 and 12.7% of their fresh egg weight. Water loss at day 21 of incubation did not differ (P >0.05) for the different incubators. At 35 days of incubation a significant difference was found between the Africa International® incubator (12.7 ± 0.3% water loss) and the Natureform® incubator (12.0 ± 0.3%; P <0.05). Previous research suggested that eggs set in the former incubator were less likely to hatch than those incubated in the other incubators (Brand et al., 2007). Embryonic survival in the Africa International® incubator was compromised throughout incubation, although eggs set in this incubator were particularly vulnerable in the period from setting to 21 days. The less than optimal performance of the Africa International® incubator pertaining to hatching performance was attributed to a design that resulted in excessive temperature gradients within the incubator. It seems reasonable to assume that the same flaws that predispose eggs in this incubator to higher levels of embryonic deaths could be involved in the excessive water loss experienced. According to Brand et al. (2007), the overall hatching performance of the other incubators was markedly better than that of the African International® incubator, and all within acceptable bounds. It is not sure whether this relatively small difference could have contributed to the higher overall levels of embryonic mortality recorded for the former incubator by Brand et al. (2007). The Buckeye® and Prohatch® incubators did not differ significantly from the other incubators. The make of the incubator affected PT, with eggs incubated in the Buckeye® incubator pipping earlier than those incubated in the other incubators. It has to be stated that the magnitude of the difference between the Buckeye® and the other incubators was still very small.
Conclusion
Ostrich incubation traits were affected by a number of environmental factors, female age and genotype. These factors need to be considered during routine genetic evaluation. Failure to consider these factors will lead to biased genetic parameters and inaccurate breeding values. Year and season effects are often transient and unpredictable, as they may depend on ambient climate during a specific period. Yet cognisance should be taken of these effects, even if it is only to include it in genetic analyses to get rid of 'nuisance' variation. It is, however, important to consider the effects of genotype, female age, incubator type and setting, as well as storage time, as it has been shown that these effects have a marked influence on shell deaths in artificially incubated ostrich eggs (Brand et al., 2007). Combinations that are compatible with commercial ostrich production (i.e. the keeping of younger females, setting of eggs in functional incubators and the prevention of storage periods exceeding seven days) have the potential to improve commercial chick production. These factors should thus be considered when planning commercial ostrich husbandry and artificial incubation operations.
Acknowledgements
The authors thank the South African Department of Trade and Industry for partial funding of the work through their THRIP programme, as well as all those involved in the maintenance and recording of the research flock at the Oudtshoorn Research farm.
References
Ar, A., 1991. Egg water movements during incubation. In: Avian Incubation. Ed. Tullet, S.G., Poultry Science Symposium 22. Butterworth-Heinemann, London. pp. 157-173. [ Links ]
Ar, A., 1996. Requirements for successful artificial incubation of ostrich eggs. In: Improving our understanding of ratites in farming environment. Ed. Deeming, D.C., Proc. Int. Conf., 27-29 March 1996, Manchester, England. pp. 131-144. [ Links ]
Badley, A.R., 1997. Fertility, hatchability and incubation of ostrich (Struthio camelus) eggs. Poult. Avian Biol. Rev. 8, 53-76. [ Links ]
Benton, C.E. & Brake, J., 1996. The effect of broiler breeder flock age and length of egg storage on egg albumen during early incubation. Poult. Sci. 75, 1069-1075. [ Links ]
Brand, M.M., Cloete, S.W.P., Hoffman, L.C. & Muller, M., 2005. A comparison of live weights, body measurements and reproductive traits in Zimbabwean Blue ostriches (Struthio camelus australis) and South African Black ostriches (S. camelus var. domesticus). Proc. 3rd Int. Ratite Sci. Sym. & XII Wrld Ostrich Cong., 14-16th October 2005. Ed. Carbajo, E., Madrid, Spain. pp. 73-80. [ Links ]
Brand, Z., Cloete, S.W.P., Brown, C.R. & Malecki, I.A., 2007. Factors related to shell deaths during artificial incubation of ostrich eggs. J. S. Afr. Vet. Ass. 78, 195-200. [ Links ]
Brand, Z., Cloete, S.W.P., Malecki, I.A. & Brown, C.R., 2008. The genetic relationships between water loss and shell deaths in ostrich eggs, assessed as traits of the dam. S. Afr. J. Anim. Sci. (Submitted). [ Links ]
Blood, J.R., Van Schalkwyk, S.J., Cloete, S.W.P. & Brand, Z., 1998. Embryonic deaths in relation to water loss of artificially incubates ostrich eggs. Proc. 2nd Int. Ratite Cong., Oudtshoorn, South Africa. pp. 148-151. [ Links ]
Bunter, K.L., & Cloete, S.W.P., 2004. Genetic parameters for egg-, chick- and live-weight traits recorded in farmed ostriches (Struthio camelus). Livest. Prod. Sci. 91, 9-22. [ Links ]
Bunter, K.L, Cloete, S.W.P. & Van Schalkwyk, S.J., 1999. Significant genetic parameters for egg, chick and juvenile weight traits in ostriches. Proc. Ass. Advmt. Anim. Breed. Genet. 13, 476-479. [ Links ]
Butler, D.E., 1991. Egg handling and storage at farm and hatchery. In: Avian Incubation. Ed. Tullet, S.G., Butterworth-Heinemann Ltd. Surrey, UK. pp. 195-204. [ Links ]
Cloete, S.W.P., Bunter, K.L. & Van Schalkwyk, S.J., 2002. Progress towards a scientific breeding strategy for ostriches. Proc. 7th Wrld. Cong. Genet. Appl. Livest. Prod. 30, 561-568. 18-23 August, Montpellier, France. [ Links ]
Cloete, S.W.P., Bunter, K., Lambrechts, H., Brand, Z., Swart, D. & Greyling, J.P.C., 2006. Variance components for live weight, body measurements and reproductive traits of pair-mated ostrich females. Br. Poult. Sci. 47, 147-158. [ Links ]
Cloete, S.W.P., Lambrechts, H., Punt, K. & Brand, Z., 2001. Factors related to high levels of ostrich chick mortality from hatching to 90 days of age in an intensive rearing system. J. S. Afr. Vet. Ass. 72, 197-202. [ Links ]
Cloete, S.W.P., Van Schalkwyk, S.J. & Brand, Z., 1998. Ostrich breeding - progress towards a scientifically based strategy. Proc. 2nd Int. Ratite Cong., Oudtshoorn, South Africa. pp. 55-62. [ Links ]
Deeming, D.C., 1995. Factors affecting hatchability during commercial incubation of ostrich (Struthio camelus) eggs. Br. Poult. Sci. 36, 51-65. [ Links ]
Fasenko, G.M., Robinson, F.E., Hardin, R.T. & Wilson, J.L., 1992. Effect of duration of egg storage period. Poult. Sci. 72, 2129-2132. [ Links ]
Gilmour, A.R., Cullis, B.R., Welham, S.J. & Thompson, R., 1999. ASREML - Reference manual. NSW Agriculture Biometric Bulletin No. 3 NSW Agriculture, Orange Agriculture Institute, Forest Road, Orange 2800, NSW, Australia. [ Links ]
Glass, G.V., Peckham, P.D. & Sanders, J.R., 1972. Consequences of failure to meet assumptions underlying the fixed effects analyses of variance and covariance. In: Review of Educational Research. Vol. 42. No. 3. pp. 237-288. [ Links ]
Horbaňczuk, J.O., Sales, J., Celeda, T. & Zieba, G., 1999. Effect of relative humidity on the hatchability of ostrich (Struthio camelus) eggs. Czech. J. Anim. Sci. 44, 303-307. [ Links ]
Ipek, A. & Sahan, U., 2004. Effect of breeder age and breeding season on egg production and incubation in farmed ostriches. Poult. Sci. 45, 643-647. [ Links ]
Jarvis, M. J.F., 1998. The subspecies and races of ostriches and their present status in the wild. Proc. 2nd Int. Ratite Cong, Oudtshoorn, South Africa. pp. 4-8. [ Links ]
Lambrechts, H., 2004 Reproductive efficiency of ostriches (Struthio camelus). PhD Agric. dissertation, University of the Free State, Bloemfontein, South Africa. [ Links ]
Malecki, I.A., Horbaňczuk, J.O., Reed, C.E. & Martin, G.B., 2005. The ostrich (Struthio camelus) blastoderm and embryo development following storage of eggs at various temperatures. Br. Poult. Sci. 46, 652-660. [ Links ]
Meijerhof, R., 1992. Pre-incubation holing of hatchling eggs. Wrld. Poult. Sci. J. 48, 57-68. [ Links ]
Narahari, D., Mujeer, K.A., Ahmed, M., Rajini, R.A. & Sundararasu, V., 1991. Factors influencing the hatching performance of duck eggs. Br. Poult. Sci. 32, 313-318. [ Links ]
Petitte, J.N. & Davis, G., 1999. Breeding and Genetics. In: The ostrich - biology, production and health. Ed. Deeming, D.C., CABI Publishing, CAB International, Wallingford. pp. 275-292. [ Links ]
Shanawany, M.M., 1984. Interrelationship between egg weight, parental age and embryonic development. Br. Poult. Sci. 25, 449-455. [ Links ]
Stewart, J.S., 1995. Hatchery management in ostrich production. American Ostrich, November 1995. pp. 22-44. [ Links ]
Suarez, M.E., Wilson, H.R., Mather, F.B., Wilcox, C.J. & McPherson, B.E., 1997. Effects of strain and age of the broiler breeder female on incubation time and chick weight. Poult. Sci. 76, 1029-1036. [ Links ]
Swart, D. & Rahn, H., 1988. Microclimate of ostrich nests: measurements of egg temperature and nest humidity using egg hygrometers. J. Comp. Physiol. B. 157, 845-853. [ Links ]
Swart, D., Rahn, H. & De Kock, J., 1987. Nest microclimate and incubation water loss of eggs of the African ostrich (Struthio camelus var domesticus). J. Exp. Zool. (Suppl.) 1, 239-246. [ Links ]
Tona, K., Malheiros, R.D., Bamelis, F., Careghi, C., Moraes, V.M.B., Onagbesan, O., Decuypere, E. & Bruggeman, V., 2003 Effects of storage time on incubating egg gas pressure, thyroid hormones and corticosterone levels in embryos and on their hatching parameters. Poult. Sci. 82, 840-845. [ Links ]
Van Schalkwyk, S.J., Brand, Z., Cloete, S.W.P. & Blood, J.R., 1998. The influence of different disinfection protocols on the hatching performance of ostrich eggs. Proc. 2nd Int. Ratite Cong, Oudtshoorn, South Africa. pp. 157-159. [ Links ]
Van Schalkwyk, S.J., Brand, Z., Cloete, S.W.P. & Brown, C.R., 1999. Effects of time of egg collection and pre-incubation treatment on blastoderm development and embryonic mortality in ostrich embryos. S. Afr. J. Anim. Sci. 29, 154-163. [ Links ]
Wilson, H.R., 1991a. Interrelationship of egg size, chick size, posthatching growth and hatchability. Wrld's Poult. Sci. J. 47, 5-20. [ Links ]
Wilson, H.R., 1991b. Effect of egg size on hatchability, chick size and post-hatching growth. In: Avian Incubation. Ed. Tullet, S.G., Butterworth-Heinemann Ltd. Surrey, UK. pp. 279-283. [ Links ]
Yildirim, I., 2005. Effects of breeder age and pre-incubation storage of eggs on hatchability, time of hatch and relative organ weight of quail chicks at hatch. S. Afr. J. Anim. Sci. 35, 135-142. [ Links ]
 Correspondence:
Correspondence:
E-mail: zanellb@elsenburg.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}