










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.4 Pretoria Apr. 2008
Effect of dietary protein content on growth, uniformity and mortality of two commercial broiler strains
E.T. Berhe; R.M. Gous
Animal & Poultry Science, School of Agricultural Sciences and Agribusiness, University of KwaZulu-Natal, Private Bag X01, Scottsville 3209, South Africa
ABSTRACT
Two experiments were conducted to determine the response in performance, including uniformity and mortality, of two broiler strains to dietary protein content. In Experiment 1, 480 Cobb 500 and 480 Ross 788 day-old sexed broiler chickens were housed in cages to 21 d with 10 chickens per cage, and in Experiment 2, lasting 42 d, 1680 sexed broiler chickens of each of these two strains were placed in 48 floor pens with 70 chickens per pen. Males and females were reared separately in both experiments. Six levels of dietary protein were fed for 21 d in both experiments, the composition changing for the period 22 to 42 d in Experiment 2. Body weight of each bird was measured at weekly intervals up to 21 d in Experiment 1, and of 20 randomly selected birds from each pen on days 1, 21 and 42 in the second experiment. Broilers in the latter trial were group-weighed by pen at weekly intervals. Mortality was monitored daily. The highest body weight gains and feed conversion efficiencies (FCE, g gain/ kg food) were recorded in Cobb, with a correspondingly higher food intake for the starter feed in both experiments. In the finisher period Ross birds consumed significantly more than Cobb broilers (9 g/d) but in this case there was no difference in growth rate between the two strains, resulting in a significantly poorer FCE for Ross (487 vs. 522 g gain/kg feed). The pattern of food intake in the finisher period also differed between the two strains: the Cobb birds increased their food intake as the dietary protein content was decreased, but food intake decreased with protein content in the Ross. Uniformity was greatest in both strains when they were fed the highest protein feeds in both experiments, the variation in live weight increasing as the protein content decreased. There was no nutritional effect on mortality, although mean overall mortality was twice as high in Cobb broilers.
Keywords: Broiler, strain, uniformity, mortality, dietary protein
Introduction
Traditionally, the major criteria for assessing the performance of broiler strains have been growth rate and feed conversion efficiency (FCE), and less frequently, carcass composition (Cahaner et al., 1987; Cabel & Waldroup, 1991; Smith & Pesti, 1998; Rezaei et al., 2004), but there is circumstantial evidence that some strains show higher mortalities and a greater variability in final body weight than others when high or low-protein feeds are fed (Corzo et al., 2004; Kemp et al., 2005). Such information is crucial when optimising the feeds for different strains. In this study, therefore, flock uniformity and mortality rate were added to the growth variables studied.
Uniformity is an important measure of performance when optimising the feed and feeding programme of a population of broilers as it relates to the spread of product yield in the processing plant. Poor uniformity reduces revenue and increases waste, and optimisation routines therefore need to take account of any factors that may influence uniformity. The theory of food intake proposed by Emmans (1981) suggested that a bird will attempt to consume sufficient of a given food to enable it to grow at its potential, thus overconsuming energy if the feed is marginally deficient in an essential nutrient such as an amino acid. The performance of a broiler population depends largely on the inherent genotype of the population and the extent to which this varies between individuals (Emmans & Fisher, 1986). It is apparent that strains of broilers and individuals within a strain overconsume energy to different extents when faced with a marginally deficient feed (Corzo et al., 2004; Kemp et al., 2005). It is likely therefore that such feeds will affect uniformity, as some birds will eat more than others and benefit from the extra feed. Although evidence of a relationship between nutrient supply and mortality is somewhat equivocal, there is some evidence (Kemp et al., 2005) that suggests that this may be an important criterion to consider when optimising the feeds and feeding programme of broilers (Gous, 2007).
The objective of this study was to determine the extent to which food intake, growth, uniformity and mortality of two strains of broilers available in South Africa are influenced by dietary protein, with a view to incorporating such strain differences, either empirically or mechanistically, into a broiler growth model and optimiser.
Materials and Methods
Experiment 1: 480 Cobb 500 and 480 Ross 788 day-old sexed broiler chickens were housed in metabolism cages on Ukulinga Research Farm, University of KwaZulu-Natal. Ninety-six cages were used with 10 chickens per cage, and with males and females being reared separately. Two nipple drinkers were available in each cage. Birds were supplied with heat by means of a gas heater/blower. Body weight of individual birds in each pen was measured at weekly intervals up to 21 d.
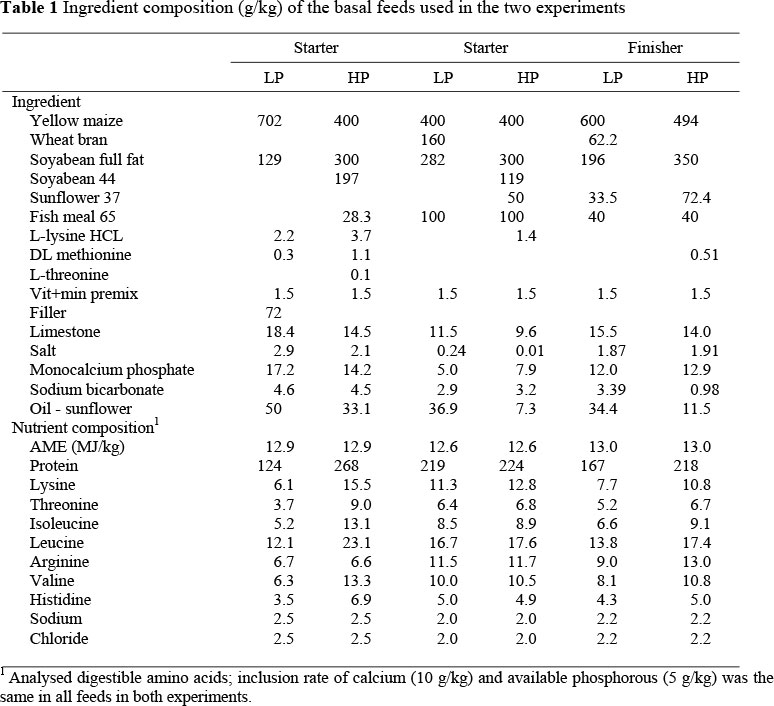
Two basal feeds, one high (H) and the other low (L) in protein were formulated to contain equal contents of metabolisable energy (ME) and major minerals (Table 1), and with amino acids conforming to the starter recommendations of Aviagen (2002). In formulating these feeds, crude protein and not cost was minimised, thereby producing the best possible amino acid balance with the ingredients available. The two basal feeds were fed alone and as blends (20H:80L, 40H:60L, 60H:40L, 80H:20L) thereby producing six levels of balanced protein in total. Feed troughs incorporating feed-saver grids were placed inside the cages for the first 10 days, and thereafter, outside the cages. The two basal feeds were sampled after mixing, and these samples were analysed for apparent metabolisable energy (AME), crude protein (CP) and amino acid content (Table 1). Feed and water were available ad libitum throughout the trial.
Experiment 2: The broiler strains used were the same as those used in the first trial, with 1680 chicks of each strain being placed at day old in 48 floor pens (1.5 m x 2 m), sexes separate, accommodating 70 birds per pen. The experiment was terminated when the broilers were 42 d. The temperature in the tunnel-ventilated broiler house was maintained at close to predetermined levels, starting at 31 °C and reducing to 21 °C by 21 d, then maintained at 21 °C until 42 d. Two basal feeds were formulated for the starter and the finisher periods (Table 1) based on the Aviagen (2002) requirements and these were blended in the same proportions as those in Experiment 1. The starter feeds were fed in a mash form whilst the grower feeds (fed from 22 to 42 d) were pelleted.
Both experiments followed a randomised blocks design with a factorial arrangement of treatments (Feed x Strain x Sex). Each treatment was replicated four times in Experiment 1 and twice in Experiment 2. Food intake was measured weekly in both experiments. In Experiment 2, all the surviving birds were group-weighed by pen at weekly intervals and 20 broilers were randomly selected from each pen and weighed individually on days 1, 21 and 42 to determine variation between individuals. Mortality was recorded whenever this occurred. Food intake was adjusted for mortality. Mortality percentage was transformed to angular values for analysis. Data were subjected to statistical analysis using analysis of variance and regression procedures (simple linear regression with groups) of Genstat (2005) in order to compare the treatment effects and to determine the response in the measures of performance to dietary protein content.
Results
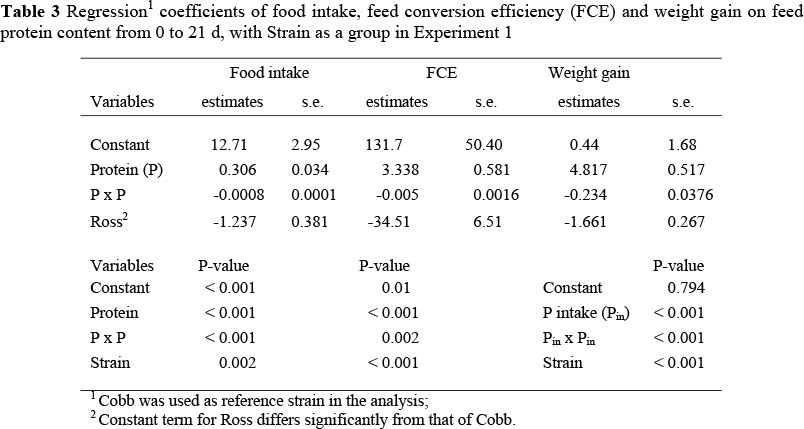
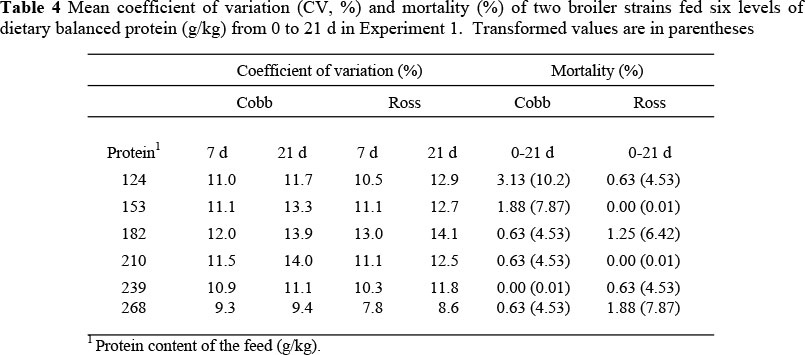
Experiment 1: Mean weight gain, food intake and FCE were greater (P <0.001) in the Cobb strain over both sexes and all feed treatments (Table 2) even though the mean differences between strains were small (2.0 and 1.2 g/d for weight gain and food intake respectively, and 34 g gain/kg feed), and for females over both strains and feed treatments (P >0.001). Each of these performance variables was quadratically related (P <0.001) to dietary protein content (Table 3), with no significant interactions evident between the three factors, i.e. the slopes of the responses to protein between the two strains were the same, although the intercepts differed significantly between strains (Table 3). Uniformity was highest (variation being lowest) at the two extremes of protein content, and lowest with the intermediate treatments (Table 4). Mortality during the trial was low (<1.0%); however, there was a higher overall mortality in the Cobb strain (1.15 vs. 0.73%), especially at low protein contents, but this trend was not apparent in the Ross strain across feed protein contents.
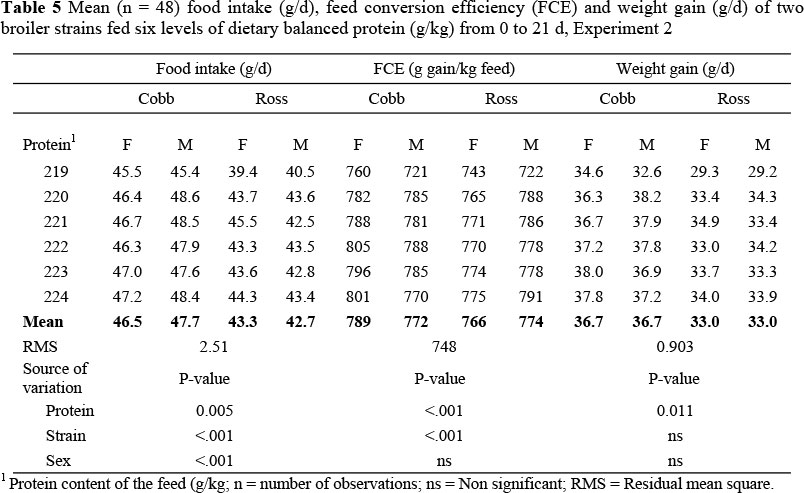
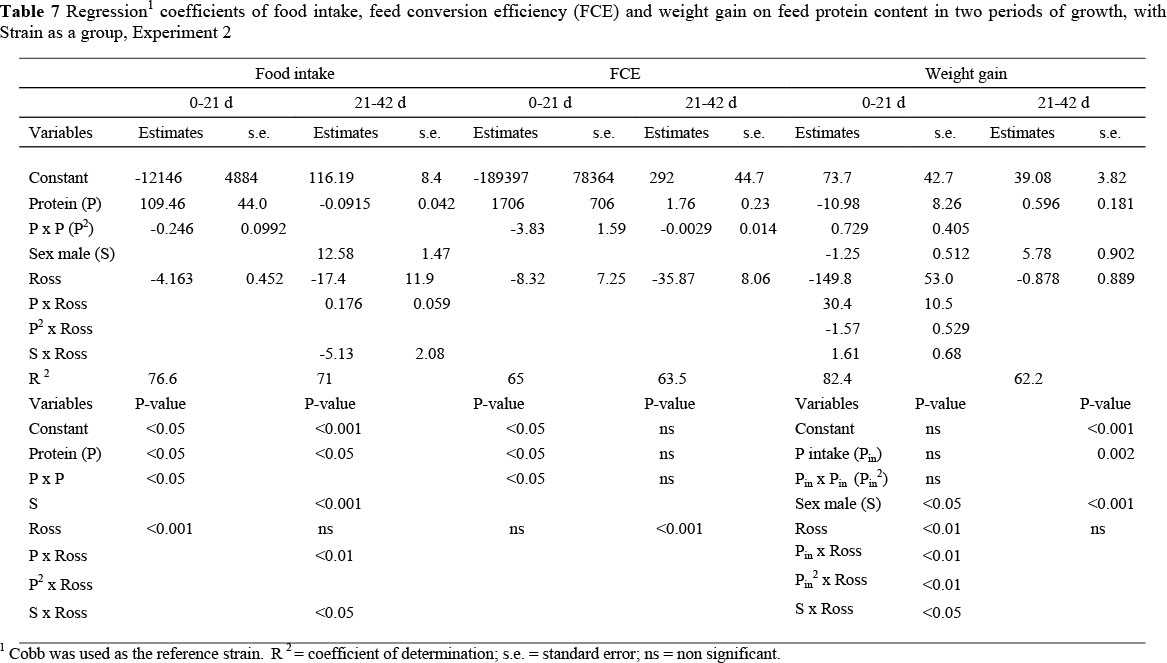
Experiment 2: The dietary crude protein contents of the basal feeds used in the starter period of this trial were not as widely different as in the first trial, nor were they as specified due to an apparent mixing error in the commercial feed mill where they were made. Instead of the protein contents being 197 and 248 g protein/kg feed, the analysed values were 219 and 224 g/kg. Nevertheless, the effect of dietary protein content on the growth, uniformity and mortality in the period to 21 d exhibited a similar pattern to that in the same period in Experiment 1 (Table 5). Cobb broilers consumed more food (4.1 g/d; P <0.001) and grew faster (3.7 g/d; P <0.001) than the Ross strain in the starter period although the response in food intake to dietary protein content was the same in both strains, namely, remaining constant on all but the lowest protein feed, when intake declined in all cases. FCE was significantly higher (23 g gain/kg feed) among Cobb females than Ross females (Tables 5 and 7), but was the same in males of the two strains. In both strains FCE decreased almost linearly (the second order term was significant, but very small compared with the linear term, Table 7) as the dietary protein content was reduced.
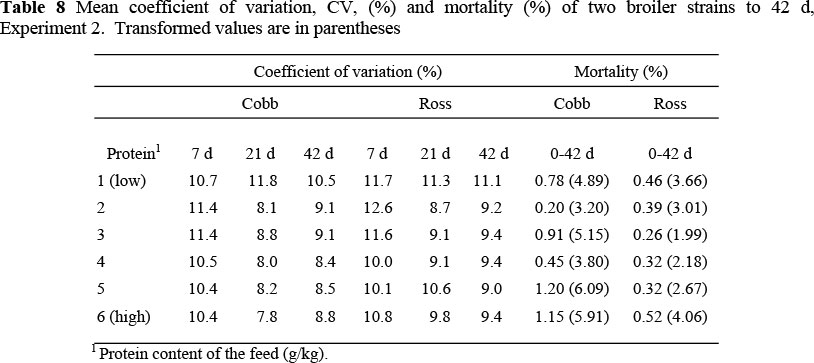
In the finisher period Ross consumed more food than Cobb broilers (9 g/d; P <0.001) (Table 6) but in this case there was no difference in growth rate between the two strains, resulting in a poorer FCE for the Ross (487 vs. 522 g gain/kg feed; P <0.001). The pattern of food intake in this period differed significantly between the two strains (P <0.001) (Table 7); Cobb birds increased their food intake as the dietary protein content was decreased, whilst food intake decreased with protein content in the Ross strain. Regression coefficients of food intake, feed conversion efficiency (FCE) and weight gain on feed protein content are presented in Table 7. Variation in live weight within a treatment decreased in both strains as the feed protein content increased. There was no nutritional effect on mortality (Table 8).
Discussion
Simulation models have been developed that predict food intake and growth of broilers (EFG Software, 2007) and are becoming increasingly sophisticated in that it is now possible to optimise the feeds and feeding programme of broilers of a given genotype in a given environment, given the cost and availability of raw materials and the value of the product being sold. Various objective functions may be either maximised or minimised in order to obtain the optimum solution. The consequence of using such an optimiser is that the solutions often move away from the conventional feeding strategy used by most nutritionists and breeding companies. For example, when cereal prices rise during periods of drought, or as is predicted to occur with the increased production of biofuels, the optimum strategy is to use feeds of lower nutrient density. Similarly, when the costs of protein sources increase, the optimal strategy, assuming that there is not a concomitant rise in the value of the product, would be to lower the protein content of the feeds. Nutritionists are loath to alter feed specifications partly because it is so difficult to predict what the consequences of such a strategy would be, and because it is even more difficult to determine the extent to which the feed should be altered in order to maximise profit. Given that simulation models are likely to be used to a greater extent in the future to optimise feeding strategies, it is of considerable practical significance that these optimisers should account accurately for all aspects of the production processes that influence profitability, including the effects of feed composition on mortality and uniformity. Strain differences too have been shown to exist, in their response to dietary balanced protein, and simulation models must be able to account for these differences. It was with this in mind that the two trials reported here were conducted.
Both weight gain and FCE decreased with dietary balanced protein content in both broiler strains and in both experiments reported here. Although no carcass analyses were conducted on the birds at the end of the trials, the lipid content of birds would have increased as protein supply was decreased (Gous et al., 1990) accounting for the relatively small decrease in body weight as dietary protein content was reduced. Thus, although the differences in weight at the termination of the trial were small, carcass composition would have varied to a greater extent, and this would impact on revenue in markets demanding a lean broiler.
In both trials, Cobb broilers performed better than the Ross strain in the starter period (0 to 21 d), over the range of protein levels used. This was in part due to the capacity of Cobb birds to consume greater quantities of feed at all protein contents, resulting in higher intakes and hence greater weight gains and FCE's than with the Ross strain on all feeds. The effect of feed protein content on food intake was of a quadratic form and almost identical in both strains, with intake first increasing as protein content was reduced, and then decreasing. This tendency to increase food intake as the limiting nutrient (in this case, protein) is reduced is a corollary to the theory of food intake regulation of Emmans (1981) incorporated into the EFG Broiler Growth Model (EFG Software, 2007) and has been confirmed in experiments by Gous et al. (1990), Skinner et al. (1992), Smith & Pesti (1998) and Smith et al. (1998). However, in the finisher period, the two strains differed in their response to dietary balanced protein, confirming the similar observation by Kemp et al. (2005). As in this earlier report, Cobb broilers exhibited the conventional increase in food intake as dietary protein was reduced, but intake by the Ross birds was reduced (Table 7), which is contrary to Emmans' (1981) theory of food intake regulation.
The Ross strain has been selected for improved growth and feed efficiency using high protein feeds. Such selection results in leaner carcasses (Pym & Solvens, 1979) and perhaps a reduced ability to fatten when faced with feeds marginally deficient in an essential nutrient. If this were the case then such strains would be incapable of increasing food intake on marginally deficient feeds, as a prerequisite for this is to be able to store the excess energy consumed as body lipid unless the environmental temperature were sufficiently low as to allow the birds to lose the excess energy as heat. It is evident from Table 6 that the Ross strain did not deposit as much body lipid as did the Cobb strain: the difference between the mean FCE of the two highest and the two lowest protein contents was 54 and 39 g/kg for male and female Cobb broilers respectively, and only 15 and -4 for the Ross, indicating that the Ross broilers, in changing the FCE only marginally as protein content was reduced, deposited the same proportions of body protein and lipid at all dietary protein contents, whereas the Cobb birds deposited considerably more lipid on the lower than on the higher protein feeds. Differences in growth rate over the range of protein contents used, calculated as for FCE above, were virtually the same in both strains (5 and 1 g/d for Cobb males and females respectively vs. 4 and 1 for the Ross strain) adding more weight to this argument. In the EFG Broiler Growth Model, the potential growth rate is described using a Gompertz growth curve (three parameters) and the lipid-to-protein ratio at maturity is an additional parameter used to describe a genotype. These are insufficient to describe the differences in food intake in the latter part of the growing period in response to dietary balanced protein observed by Kemp et al. (2005) and here. Some theoretical arguments are put forward by Gous (2007) to explain these differences, but as yet no satisfactory solution to the problem has been found.
An increasingly important characteristic of broiler production is the uniformity of body weight and conformation of birds when harvested, since consumers have become more sophisticated, demanding highly uniform whole birds and portions. There is evidence that uniformity decreases as the feed becomes marginally deficient in protein (Corzo et al., 2004) and this may be simulated using a population model such as that described by Gous & Berhe (2006) in which the performance of each individual in the population is simulated and the responses averaged. The probable cause of decreased uniformity on feeds marginally deficient in an essential nutrient is the variation in the ability of broilers to overconsume energy when faced with a deficiency. This characteristic will enable the successful birds to show little reduction in growth, whereas the food intake of others, without the capacity to deposit as much lipid, could be severely constrained, resulting in poor growth. The simulation exercise above suggests that at both high and low (limiting) concentrations of dietary protein uniformity increases, the requirements of all individuals in the one case being met, and in the other, all individuals are similarly constrained. At the lowest protein levels, uniformity in the first experiment was higher than on the intermediate protein levels, probably because all birds were equally constrained by the severely deficient protein content of the feed offered. Because the feeds were not as deficient in protein in the second trial, i.e. none was severely deficient, uniformity diminished linearly throughout the range of protein levels used. Uniformity is clearly compromised as dietary protein content is reduced, a situation that is likely to occur frequently in commercial broiler operations. Thus, uniformity in a population of birds, caused by feeds varying in essential nutrient content, may be simulated mechanistically.
Mortality did not appear to be related to the nutrition treatments imposed on the Ross strain in either of the experiments, although the difference between strains was statistically significant (P <0.01) only in Experiment 2. In both experiments the Cobb birds showed a tendency to a higher mortality at the lowest dietary protein contents, and in the second experiment mortality increased exponentially on the highest feed protein contents, which conforms to the results of Rezaei et al. (2004) and Kemp et al. (2005), and circumstantial evidence from the field that has been ascribed to the faster growth of the Cobb broiler relative to Ross. Classen (2000) suggested that rapid growth has produced problems such as skeletal and cardiovascular disease (sudden death syndrome and ascites) not seen in slower growing birds. The stage of growth at which mortality occurs is of considerable importance when evaluating the effects of mortality on profitability: the older the birds are when they die, the more costly this becomes to the enterprise.
Broiler breeder companies sometimes suggest modifications to the lighting and feeding programmes as a means of reducing mortality in commercial broiler flocks. Some lighting programmes have been shown to reduce mortality in fast-growing strains of broilers (Classen & Riddle, 1989) and indeed may eventually become obligatory in reducing the incidence of metabolic disorders if these problems are not adequately addressed by the breeding companies involved. In only the second experiment did it appear that high dietary protein was implicated in the higher incidence of mortality in the Cobb strain, and one might draw the conclusion from this that modifications to the feeding programme are unnecessary or ineffective in this regard. However, in the experiment conducted by Kemp et al. (2005), the mortality of one of the strains used was significantly influenced by the protein content of the feed, increasing linearly from 8% on the lowest protein feed to 16% on the highest, whilst mortality remained constant, at 3%, on all protein levels in the other strain: clearly, it is not economically feasible to feed high protein feeds to both strains. Once an accurate estimate of the effects of protein content on uniformity and mortality within each of the two strains are known, such information would be usefully included in models designed to optimise the feeds and feeding programmes of broilers.
Conclusion
When determining the dietary balanced protein contents that will optimise performance in a flock of broilers, account needs to be taken of all the variables that are influenced by the composition of the feed, including food intake, uniformity and mortality, all of which were shown here to have been influenced by feed protein level. Although the two strains used in these trials exhibited similar responses to dietary protein content to 21 d, differences in response in food intake in the finisher period were of such a magnitude that the feed protein levels chosen to optimise performance are unlikely to be the same in both strains. The response in mortality was also shown to differ between the two strains, whereas uniformity was similarly affected between strains but differed according to the dietary protein content offered. Simulation models may be used to predict the responses in food intake and uniformity mechanistically, but the effect of feed protein on mortality would necessarily need to be modelled empirically until the reasons for the observed differences between strains are understood.
References
Aviagen, 2002. Ross Broiler Management Manual. Aviagen Ltd., Newbridge, UK. [ Links ]
Cabel, M.C. & Waldroup, P.W., 1991. Effect of dietary protein level and length of feeding on performance feeding on performance and abdominal fat content of broiler chickens. Poult. Sci. 70, 1550-1558. [ Links ]
Cahaner, A., Dunnington, E.A., Jones, D.E., Cherry, J.A. & Siegel, P.B., 1987. Evaluation of two commercial broiler male lines differing in efficiency of feed utilization. Poult. Sci 66, 1101-1110. [ Links ]
Classen, H.L., 2000. Managing metabolic disease in rapidly growing strains of poultry. In: The Challenge of Genetic Change in Animal Production. Eds Hill, W.G., Bishop, S.C., McGuirkay, B., Simm, G. & Webb, A.J., BSAS Occasional Publication No. 27, Edinburgh, UK. British Society of Animal Science. pp. 63-65. [ Links ]
Classen, H.L. & Riddle, C., 1989. Photoperiodic effects on performance and leg abnormalities in broiler chickens. Poult. Sci., 68, 873-879. [ Links ]
Corzo, A., McDaniel, C.D., Kidd, M.T., Miller, E.R., Boren, B.B. & Fancher, B.I., 2004. Impact of dietary amino acid concentration on growth, carcass yield, and uniformity of broilers. Aus. J. Agric. Res. 55, 1133-1138. [ Links ]
Dozier III, W.A. & Moran Jr., E.T., 2001. Response of early- and late-developing broilers to nutritionally adequate and restrictive feeding regimens during the summer. J. Appl. Poult. Res. 10, 92-98. [ Links ]
EFG Software, 2007. Broiler growth model and optimiser. www.efgsoftware.net (accessed April 2007). [ Links ]
Emmans, G.C., 1981. A model of the growth and feed intake of ad libitum fed animals, particularly poultry. In: Computers in Animal Production. Occasional Publication No. 5. British Society of Animal Production. pp.103 - 110. [ Links ]
Emmans, G.C. & Fisher, C., 1986. Problems of nutritional theory. In: Nutritional Requirements and Nutritional Theory. Eds Fisher, C. & Boorman, K.N., London, Butterworths. pp. 9-39. [ Links ]
GenStat executable, 2005. GenStat statistical software, Release 8.1 Lawes Agricultural Trust. [ Links ]
Gous, R.M., 2007. Predicting nutrient responses in poultry: future challenges. Animal 1, 57-65. [ Links ]
Gous, R.M. & Berhe, E.T. 2006. Modelling populations for purposes of optimisation. In: Mechanistic Modelling in Pig and Poultry Production. Eds Gous, R.M., Morris, T.R. & Fisher, C., CABI, Wallingford, UK. pp. 76-96. [ Links ]
Gous, R.M., Emmans, G.C., Broadbent, L.A. & Fisher, C., 1990. Nutritional effects on the growth and fatness of broilers. Br. Poult. Sci., 31, 495-505. [ Links ]
Kemp, C., Fisher, C. & Kenny, M., 2005. Genotype - nutrition interactions in broilers; response to balanced protein in two commercial strains. 15th European Symp. Poultry Nutrition, Balatonfured, Hungary. [ Links ]
Orr, H.L., Hunt, E.C. & Randall, C.J., 1984. Yield of carcass, parts, meat, skin, and bone of eight strains of broilers. Poult. Sci. 63, 2197-220. [ Links ]
Pym, R.A.E. & Solvyns, A.J., 1979. Selection for food conversion in broilers: body composition of birds selected for increased body-weight gain, food consumption and food conversion ratio. Br. Poult. Sci. 20, 87-97. [ Links ]
Rezaei, M., Nassiri, H., Moghaddam, Pour Reza, J. & Kermanshahi, H., 2004. The effect of dietary protein and lysine levels on broiler performance, carcass characteristics and N. excretion. Int. J. Poult. Sci. 3 (2), 148-152. [ Links ]
Skinner, J.T., Waldroup, A.L. & Waldroup, P.W., 1992. Effects of dietary amino acid levels and duration of finisher period on performance and carcass content of broilers forty-nine days of age. Poult. Sci. 71, 1207-1214. [ Links ]
Smith, E.R. & Pesti, G.M., 1998. Influence of broiler strain cross and dietary protein on the performance of broilers. Poult. Sci. 77, 276-281. [ Links ]
Smith, E.R., Pesti, G.M., Bakalli, R.I., Ware, G.O. & Menten, J.F.M., 1998. Further studies on the influence of genotype and dietary protein on the performance of broilers. Poult. Sci. 77, 1678-1687. [ Links ]
 Correspondence:
Correspondence:
E-mail: gous@ukzn.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}