










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 no.3 Pretoria mar. 2008
Effects of L-carnitine in layer diets containing different fat sources and energy levels on hen performance and egg quality
M. CordukI; S. SaricaII
IDepartment of Animal Science, Faculty of Agriculture, University of Ankara, 06110 Ankara, Turkey
IIDepartment of Animal Science, Faculty of Agriculture, University of Gaziosmanpasa, 60240 Tokat, Turkey
ABSTRACT
The study was conducted to determine the effects of feeding diets containing two levels of metabolisable energy (normal or low) and three sources of fat (palm-, sunflower- or fish-oil) with or without L-carnitine (0 or 500 mg/kg diet) on the performance and egg quality parameters of Brown egg-type laying hens. The hens receiving the low energy diet had significantly higher body weights and feed intakes (FI) than the birds on the normal energy diet, but their feed conversion ratio (FCR) was poorer. L-carnitine in the diet containing sunflower oil (SO) increased FI significantly, resulting in a poorer FCR. However, the FI and FCR were higher when the fish oil (FO) was added to diets without L-carnitine compared to the other diets. The palm oil (PO) supplemented diet without L-carnitine significantly decreased the specific gravity of the eggs. L-carnitine inclusion into the diet containing PO significantly increased the Haugh unit and albumen index of the eggs, while the inclusion of FO in the low energy diet increased the Haugh unit and albumen index significantly. However, when PO was included these parameters decreased significantly. The addition of L-carnitine to the normal energy diet reduced the pH of the egg yolk, while it increased egg yolk pH when added to the low energy diet. The inclusion of FO in the low energy diet resulted in a significant increase in yolk pH. Albumen pH was significantly decreased by dietary L-carnitine supplementation, but increased when the diet low in energy contained FO, as well as the normal energy diet containing SO. The inclusion of FO to both the normal and low energy diets, except the low energy diet with L-carnitine, improved the breaking strength of the eggshells. Similarly, SO and PO inclusion to low energy diets with L-carnitine increased eggshell breaking strength. The supplementation of PO and L-carnitine to the diet enhanced the Haugh unit and albumen index. Palm oil and the normal energy diet decreased egg yolk indices.
Keywords: Layers, sunflower oil, fish oil, palm oil, metabolisable energy, carnitine, egg traits
Introduction
L-carnitine (ß-OH-y-N-trimethylaminobutyric acid) is a water-soluble quaternary amine with a low molecular weight, and occurs naturally in microorganisms, plants and animals (Bremer, 1983). L-carnitine is biosynthesised in vivo from the essential amino acids, lysine and methionine, in the kidneys and liver in the presence of ferrous ions. The three vitamins, ascorbate, niacin and pyridoxine, are required as cofactors for the enzymes involved in the metabolic pathway of L-carnitine synthesis (Bieber, 1988; Mast et al., 2000).
L-carnitine has antioxidative properties. The major metabolic role of L-carnitine is reducing the availability of lipids for peroxidation by facilitating the transport of long-chain fatty acids across the inner mitochondrial membrane for ß-oxidation. Thus, dietary L-carnitine supplementation promotes the β-oxidation of these fatty acids to generate adenosine triphosphate (ATP) energy and improves energy utilisation (Neuman et al., 2002). Rabie et al. (1997) observed that L-carnitine (500 mg/kg diet) did not influence the laying performance of hens, nor external egg quality, yolk index (YI) or yolk colour score of eggs. However, albumen quality (albumen height (AH) and Haugh unit (HU)) was improved by supplemental dietary L-carnitine. They concluded that L-carnitine has a beneficial effect on albumen quality and could modify the components of the edible part of the egg during the late laying period. Leibetseder (1995) pointed out that the supplementation of a standard layer diet with either 500 mg L-carnitine or nicotinic acid, or a combination of the two substances had neither beneficial nor detrimental effects on body weight (BW), feed intake (FI), egg production (EP) and concentrations of serum and yolk cholesterol during the early laying period. However, the L-carnitine content of yolks was significantly increased in supplemented groups. Celik et al. (2004) reported that body weight change (BWC), FI, EP and some external egg qualities were not affected by L-carnitine supplementation although relative albumen weight (AW) and height (AH) were significantly increased by supplemental L-carnitine.
There does not seem to be literature dealing with the effects on performance and egg quality parameters of laying hens receiving diets containing L-carnitine and different fat sources and energy levels.
The present study was planned to determine the effects of feeding diets at two levels of metabolisable energy and three sources of fat with or without supplemental L-carnitine on the performance and egg quality parameters of Brown egg-type laying hens.
Materials and Methods
One hundred and eighty Isabrown laying hens, 27 weeks old, were randomly allocated to laying cages. The birds were assigned to 12 dietary treatments replicated five times with three hens per replicate. In a 2 x 2 x 3 factorial arrangement, laying hens were fed diets containing two levels of metabolisable energy (ME) (11.51 or 10.88 MJ ME/kg) and three sources of fat, palm (PO)-, sunflower (SO)- or fish-oil (FO), with or without supplemental L-carnitine (0 or 500 mg/kg diet) in the form of Carniking (LONZA Ltd., Basel, Switzerland). The L-carnitine content of the diets was calculated based on the L-carnitine content of the ingredients used (Harmeyer et al., 1998). The composition and the calculated nutrient content of the experimental diets are presented in Table 1. Prior to diet formulation, feed ingredients were analyzed for their dry matter, crude ash, crude protein (CP) (N x 6.25), crude fat, calcium, starch and total sugar content, according to the methods of the Association of Analytical Chemists (AOAC, 2007). The AOAC numbers for dry matter, crude ash, crude protein, crude fat, calcium, starch and total sugar laboratory techniques were 934.01, 942.05, 988.05, 920.39, 927.02, 920.40 and 974.06, respectively. The ME levels ofthe feed ingredients were calculated based on the analyzed values of the feedstuffs (WPSA, 1989). Except for energy level, all diets were formulated to meet the minimum nutrient requirements for laying hens, as recommended in the Isabrown Technical Handbook (Anonymous, 2000a). During the feeding period that lasted 84 days, the experimental diets in mash form and drinking water were provided ad libitum. The photoperiod was set at 16L : 8D throughout the experimental period. All birds were weighed at the start and end of the experiment. Feed intake (FI) was recorded weekly and EP and egg weight (EW) were recorded daily. Performance characteristics (FI, EP, EW, egg mass (EM), feed conversion ratio (FCR) and mortality), egg internal qualities (HU, yolk index (YI) and albumen index (AI), yolk and albumen pH) and external qualities (egg shell thickness (EST), egg specific gravity (ESG) and percentage of cracked eggs (CE)) were evaluated three times during the 84-d experiment period. All eggs laid during the 26 to 28 d period were collected for these measurements. The internal and external parameters of egg were measured for three consecutive days. Body weight, BWC, FI, EM, EW, EP, FCR (g FI/g EM), CE ratio, ESG, albumen and yolk pH, shell breaking strength (SBS), EST, AI, YI and HU (Rauch, 1965; Card & Nesheim, 1972) were calculated throughout the experiment. The ESG of a whole egg was measured using the Archimedes' method with an instrument designed for the measurement of egg weight (EW) in air (EWa) and in water (EWw) and ESG was calculated using the following formula [ESG= EWa/(EWa-EWw)] (Hamilton, 1982; Hempe et al., 1988). Shell breaking strength was measured using an instrument (Dr. ING. George Wasna Mess+Prüftechnick, Berlin) that assessed the resistance of the egg to crushing (kg/cm2). Eggshell thickness was measured using a micrometer (Mitutoya, 0.01 mm, Japan). Albumen height (H) was measured using a tripod micrometer (Mitutoya, 0.01 mm, Japan), albumen length (AL) and albumen width (AW) using a compass (SMI, 0.1 mm, Italy) and then AI was calculated using the following formula [AI=AH/(AL+AW/2) x 100]. Yolk height (YH) was measured using a tripod micrometer (Mitutoya, 0.01 mm, Japan) and yolk diameter (YD) using a compass. The YI was calculated using the following formula [YI= (YH/YD)x100].
The HU was calculated using the records of AH and EW and employing the formula: [HU = 100 x Log (AH+7.57-1.7 EW0.37)]. For yolk and albumen pH measurements, eggs were broken and separated into yolk and albumen. The albumen and yolk samples were stirred with a glass rod during pH measurements (Hanna, Instrument, HI 9321 microprocessor pH meter, Portugal).
The data obtained from the experiment were analyzed statistically using the General Linear Model Procedure of the SPSSWIN (1994). Significant differences between treatment means were separated using Duncan's Multiple Range Test with a 5% probability (Duncan, 1955).
Results and Discussion
There were no significant differences in BW (1816 ± 14.19 g), EW (59.31 ± 1.09 g) or EP (97.25 ± 1.69%) of the laying hens between treatments at the beginning of the experiment. No mortalities were recorded over the total feeding period.
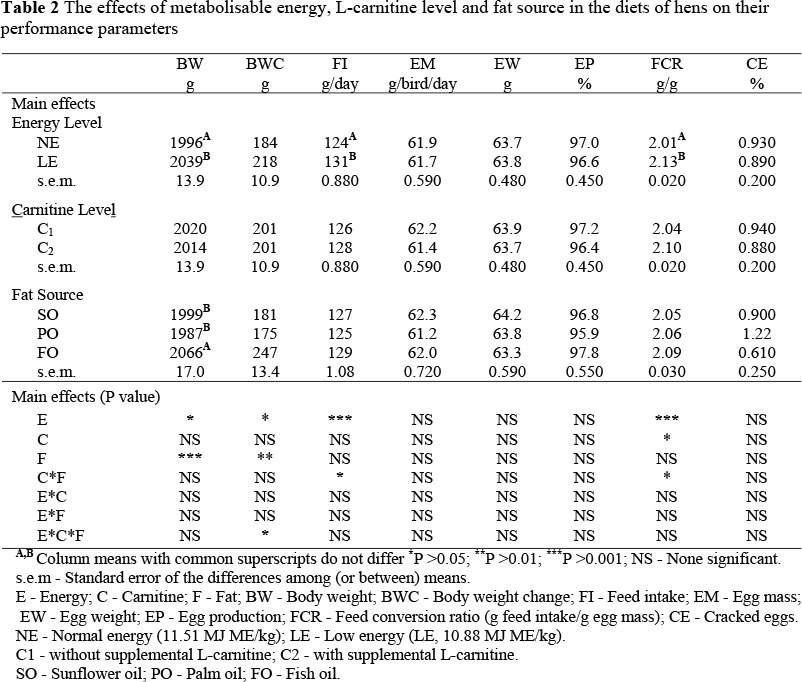
The effects of dietary treatments on performance parameters are presented in Table 2. Metabolisable energy and L-carnitine levels and fat sources did not affect EM, EW, EP and CE significantly.
Feeding the diet low in energy (LE) significantly increased BW and FI, but resulted in poorer FCR compared to those of hens receiving the normal energy (NE) diet. The level of energy intake of the laying hens was compensated for, with the increased FI. The dietary fat intake and net energy intake were probably increased due to the increased FI in the LE diet. Bish et al. (1985) observed that when fat replaced starch as a source of energy, the net energy available to the bird was enhanced, and this could result in an increase in the BW of hens. The effect of fat sources on the body weight of hens differed between treatments (P <0.001) (Table 2). The BW of hens fed a diet containing FO was significantly higher than that of hens fed diets with SO and PO. Being rich in polyunsaturated fatty acids (PUFAs), FO might have stimulated the increase in BW (Hu et al., 2002).
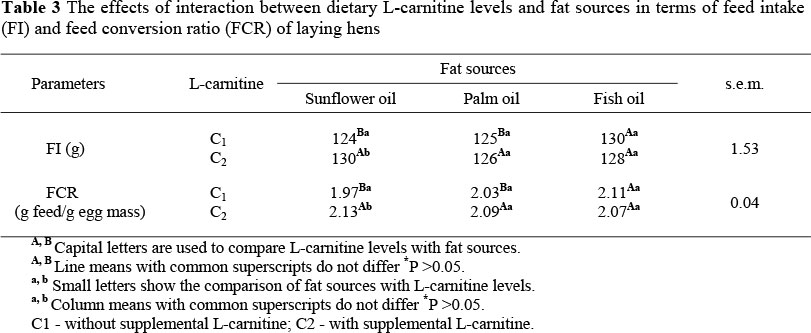
The significant interaction between L-carnitine levels and dietary fat sources in terms of FI and FCR is presented in Table 3. The use of FO in the diet of the hens without L-carnitine significantly increased the FI and worsened the FCR compared to the diet containing SO or PO. However, the effect of fat sources on the FI was not significant due to L-carnitine supplementation. L-carnitine supplementation to the diets promotes the mitochondrial ß-oxidation of long-chain fatty acids (LCFAs) by facilitating their transfer across the inner mitochondrial membrane. Thus, supplemental L-carnitine could improve fatty acid and energy utilization (Mast et al., 2000). A non-profitable conversion of dietary energy due to a deficiency of L-carnitine in the diet containing a fat source rich in long-chain fatty acids might have increased the FI and worsened the FCR of the hens. The L-carnitine supplementation to the diet containing SO significantly increased the FI and resulted in a poorer FCR compared to a diet without L-carnitine.
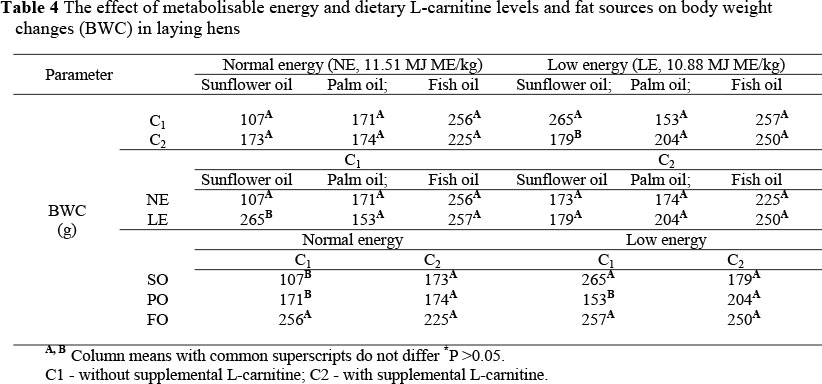
The interaction between metabolisable energy and L-carnitine levels and fat sources in terms of body weight change (BWC) (P <0.05) is summarized in Table 4. Supplementing the LE diet containing SO with L-carnitine significantly decreased BWC compared to the diet without L-carnitine (Table 4). In addition, the BWC of hens fed LE diet containing SO and without L-carnitine was significantly higher than that of hens receiving the NE diet (Table 4).
The BWC of hens fed a diet with LE and containing SO without L-carnitine was significantly higher than that of hens fed a diet with supplemental L-carnitine. Consequently, L-carnitine supplementation to diets reduced the amount of LCFAs availability for esterification to triacylglyserols and storage fat in adipose tissue (Barker & Sell, 1994; Xu et al., 2003). Thus, dietary L-carnitine supplementation promotes the ß-oxidation of these fatty acids to generate adenosine triphosphate (ATP) energy and improves energy utilisation (Rabie et al., 1997; Neuman et al., 2002). In addition, the use of SO or PO in diets with a NE and without L-carnitine significantly reduced BWC compared to the diet containing FO. However, feeding the diet with LE containing SO or FO and without supplemental L-carnitine significantly increased BWC compared to that of hens fed the diet that contained PO (Table 4).
Palm oil is rich in the saturated fatty acid, palmitic acid (C16:0), constituting about 45% of total fatty acids. Saturated fats rich in LCFAs (>14C atoms) are less digestible than fats high in medium-chain fatty acids or unsaturated fatty acids (Vila & Esteve-Garcia, 1996). It has been shown that a large percentage of saturated fatty acids in chicken excreta is in the form of unutilized soap in contrast to that of unsaturated fatty acids (Atteh & Leeson, 1983). There is evidence that saturated fatty acids, particularly palmitic and stearic acid, are less efficiently utilized compared to unsaturated fatty acids (Renner & Hill, 1961;Young & Garrett, 1963). In addition, a high fraction of C16:0 in PO is bound at the sn-1 or sn-3 position of the glycerol molecule (Mu & Hoy, 2004). Long-chain saturated fatty acids bound at the sn-1 and sn-3 positions are absorbed less efficiently than those bound at the sn-2 position (Smink et al., 2008).
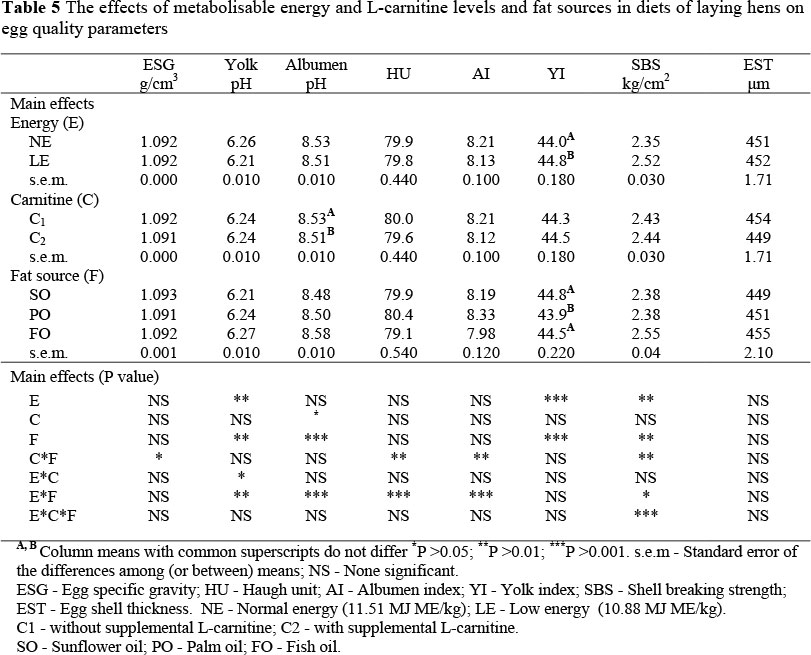
The effects of metabolisable energy and L-carnitine levels and fat sources on egg quality parameters are summarized in Table 5.
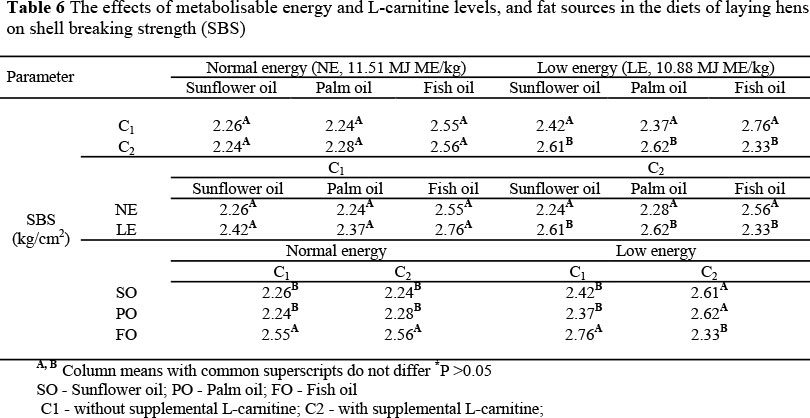
The interactions between metabolisable energy and L-carnitine levels and fat sources observed for SBS (P <0.001) are presented in Table 6. L-carnitine supplementation to LE diets containing SO or PO significantly increased SBS compared to a NE diet without L-carnitine. However, L-carnitine in the LE diet containing FO significantly decreased SBS compared to the diet without L-carnitine.
The inclusion of FO to the NE diet with or without L-carnitine significantly increased SBS compared to the diets containing SO and PO. Although FO inclusion to the LE diet without L-carnitine enhanced SBS (P <0.001), supplemental L-carnitine in the same diet decreased SBS (P <0.001) compared to diets containing SO or PO.
Peterson et al. (1960) and Clunies et al. (1992) recorded that dietary calcium levels required to maintain optimum eggshell quality are between 2.7 and 3.5% or 3.5 or 4.5%, respectively. Dietary Ca levels in the present study ranged from 3.55 to 3.64%. Increasing the calcium content of the diets seems to be detrimental not only to fat and energy utilization but also to calcium retention (Atteh & Leeson, 1984). This is mainly due to the problem of soap formation, which tends to increase with an increase in dietary calcium level, especially in the presence of saturated fatty acids (Atteh & Leeson, 1984). Increased calcium demands during the laying cycle in hens are associated with a proportional increase in intestinal calcium absorption (Bar et al., 1978) and a decrease in renal calcium excretion (DeLuca et al., 1990). Low dietary ratios of n-6 : n-3 fatty acids might increase intestinal calcium absorption, and lower bone Ca turnover (Classen et al., 1995). Fish oil reduced urinary calcium excretion significantly. Particularly, eicosapentaenoic acid (C20 : 5n-3) is effective in inhibiting bone resorption (Buck et al., 1991). Increasing renal calcium excretion due to the ß-oxidation of LCFAs in metabolism by supplemental L-carnitine might reduce SBS. Increase of SBS in hens fed a diet without L-carnitine might be a result of a reduced urinary calcium excretion.
Interactions between L-carnitine levels and fat sources for ESG, HU and AI are presented in Table 7. The use of PO in diets without L-carnitine significantly decreased the ESG compared to the diets containing FO or SO. However, the ESG of hens fed the diet containing FO with L-carnitine was lower than that of hens fed the diets containing SO or PO, or the diet without L-carnitine (P <0.001).
L-carnitine inclusion in the diet containing PO significantly increased HU and AI compared to the diet without L-carnitine. However, L-carnitine supplementation to the diet containing SO significantly decreased HU and AI compared to the diet without L-carnitine. HU and AI were significantly reduced by L-carnitine supplementation of the diets containing FO or SO compared to the diet with PO. Albumen quality usually depends on the amount of ovamicin, and ovamicin is synthesized from magnum (Anonymous, 2000b). High concentrations of dietary carnitine stimulate palmitate oxidation (Chiu et al., 1999). The inclusion of L-carnitine in the diet containing PO rich in palmitic acid might have increased ovamicin synthesis by enhancing energy production due to its stimulating effect on palmitate oxidation.
There was a significant interaction between metabolisable energy levels and fat sources in terms of HU and AI (Table 8).
Fish oil inclusion in the NE diet significantly decreased HU and AI of eggs compared to the diets containing PO or SO, or in the LE diet. The inclusion of PO to the NE diet significantly increased the HU and AI of the eggs compared to the LE diets. The fatty acid digestibility was increased by increasing unsaturation. The high digestibility of the fish lipids rich in PUFAs indicates a high energy value and positively affected HU and AI (Anonymous, 1985). The YI was significantly influenced by the dietary energy level and fat source (Table 5). The inclusion of SO or FO in the diet enhanced (P <0.001) the YI compared to the diet containing PO. Feeding LE diets significantly increased YI compared to the NE diet. The increase in YI might be due to an improvement in the hepatic biosynthesis rate of yolk precursors and/or an alteration in the mode of their transport from the liver into the ovarian follicle and oocyte (Rabie et al., 1997). In this connection, it has been reported that 95% of the total lipids of yolk is derived from triacylglycerol-rich lipoprotein which is synthesized in the liver and transferred into rapidly-developing yolk from the plasma over a period of several days before ovulation. The remaining yolk lipid is derived from the lipovitellin component of plasma vitollegenin (Griffin et al., 1984).
A significant interaction between metabolisable energy and L-carnitine levels in terms of yolk pH is presented in Table 9. L-carnitine supplementation to the NE diet significantly reduced yolk pH compared to those without L-carnitine. In contrast to these findings, egg yolk pH was increased significantly when L-carnitine was included in the LE diet compared with the LE diet without L-carnitine. In the NE diet without L-carnitine egg yolk pH increased (P <0.01), compared to that in the LE diet.
Albumen pH was significantly decreased by supplemental L-carnitine compared to the diet without L-carnitine (Table 5). A significant interaction between metabolisable energy levels and fat sources in terms of yolk pH and albumen pH was demonstrated (Table 10). The inclusion of FO to the LE diet resulted in a significantly higher increase in yolk pH than when the hens were fed the diets containing SO or PO. However, the yolk pH was significantly reduced by the use of SO or PO in the LE diet compared to the NE diet. Albumen pH was significantly increased with FO inclusion to the LE diet compared to a diet containing SO or PO, or the NE diet. The NE diet containing SO caused a significantly higher albumen pH compared to the LE diet containing SO.
Dietary treatments did not significantly affect EST (Table 5). In fact, reports on the effects of dietary L-carnitine on the egg quality of laying hens are limited. No comparable data could be found in the literature in connection with the effects of dietary L-carnitine level, energy level and fat source on egg qualities.
The results in the present study are consistent with those of Leibetseder (1995), Rabie et al. (1997) and Celik et al. (2004). Leibetseder (1995) pointed out that EP, FI or BW during the early laying period were not significantly affected by the supplementation of a laying hen's diet with either 500 mg L-carnitine or nicotinic acid, or a combination of the two substances. Celik et al. (2004) reported that L-carnitine had no effect on any performance parameters or internal egg quality with the exception of albumen weight and height. Rabie et al. (1997) observed that L-carnitine (500 mg/kg diet) did not influence EP, external egg quality and YI. They concluded that L-carnitine has a beneficial effect on albumen quality and could modify the components of the edible part of the egg during the late laying period.
Conclusions
Dietary supplemental L-carnitine or dietary ME levels or fat sources and/or interaction among dietary treatments significantly affected FI and FCR and egg quality parameters, except EST. Supplemental L-carnitine in a diet containing SO or PO and with LE may improve broken egg problems of egg producers' by increasing SBS. The inclusion of FO to the LE diet with L-carnitine is not recommended because FO decreases SBS. In conclusion, results obtained from the present study may be put into practice with the versatile preferences due to the positive effects of different energy levels or L-carnitine levels or fat sources used in laying hen diets on performance and egg quality.
References
Anonymous, 1985. International Association of Fish Meal Manufacturers. Technical Bulletin. No: 22, October. [ Links ]
Anonymous, 2000a. Isabrown Technical Handbook. p. 40. [ Links ]
Anonymous, 2000b. Egg-Grading Manuel. U.S. Dept. Agri. Agricultural Marketing Servise, Agricultural Handbook, Number 75. [ Links ]
AOAC, 2007. Official Methods of Analysis (18th ed.). Association of Official Analytical Chemists. Washington. D.C., USA. [ Links ]
Atteh, J.O. & Leeson, S., 1983. Influence of increasing dietary calcium and magnesium levels on performance, mineral metabolism, and egg mineral content of laying hens. Poult. Sci. 62, 1261-1268. [ Links ]
Atteh, J.O. & Leeson, S., 1984. Effects of dietary saturated or unsaturated fatty acids and calcium levels on performance and mineral metabolism of broiler chicks. Poult. Sci. 63, 2252-2260. [ Links ]
Bar, A., Cohen, A., Edelstein, S., Shemesh, M., Montecuccoli, G. & Hurwitz S., 1978. Involvement of cholecalciferol metabolism in birds in the adaptation of calcium absorptions to the needs during reproduction. Comp. Biochem. Physiol. 59B, 245-249. [ Links ]
Barker, D.L. & Sell, J.L., 1994. Dietary carnitine did not influence performance and carcass composition of broiler chickens and young turkeys fed low- or high-fat diets. Poult. Sci. 73, 281-287. [ Links ]
Bieber, L.L., 1988. Carnitine. Annu. Rev. Biochem. 57, 261-283. [ Links ]
Bish, C.L., Beane, W.L., Ruszler, P.L. & Cherry, J.A., 1985. Body weight influence on egg production. Poult. Sci. 64, 2259-2262. [ Links ]
Bremer, J., 1983. Carnitine metabolism and functions. Physiol. Rev. 63, 1420-1480. [ Links ]
Buck, A.C., Davies, R.L. & Harrison, T., 1991. The protective role of eicosapentaenoic acid (EPA) in the pathogenesis of nephrolithiasis. J. Urol. 146, 188. [ Links ]
Card, L.E. & Nesheim, M.C., 1972. Poultry Production. 11th ed. Lea and Febiger, Philadelphia, USA. [ Links ]
Celik, L.B., Tekeli, A. & Ozturkcan, O., 2004. Effects of supplemental L-carnitine in drinking water on performance and egg quality of laying hens exposed to a high ambient temperature. J. Anim. Physiol. 88, 229-233. [ Links ]
Chiu, K.M., Keller, E.T., Crenshaw, T.D. & Gravenstein, S., 1999. Carnitine and dehydroepiandrosterone sulfate induce protein synthesis in porcine primary osteoblast-like cells. Calcif. Tissue. Int. 64, 527-533. [ Links ]
Classen, N., Potgieter, H.C. & Seppa, M., 1995. Supplemented gamma-linolenic and eicosapentaenoic acid influence bone status in young male rats: effects on free urinary collagen crosslinks, total urinary hydroxyproline and bone calcium content. Bone 16, 385. [ Links ]
Clunies, M., Parks, D. & Leeson, S., 1992. Calcium and phosphorus metabolism and eggshell formation of hens fed different amounts of calcium. Poult. Sci. 71, 482-489. [ Links ]
DeLuca, H.F., Krisinger, J. & Darwick, H., 1990. The vitamin D system. Kidney Int. 38 (Suppl. 29), 2-8. [ Links ]
Duncan, D.B., 1955. Multiple range test and multiple F tests. Biometrics 11, 1-42. [ Links ]
Hamilton, R.M.G., 1982. Methods and factors that affect the measurement of egg shell quality. Poult. Sci. 61, 2022-2039. [ Links ]
Harmeyer, J., Schlumbohm, C. & Baumgartner, M., 1998. Der gehalt an L-carnitin in futter- und lebensmitteln pflanzlicher und tierischer herkunft und der einfluss von herstellungsverfahren auf dessen gehalt (L-carnitine content in feed and food of vegetable and animal origin and the impact of processing on the content). In: Einfluss von erzeugung und verarbeitung auf die qualitat landwirtschaftlicher produkte, Ed VDLUFA Verlag, Darmstad, Germany. pp. 489-492. [ Links ]
Hempe, J.M., Lauxen, R.C. & Savage, J.E., 1988. Rapid determination of egg weight and specific gravity using a computerised data collection system. Poult. Sci. 67, 902-907. [ Links ]
Hu, F.B., Bronner, L., Willett, W.C., Stampfer, M.J., Rexrode, K.M., Albert, C.M., Hunter, D. & Manson, J.E., 2002. Fish and omega-3 fatty acid intake and risk of coronary heart disease in women. JAMA 2002 Apr 10; 287 (14), 1518-1821. [ Links ]
Leibetseder, J., 1995. Studies of L-carnitine effects in poultry. Arch. Anim. Nutr. 48, 97-108. [ Links ]
Mast, J., Buyse, J. & Goddeeris, B.M., 2000. Dietary L-carnitine supplementation increases antigen-specific immunoglobulin G production in broiler chickens. Br. J. Nutr. 83, 161-166. [ Links ]
Mu, H. & Hoy, C.E., 2004. The digestion of dietary triacylglycerols. Prog. Lipid Res. 43,105-133. [ Links ]
Neuman, S.L., Lin, T.L. & Hester, P.Y., 2002. The effect of dietary carnitine on semen traits of white leghorn roosters. Poult. Sci. 81, 495-503. [ Links ]
Peterson, C.F., Conrad, D.H., Lumijarui, D.H., Sauter, E.A. & Lampman, C.E., 1960. Studies on the calcium requirements of high producing White Leghorn hens. Idaho Agric. Exp. Stn. Res. Bull. No. 44, 1-35. [ Links ]
Rabie M.H., Szilagyi M. & Gippert T., 1997. Effects of dietary L-carnitine on the performance and egg quality of laying hens from 65-73 weeks of age. Br. J. Nutr. 78, 615-623. [ Links ]
Rauch, W., 1965. Die elastische verformung von hühnereiern als mabstab für die beurteilung der schalenstabitaet (The elastic forming of hens' egg as a criteria for the evaluation of shell stability) Arch. Geflugelk. 29, 467-477. [ Links ]
Renner, R. & Hill, F.W., 1961. Factors affecting the absorbability of saturated fatty acids in the chick. J. Nutr. 74, 254-258. [ Links ]
Smink, W., Gerrits, W.J.J., Hovenier, R., Geelen, M.J.H., Lobee, H.W.J., Verstegen, M.W.A. & Beynen, A.C., 2008. Fatty acid digestion and deposition in broiler chickens fed diets containing either native or randomized palm oil. Poult. Sci. 87, 506-513. [ Links ]
SPSSWIN, 1994. SPSS for Windows 6.1.3. SPSSWIN, Istanbul, Turkey. [ Links ]
Vila, B. & Esteve-Garcia, E., 1996. Studies on acid oils and fatty acids for chickens. I. Influence of age, rate of inclusion and degree of saturation on fat digestibility and metabolisable energy of acid oils. Br. Poult. Sci. 37, 105-117. [ Links ]
WPSA, 1989. European Table of Energy Values for Poultry Feedstuffs (3rd ed.). WPSA Subcommittee, Beekbergen. The Netherlands. [ Links ]
Xu, Z.R., Wang, M.Q., Mao, H.X., Zhan, X.A. & Hu, C.H., 2003. Effects of L-carnitine on growth performance, carcass composition and metabolism of lipids in male broilers. Poult. Sci. 82, 408-413. [ Links ]
Young, R.J. & Garret, R.L., 1963. Effect of oleic and linoleic acids on the absorption of saturated fatty acids in the chick. J. Nutr. 81, 321-329. [ Links ]
 Correspondence:
Correspondence:
E-mail: corduk@agri.ankara.edu.tr


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}