










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 no.3 Pretoria mar. 2008
Effects of ovariectomy and anabolic steroid implantation on the somatotrophic axis in feedlot heifers
C.R. BaileyI, II; G.C. DuffI; S.R. SandersI; S.P. CuneoI; C.P. McMurphyI; S.W. LimesandI; J.A. MarchelloI; D.W. SchaferI; M.L. RhoadsI; D.M. HallfordIII
IDepartment of Animal Sciences, The University of Arizona, P.O. Box 210038, Tucson, Arizona 85721-0038, U. S. A
IIDepartment of Agriculture and Natural Resources, University of Tennessee-Martin, Martin, TN 38237, USA
IIIDepartment of Animal and Range Sciences, New Mexico State University, Las Cruces, 88003, USA
ABSTRACT
A 2 x 2 factorial experimental design was used to evaluate effects of ovariectomy and implantation (200 mg trenbolone acetate and 28 mg estradiol benzoate; Synovex-Plus) on the performance, serum urea nitrogen, serum insulin like growth factor -1 (IGF-1), and mRNA expression of hepatic IGF-1, growth hormone (GH) receptor and estrogen receptor-a as well as pituitary GH, estrogen receptor-a and GH releasing hormone receptor in feedlot heifers. Thirty-two British x Continental heifers were randomly assigned to either an ovariectomized (OVX) or intact group and then to either a control group or a Synovex-Plus implant group. Heifers were fed a 900 g/kg concentrate steam-flaked maize based diet for 42 d. Liver biopsies were taken before OVX for baseline expression of IGF-1, GH receptor and hepatic estrogen receptor-a, which did not differ between treatment groups. Blood and body weight (BW) were taken on d 0, 28, and 42 and one heifer/pen was slaughtered on d 42 for liver and pituitary tissue samples. Initial and final BW did not differ due to OVX or implant. No gender x treatment interaction was observed for average daily gain (ADG) or final BW. Neither OVX nor implant affected ADG for the final 14 d of the feeding period, but 28 and 42 d ADG were greater in implanted than in control heifers. No 2- or 3-way interactions with day were observed for serum urea nitrogen. Serum urea nitrogen was greater in control heifers than in implanted heifers, but gender had no effect. Both implant and OVX increased serum IGF-1 and a gender x treatment interaction was observed. Neither gender nor implant treatment affected hepatic estrogen receptor-a, hepatic IGF-1, hepatic GH receptor, GH releasing hormone receptor, GH, or pituitary estrogen receptor-a. Data indicate that implanting heifers with Synovex-plus increases serum IGF-1, but the reason for this increase cannot be explained by mRNA expression of key somatotropic genes in the present study.
Keywords: Synovex-plus, mRNA, gender
Introduction
United States law stipulates that all heifers entering from Mexico must either be ovariectomized (OVX) or have originated in a tuberculosis-free herd and test negative for brucellosis. Ovariectomization is a female castration procedure that not only renders the animal infertile, but also ceases the oestrous cycle and the behaviour associated with oestrus; thereby changing the gender of the animal, as gender can refer to male, female or neuter. Research concerning the effects of ovariectomy is of particular importance in the southwest United States. Early research showed that ovariectomy decreased average daily gain (ADG) and gain : feed (G:F) (Ray et al., 1969), but more recent research has indicated that ovariectomy does not reduce ADG (Klindt & Crouse, 1990). Specifically, when implanted with either 20 mg estradiol benzoate and 200 mg testosterone propionate (Synovex-H) or 20 mg estradiol benzoate and 200 mg progesterone (Synovex-S), ovariectomized (OVX) heifers have exhibited a four-fold greater response in ADG to implantation than intact heifers (Garber et al., 1990); and when implanted with Synovex-H, OVX heifers had similar ADG to intact heifers implanted with Synovex-H (Adams et al., 1990). However, the physiological and molecular explanation for this performance recovery, via implantation is poorly understood.
Vestergaard et al. (1995) found that ovariectomy did not affect circulating insulin like growth factor -1 (IGF-1) in unimplanted Holstein heifers, and Mader & Kreikemeier (2006) found IGF-1 concentrations to be greater in estradiol + trenbolone acetate (TBA) implanted heifers compared with unimplanted heifers, but found no differences in IGF-1 concentrations between controls and those animals implanted only with estradiol or TBA. However, TBA alone or implanted synergistically with an oestrogenic implant had no effect on growth hormone (GH; Hunt et al., 1991). Implanting lambs with estradiol + trenbolone acetate increased hepatic mRNA expression of IGF-1 and implanting steers with estradiol+ TBA increased expression of IGF-1 in the muscle (Johnson et al., 1998). These data suggest that increases in serum IGF-1 may be the result of increased hepatic and muscular IGF-1 mRNA expression. However, the mechanism behind the increased IGF-1 expression in implanted animals is still poorly understood.
Therefore, the hypothesis of this study was that the mechanism controlling increased IGF-1 expression might be associated with increased expression of GH or oestrogen receptors in the hepatocyte. Thus, the purpose of this study was to utilize quantitative reverse transcriptase polymerase chain reaction (RT-PCR) technology to examine the physiological mechanisms by which anabolic implants improve the growth performance of intact and OVX heifers in the feedlot.
Materials and Methods
All procedures were approved by the University of Arizona Institutional Animal Care and Use Committee.
A 2 x 2 factorial treatment arrangement was used to evaluate effects of OVX and implantation with Synovex-Plus [200 mg of trenbolone acetate and 28 mg of estradiol benzoate (EB); Fort Dodge Animal Health, Overland Park, KS] on ADG, serum urea nitrogen (SUN), serum IGF-1 and mRNA expression of hepatic IGF-1, total GH receptor and estrogen receptor-a as well as GH, estrogen receptor-a, and GH releasing hormone receptor in the pituitary.
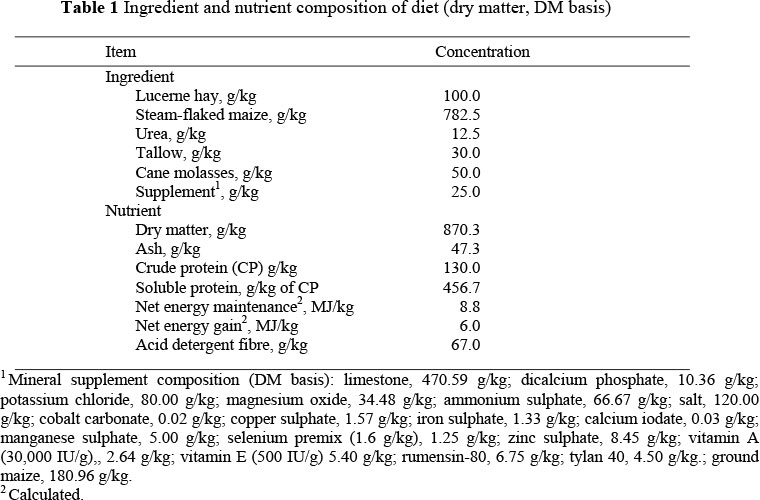
Thirty-two, two year old British x Continental heifers with an initial, unshrunk body weight (BW) of 374 ± 3.8 kg (mean ± s.e.) were stratified by BW and assigned to one of two gender groups (OVX or intact). Heifers entering this study were heavier than heifers normally entering the feedlot. However, this is typical for cull heifers of Mexican origin entering feedlots in the southwest United States. Heifers were then blocked by gender (n = 8 pens/gender) and randomly assigned to treatment [implanted (implant; n = 8) or non-implanted (control; n = 8)] and then to 1 of 16, partially shaded, soil surfaced pens (6.1 x 21.3 m; 2 heifers/pen), with concrete feed bunks and a shared water source for a 42-d feeding period. On d 0 cattle assigned to the implanted group were implanted with Synovex-Plus. Before implanting, the right ear was cleaned with an anti-microbial disinfectant, sprayed with 70% ethanol and wiped dry to prevent infection. Additionally, before use and between each animal, the implant gun was cleaned and sterilized using a sponge immersed in antimicrobial disinfectant and then wiped dry. Following implantation the ear was thoroughly inspected to ensure that all pellets were inserted and securely in place. Heifers were fed a 900 g/kg concentrate, steam-flaked maize based diet (Table 1), which was mixed daily prior to feeding (07:00). Feed samples were collected at 2-wk intervals and analyzed for dry matter (DM), crude protein (CP), soluble protein (g/kg of CP), acid detergent fibre (ADF), ash, NEg and NEm (Dairy One, Forage Testing Laboratory, Dairy One, Inc., Ithaca, NY 14850). Ort samples were taken at d 28, 42 and when inclement weather so required. Feed refusals were weighed and a representative sample from each pen was taken and mixed with samples from all other pens to form a composite sample that was then sub-sampled and analyzed for DM.
All surgical procedures were performed by, or under the direct supervision of a licensed veterinarian.
Liver biopsies were taken on d -14 between the 11th and 12th ribs on the perceptual line from the tuber coxae to the point of the scapula. The area was shaved, cleaned, and anesthetized by local infiltration of 5 to 10 mL of 2% lidocaine (lidocaine hydrochloride, injectable, 2%; Vedco Inc., St. Joseph, MO). A puncture incision was made through the skin with a scalpel blade and a biopsy needle was subsequently inserted through the underlying muscle layers and into the liver. Two biopsies (~100 mg/each) were taken and combined on the anterior side of a sterile collection glove. The samples were split equally into two screw top tubes, and subsequently snap frozen in liquid N. Upon completion of all biopsies, samples were transported to the University of Arizona Ruminant Nutrition Laboratory and stored at -80 °C for later mRNA expression analysis.
Surgical procedures for ovariectomy of animals assigned to the OVX group were performed on d -7 and -5. Heifers were restrained in a hydraulic working chute and bi-lateral ovariectomies were performed via entrance through the left para lumbar fossa. Before surgery the area was shaved, cleaned, and anesthetized by local infiltration of 25 to 30 mL of 2% lidocaine (lidocaine hydrochloride, injectable, 2%; Vedco Inc., St. Joseph, MO). A 10 to 15 cm vertical incision was made through the skin. The underlying muscle layers were separated via blunt dissection and the abdominal cavity was entered. Each ovary was located and removed from the broad ligament by hand and allowed to drop into the abdominal cavity. The skin incision was then closed by using a horizontal mattress suture pattern of #1 Vetafil. Heifers were subsequently maintained in individual pens on the previously mentioned diet until initiation of the study.
Blood samples were taken via jugular puncture on d 0, 28, and 42 and allowed to clot at room temperature before serum was harvested via centrifugation at 1,000 x g for 20 min and stored (-20 °C) for later analysis of SUN and serum IGF-1.
On d 42, following a 12-h fast, one heifer/pen was randomly selected and slaughtered via captive bolt and exsanguination, beginning at 07:00, at the University of Arizona Meats Laboratory, to obtain liver and pituitary samples. Pituitary glands were excised, trimmed and divided midsagitally and each half was placed in a separate screw top tube and snap frozen in liquid N within 30 min after exsanguination. Liver samples (5 to 10 g) were taken immediately following USDA inspection and further processed into two sub-samples (1 to 2 g), divided equally into two screw-top tubes and snap frozen in liquid N within 30 min after exsanguination. All pituitaries and liver samples were kept frozen in liquid N until they were transported to the University of Arizona Ruminant Nutrition Laboratory and stored at -80 °C for later mRNA expression analysis.
Serum urea N was determined using a direct colorimetric determination method (TECO Diagnostics, Anaheim, CA 92807) and serum IGF-1 (ng/mL) was determined by RIA in single assay (CV = 12%), as described by Berrie et al. (1995).
Pituitary (0.2 to 0.3 g; as-is basis), liver biopsy (0.1 g; as-is basis) and post-mortem liver (0.1 g as-is basis) samples were homogenized (Polytron, Brinkmann Instruments, Inc., Westbury NY) and total RNA was extracted with TRIzol (Invitrogen; Carlsbad, CA), according to manufacturers guidelines. Pelleted RNA was then re-suspended in molecular biology grade H2O and stored at -80 °C. Total RNA was DNase treated (DNase I - Amplification grade, Invitrogen). Final RNA sample concentration and quality were determined by measuring the absorbance at 260 nm and 280 nm with a Nanodrop ND-1000 Spectrophotometer. Aliquots of DNase-treated RNA, taken from pituitary and liver, were normalized to a concentration of 1 µg/(jL or 0.40 µg/µL, respectively, and cDNA was created using Super Script III (Super Script III First-Strand Synthesis System for RT-PCR; Invitrogen), according to product guidelines. Synthetic oligonucleotide primers (Table 2) were generated from published primer sequences for growth hormone (GH; Chen et al., 1997) and ERA (Lamote et al., 2006). Primers were designed with Primer Express 2.0 software for the GH releasing hormone receptor nucleotide sequence in GenBank (Accession # AF1848960) and un-published nucleotide sequences for IGF-1, hypoxanthine-guanine phosphoribosyltransferase and GH receptor (M. Rhodes, The University of Arizona, Tucson, AZ, personal communication).
Quantitative real time PCR was performed at the University of Arizona Genomic Analysis and Technology Core facilities on an ABI PRISM 7300 Sequence Detection System (Applied Biosystems; Foster City, CA). ABI PRISM 7300 Sequence Detection System Software (Applied Biosystems) was used to visualize and interpret data. Real time PCR conditions for amplification of all gene products of interest were 1 cycle at 50 °C (2 min), 1 cycle at 95 °C (3 min), followed by 40 cycles of 95 °C (15 s) and 66 °C (60 s), followed by 1 cycle at 95 °C (15 s) and 60 °C (30 s). Disassociation curves were run on every plate to ensure single product amplification per primer set. A reaction volume of 25 uL was identical for each sample well on every plate and all samples were run with iTaq SYBR Green Supermix with ROX (Bio-Rad Laboratories, Hercules, CA), in triplicate, alongside triplicate standard curves, in 96 well optical reaction plates with an optical reaction cover (Applied Biosystems).
Following each run, primer efficiencies were calculated using the equation: Efficiency = 1 - 10(-1/slopeof thestandardcurve). Average of all triplicates, across all genes, resulted in efficiencies of 98, 94, and 97% for liver biopsy, post mortem liver and pituitary plates, respectively. Standard deviation and CV values were calculated for cycle threshold values and all values had a CV of lower than 1.0%. Cycle threshold values for hypoxanthine-guanine phosphoribosyltransferase were statistically analyzed to ensure no treatment or gender differences existed, thereby quantifying the proper choice for an endogenous reference gene. The mathematical model used to quantify the relative expression or the change in the cycle threshold value (ACT) of each target gene is described by Pffafl (2001).
Due to insufficient RNA yield, three animals were excluded from liver biopsy gene expression analysis, two animals were excluded from post mortem liver gene expression analysis, and one animal was excluded from pituitary gene expression analysis. Therefore, these data were analyzed as missing values and statistical analysis was performed using the PROC MIXED procedures of SAS (2004). The model for all data included gender (intact vs. OVX), treatment (implant vs. control), pen (experimental unit) and the gender x implant treatment interaction as fixed effects and the random effect was heifer. The model for SUN and serum IGF-1 also included day as a repeated measure and tested all 2- and 3-way interactions with day and the gender x implant treatment x day interaction was considered the residual. Additionally, d 0 SUN was analyzed as a covariate, within sampling date, due to initial differences observed between gender groups; autoregressive of order 1 was used for a covariance structure. Significance was considered P <0.05 and LS means along with the most conservative estimate for s.e.m. were reported for each variable.
Results and Discussion
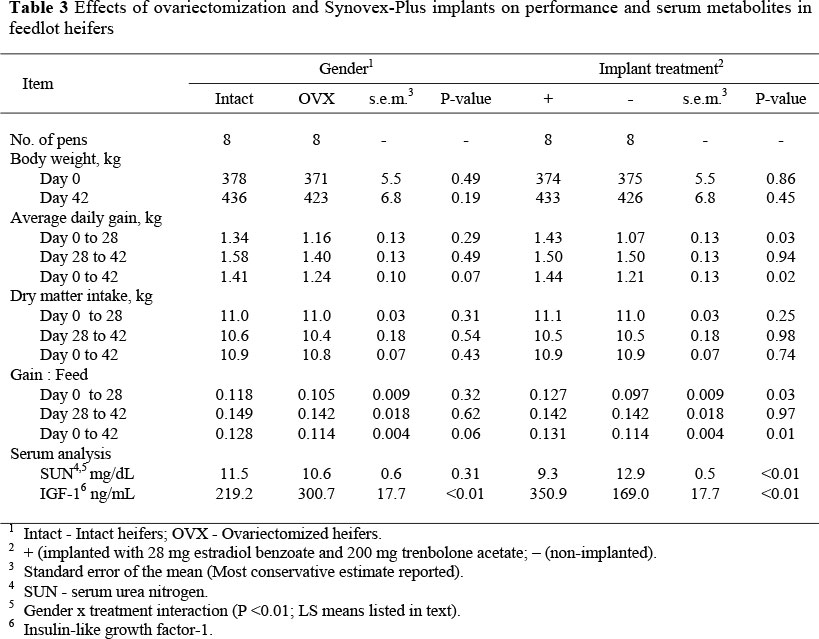
No gender x treatment interaction was observed for any performance parameters. No differences in d 0 BW were detected for either gender or treatment group, and d 42 BW was not changed as a result of gender or treatment (Table 3). Likewise, ADG (Table 3) for the final 14 d of the feeding period was not affected by gender or implant treatment. However, 28 d (P = 0.03) and overall feeding period ADG were greater (P = 0.02) in implanted than control heifers. These data agree with previous research conducted in our laboratory, which indicated that long-fed Holstein steers implanted initially (d 0) with 36 mg of zeranol followed by two subsequent implants of estradiol + trenbolone acetate on d 84 and 168 had greater ADG than non-implanted controls, when individually fed (Cheatham, 2005). Perry et al. (1991) reported that both Holstein and beef steers implanted with Revalor (28 mg estradiol + 140 mg trenbolone acetate) had greater ADG than non-implanted controls. Furthermore, Herschler et al. (1995) showed that implanting estradiol + trenbolone acetate ratios equal to those used in the present study improved ADG in both steers and heifers.
Gender did not affect ADG (Table 3) for the initial 28 d or the final 14 d of the study. While these data support research showing that OVX does not reduce ADG (Hamernik et al., 1985; Klindt & Crouse, 1990; Field et al., 1996), it is in contrast to early research that indicated that OVX decreased ADG (Dinusson et al., 1950; Ray et al., 1969; Horstman et al., 1982). However, heifers studied by Horstman et al. (1982) were implanted with zeranol only, a much less aggressive implant than used in the present study. Additionally, OVX heifers in that study only exhibited reduced ADG for the first 24 d of the study and surgeries were performed between d 8 and 10, whereas, in the present study, OVX surgeries were performed on d -5 and -7 and heifers were allowed to recover before the initiation of the study. Furthermore, no gender x implant treatment interaction was found. Therefore, this lack of significance, in relation to gender, could have been due to the fact that implanted OVX heifers exhibited ADG similar to that of implanted intact heifers and was numerically greater than intact control heifers for the initial 28 d of the study (data not reported). This is in agreement with Garber et al. (1990), as they found that OVX heifers exhibited a fourfold greater response to implantation than intact heifers. Also, Adams et al. (1990) reported that implanting OVX heifers with Synovex-H resulted in BW gains similar to those in intact heifers implanted with Synovex-H.
Neither gender nor implant treatment affected dry matter intake (DMI) and gender did not affect G:F. The lack of a gender effect on DMI agrees with early research that indicated that OVX did not have any affect on DMI (Adams et al., 1990). On the other hand, reports on the effects of anabolic implants on DMI vary. Adams et al. (1990) reported that implanting heifers with Synovex-H increased DMI; but Apple (1989) reported no difference in DMI between implanted and control steers. The lack of an effect of Synovex-Plus on DMI, in the present study, could also be attributed to the short time that these heifers were on feed. The lack of a gender effect on G:F disagrees with early research that indicated that OVX had an adverse influence on efficiency of growth (Dinusson et al., 1950; Horstman et al., 1982), but in agreement with later research that indicated that OVX did not have any effect on efficiency of growth (Klindt & Crouse, 1990). Due to the lack of difference in DMI and the increased ADG in implanted heifers compared to control heifers, G:F was greater for implanted heifers at both d 28 (P = 0.03) and 42 (P = 0.01), but did not differ between d 28 and 42. This finding agrees with previous reports that indicate implantation improves feed efficiency (Adams et al., 1990; Garber et al., 1990).
Serum urea N concentrations (Table 3) were lower (P < 0.01) in intact than OVX heifers on d 0 (data not reported), thus baseline SUN was analyzed as a covariate, within sampling date, to alleviate any pre-treatment effects at d 28 and 42. This is not surprising as OVX operations were completed on d -7 and -5 and OVX heifers were likely still recovering from the surgical procedures and therefore N utilization would have been reduced. Following covariate analysis, no 2 or 3 way interactions with time were observed for SUN; thus only overall means are reported for implant treatment and gender. These data indicate a trend for a gender x implant treatment interaction for SUN (P = 0.09; s.e.m. = 0.87) with LS means of 10.41 (intact implanted), 12.64 (intact control), 8.10 (OVX implanted) and 13.10 mg/dL (OVX control). Additionally, SUN was greater in control heifers than in implanted heifers (P <0.01). Considering the greater ADG and G:F observed in implanted heifers, one would expect that they utilized N better than control heifers. Supporting the present study, previous research indicated that heifers implanted with estradiol+trenbolone acetate had reduced plasma urea N compared to non-implanted controls or heifers implanted with either estradiol or trenbolone acetate alone (Mader & Kreikemeier, 2006). On the other hand, Cheatham (2005) indicated that SUN did not differ between implanted and non-implanted Holstein steers. In that study, SUN concentrations were much lower (near 5 mg/dL) than in the present study (>8.0 mg/dL). Johnson & Preston (1995) reported that plasma urea N concentrations greater than 5.0 to 8.0 mg/dL were indicative of excessive N intake and N wasting. This would indicate that heifers in the present study were fed at or in excess of their CP requirement, whereas Holstein steers in the previous study were fed at or possibly somewhat below their optimal CP, which would explain the reason for treatment differences in the present study and not in the previous study. Finally, gender had no effect on SUN (Table 3). However, this could be expected considering the lack of change in ADG due to OVX, the trend for a gender x treatment interaction observed for SUN, and previous research conducted in our laboratory, indicated gender did not affect SUN when steers were compared to heifers (Bailey et al., 2006).
Implants increased (P <0.01) serum IGF-1 (Table 3), which was greater (P <0.01) in OVX than intact heifers. This, however, is explained by the gender x treatment interaction (P <0.01; s.e.m. = 25.5) observed for serum IGF-1 with LS means of 267.4 (intact implanted), 170.9 (intact control), 434.4 (OVX implanted) and 167.0 ng/mL (OVX control) indicating that the greatest response in serum IGF-1 was in the OVX implanted heifers. This agrees with many previous reports indicating that implanting steers increases circulating IGF-1 (Lee & Henricks, 1990; Hunt et al., 1991; Cheatham, 2005). Furthermore, Mader & Kreikemeier (2006) found IGF-1 concentrations to be greater in estradiol+trenbolone acetate implanted heifers compared to controls, but found no differences in IGF-1 between controls and those animals implanted only with estradiol or trenbolone acetate. Moreover, serum IGF-1 is generally considered to be a GH dependent peptide (Florini et al., 1991). However, implants do not increase circulating GH (Hunt et al., 1991; Hayden et al., 1992). In fact, Cheatham (2005) reported that, in non-implanted Holstein steers, the GH response to a GH releasing hormone challenge was actually greater than that found in implanted steers. Although circulating GH was not measured in our study, data reported for serum IGF-1 agrees with previous research, which may indicate that anabolic implants may directly stimulate IGF-1, as opposed to acting through GH and the somatotropic axis. Therefore, to evaluate effects of implanting feedlot heifers on the somatotropic axis, mRNA expression of genes associated with OVX, implants and the somatotropic axis were analyzed.
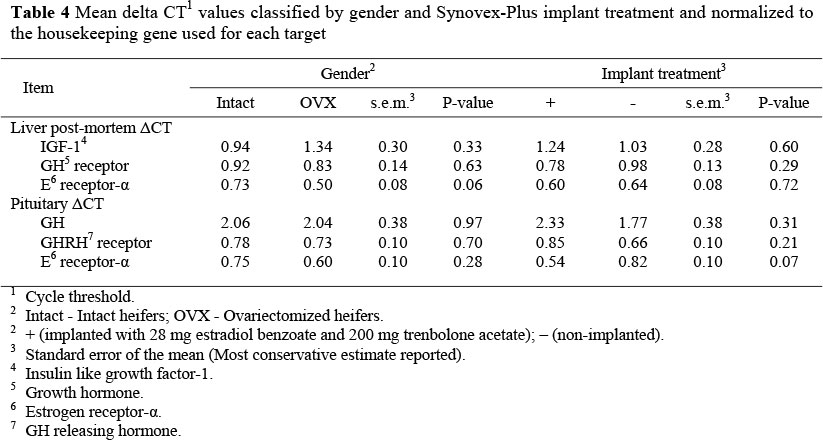
An initial liver biopsy was taken on d -14 to quantify expression of hepatic IGF-1, estrogen receptor-a, and GH receptor. Relative gene expression analysis revealed no pre-treatment differences (data not reported) for any of the genes examined.
No difference was observed in post mortem hepatic IGF-1 ACT values (Table 4) as a result of OVX or implant. Previous gene expression data has showed that implanting wethers with estradiol (8 mg) + trenbolone acetate (40 mg) increased steady-state hepatic IGF-1 expression by 150% and suggested that the liver may be the source of at least part of the increased circulating IGF-1 in steroid implanted animals (Johnson et al., 1998). Additionally, the same authors reported that implanting steers with Revalor-S (24 mg E2 + 120 mg trenbolone acetate) increased IGF-1 mRNA in the longissimus muscle by 68%. Furthermore, in vitro research indicates that treating bovine satellite cell cultures with either estradiol or trenbolone increases IGF-1 mRNA level (Kamanga-Sollo et al., 2004). However, the timeline for measurable effects of implantation with estradiol + TBA on gene expression is not consistent. Reports have varied from as early as 7 d post implantation (Pampusch et al., 2003) to as late as 28 d post-implantation (Dunn et al., 2003). It should be noted, however, that the Pampusch et al. (2003) study was a comparison within steer (i.e. d 0 to d 7) and the Dunn et al. (2003) study compared implanted to non-implanted steers, which could explain differences in the final expression between studies. Pampusch et al. (2003) observed that IGF-1 expression on d 26 was three times greater than on d 0. Dunn et al. (2003) showed that, although IGF-1 in circulation increased as early as 14 d post-implantation, mRNA expression of IGF-1 in the muscle did not increase until d 28 and was 2.4 fold greater in implanted calves than non-implanted calves at that time. This latter study demonstrates that an increase in serum IGF-1 may be observed without an increase in hepatic IGF-1 expression, as noted in the present study. In our study, a trend (P = 0.11) for a gender x treatment interaction was observed with ACT values of 0.70 (intact implanted), 1.18 (intact control), 1.80 (OVX implanted) and 0.89 (OVX control). This is similar to the response we observed for serum IGF-1 in that the difference in the relative hepatic IGF-1 expression between intact implanted and OVX implanted heifers (P = 0.07) could explain the reason for an increased response of OVX heifers to anabolic implants.
Hepatic estrogen receptor-a expression (Table 4) was not affected by treatment and no gender x treatment interaction was observed. In vitro research indicated that when bovine satellite cell cultures were treated with estradiol or TBA, both estrogen receptor-a and androgen receptor mRNA increased when added at 0.001 nM each. However, neither ER-a nor androgen receptor mRNA levels were affected by estradiol or TBA, compared to non-treated controls, when the steroids were added at greater concentrations (Kamanga-Sollo et al., 2004). Notably, a trend for intact heifers to have a greater hepatic estrogen receptor-a expression than OVX heifers (P = 0.06) was observed. A more in depth analysis revealed that this trend may be influenced by numerical differences in the relative expression of hepatic estrogen receptor-a between OVX implanted and intact implanted heifers with ACT values of 0.43 compared to 0.76 (s.e.m. = 0.16; P = 0.06), respectively. This could allow for estradiol supplied by the implant to anabolically act through the IGF-1 receptor, as was demonstrated in the mouse (Klotz et al., 2002). However, further research should be conducted analyzing IGF-1 receptor expression to potentially validate this hypothesis.
Pituitary estrogen receptor-a expression results are different, in that treatment (P = 0.07) and not gender, decreased the ACT value of pituitary estrogen receptor-a. Additionally, differences observed among interaction means showed that OVX implanted heifers had lower (P <0.05) pituitary estrogen receptor-a expression (ACT = 0.39) than either intact or OVX controls, which had ACT values of 0.82 and 0.81, respectively and they also tend to express lower (P = 0.13) pituitary estrogen receptor-a than intact implanted heifers (ACT = 0.69). These effects suggest that implantation with estradiol + TBA implants may also suppress pituitary estrogen receptor-a more in OVX heifers than in intact heifers which could also suggest an increase in binding to the IGF-1 receptor. However, further analysis is warranted to examine this response.
Neither gender nor treatment affected (P >0.65) the ACT values of GH releasing hormone receptor, GH, hepatic GH receptor, and there were no gender x treatment interactions observed for GH releasing hormone receptor, GH or hepatic GH receptor. Lack of differences in the relative ACT values in the expression of mRNA from these genes strengthens the hypothesis that estradiol + TBA implants are acting outside the somatotropic axis to increase IGF-1.
Conclusions
Data indicate that reductions in performance of OVX heifers can be eliminated by using an estradiol + TBA implant, which is likely due to an increase in serum IGF-1. However, the reason for the increase in serum IGF-1 is not clear. Trends in gene expression analysis suggest that the increased serum IGF-may be controlled outside of the somatotropic axis. Further research is warranted to examine the effects of OVX and anabolic implants on the somatotropic axis.
Acknowledgements
The authors would like to thank Elanco Animal Health, DSM Nutritional Products, Inc. and Fort Dodge Animal Health for product support; J. English, K. Cannon and M. Townsend for their assistance at the University of Arizona Feedlot.
References
Adams, T.E., Dunbar, J.R., Berry, S.L., Garrett, W.N., Famula, T.R. & Lee Y.B., 1990. Feedlot performance of beef heifers implanted with Synovex-H: effect of melengesterol: acetate, ovariectomy or active immunization, against GnRH. J. Anim. Sci. 68, 3079-3085. [ Links ]
Apple, J.K., 1989. Effects of Finaplix, Synovex-S, and Ralgro implants, singularly or in combinations on performance carcass traits and longissimus palatability of Holstein steers. M.S. thesis, Kansas State University, Manhattan, USA. [ Links ]
Berrie, R.A., Hallford, D.M. & Galyean, M.L., 1995. Effects of zinc source and level on performance and metabolic hormone concentrations of growing and finishing lambs. Prof. Anim. Sci. 11, 149-156. [ Links ]
Cheatham, R.C., 2005. Effects of anabolic implants on live performance, carcass characteristics, serum metabolites, somatotropin response to GRF, fatty acid profiles and lipogenic gene expression in adipose tissue in long-fed Holstein steers. M.S. thesis, University of Arizona, Tucson. [ Links ]
Chen, H.T., Schuler, L.A. & Schultz, R.D., 1997. Growth hormone and PIT-1 expression in bovine fetal lymphoid cells. Dom. Anim. Endocrinol. 14, 399-407. [ Links ]
Dinusson, W.E., Andrews, F.N. & Beeson, W.M., 1950. The effects of stilbestrol, testosterone, thyroid alteration, and spaying on the growth and fattening of beef heifers. J. Anim. Sci. 9, 321-330. [ Links ]
Dunn, J.D., Johnson, B.J., Kayser, J.P., Waylan, A.T., Sissom, E.K. & Drouilliard, J.S., 2003. Effects of flax supplementation and combined trenbolone acetate and estradiol implant on circulating insulin-like growth factor-1 and muscle insulin-like growth factor-1 messenger RNA levels in beef cattle. J. Anim. Sci. 81, 3028-3034. [ Links ]
Field, R., McCormick, R., Balasubramanian, V., Sanson, D., Wise, J., Hixson, D., Riley, M. & Russell, W., 1996. Growth, carcass, and tenderness characteristics of virgin, spayed, and single-calf heifers. J. Anim. Sci. 74, 2178-2186. [ Links ]
Florini, J.R., Ewton, D.A. & Magri, K.A., 1991. Hormones, growth factors, and myogenic differentiation. Ann. Rev. Physiol. 53, 201-216. [ Links ]
Garber, M.J., Roeder, R.A., Combs, J.J., Eldridge, L., Miller, J.C., Hinman, D.D. & Ney, J.J., 1990. Efficacy of vaginal spaying and anabolic implants on growth and carcass characteristics in beef heifers. J. Anim. Sci. 68, 1469-1475. [ Links ]
Hamernik, D.L., Males, J.R., Gaskins, C.T. & Reeves, J.J., 1985. Feedlot performance of hysterectomized and ovariectomized heifers. J. Anim. Sci. 60, 358-362. [ Links ]
Hayden, J.M., Bergen, W.G. & Merkel, R.A., 1992. Skeletal muscle protein metabolism and serum growth hormone, insulin and cortisol concentrations in growing steers implanted with estradiol-17β, trenbolone acetate, or estradiol-17ß plus trenbolone acetate. J. Anim. Sci. 70, 2109-2119. [ Links ]
Herschler, R.C., Olmsted, A.W., Edwards, A.J., Hale, R.L., Montgomery, T., Preston, R.L., Bartle, S.J. & Sheldon, J.J., 1995. Production responses to various doses and ratios of estradiol benzoate and trenbolone acetate implants in steers and heifers. J. Anim. Sci. 73, 2873-2881. [ Links ]
Horstman, L.A., Callahan, C.J., Morter, R.L. & Amstutz, J.E., 1982. Ovariectomy as a means of abortion and control of estrus in feedlot heifers. Theriogenology 17, 273-292. [ Links ]
Hunt, D.W., Henricks, D.M., Skelley, G.C. & Grimes, L.W., 1991. Use of trenbolone acetate and estradiol in intact and castrate male cattle: Effects on growth, serum hormones, and carcass characteristics. J. Anim. Sci. 69, 2452-2462. [ Links ]
Johnson, B.J., White, M.E., Hathway, M.R., Christians, C.J. & Dayton, W.R., 1998 Effect of combined trenbolone acetate and estradiol implant on steady-state IGF-1 mRNA concentrations in the liver of wethers and the longissimus muscle of steers. J. Anim. Sci. 76, 491-497. [ Links ]
Johnson, J.W. & Preston, R.L., 1995. Minimizing N waste by measuring plasma urea-N levels in steers fed different dietary crude protein levels. Texas Tech University Technical Report T-5-356, 62-63. [ Links ]
Kamanga-Sollo, E., Pampusch, M.S., XI, G., White, M.E., Hathaway, M.R. & Dayton, W.R., 2004. IGF-1 mRNA levels in bovine satellite cell cultures: Effects of fusion and anabolic steroid treatment. J. Cell. Physiol. 20, 181-189. [ Links ]
Klindt, J. & Crouse, J.D., 1990. Effect of ovariectomy and ovariectomy with ovarian auto transplantation on feedlot performance and carcass characteristics of heifers. J. Anim. Sci. 68, 3481-3487. [ Links ]
Klotz, D.M., Hewitt, S.C., Ciana, P., Raviscioni, M., Lindzey, J.K., Foley, J., Maggi, A., DiAugustine, R.P. & Korach, K.S., 2002. Requirement of E receptor-a in insulin-like growth factor-1 (IGF-1)-induced uterine responses and in vivo evidence for IGF-1/estrogen receptor cross-talk. J. Biol. Chem. 277, 8531-8537. [ Links ]
Lamote, I., Meyer, E., De Ketelaere, A., Duchateau, L. & Burvenich, C., 2006. Expression of the estrogen receptor in blood neutrophils of dairy cows during the periparturient period. Theriogenology 65, 1082-1098. [ Links ]
Lee, C.Y. & Henricks, D.M., 1990. Comparisons of various acidic treatments of bovine serum on insulin-like growth factor-1 immunoreactivity and binding activity. J. Endocrinol. 127, 139-148. [ Links ]
Mader, T.L. & Kreikemeier, W.M., 2006. Effects of growth promoting agents and season on blood metabolites and body temperature in heifers. J. Anim. Sci. 84:1030-1037. [ Links ]
Pampusch, M.S., Johnson, B.J., White, M.E., Hathaway, M.R., Dunn, J.D., Waylan, A.T. & Dayton, W.R., 2003. Time course of changes in growth factor mRNA levels in muscle of steroid-implanted and non-implanted steers. J. Anim. Sci. 81, 965-972. [ Links ]
Pfaff, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nuc. Acids Res. 29, 2002-2007. [ Links ]
Perry, T.C., Fox, D.G. & Beerman, D.H., 1991. Effect of an implant of trenbolone acetate and estradiol on growth, feed efficiency, and carcass composition of Holstein and beef steers. J. Anim. Sci. 69, 4696-4702. [ Links ]
Radcliff, R.P., McCormack, B.L., Crooker, B.A. & Lucy, M.C., 2003. Plasma hormones and expression of growth hormone receptor and insulin-like growth factor-I mRNA in hepatic tissue of periparturient dairy cows. J. Dairy Sci. 2003 86: 3920-3926. [ Links ]
Ray, D.E., Hale, W.H. & Marchello, J.A., 1969. Influence of season, sex and hormonal stimulants on feedlot performance of beef cattle. J. Anim. Sci. 29, 490-495. [ Links ]
SAS, 2004. Statistical Analysis Systems user's guide, (9th ed.) SAS Institute Inc., Cary, NC, USA. [ Links ]
Vestergaard, M., Purup, S., Henckel, P., Tonner, E., Flint, D.J., Jensen, L.R. & Sejrsen, K., 1995. Effects of growth hormone and ovariectomy on performance, serum hormones, insulin-like growth factor-binding proteins and muscle fiber properties of prepubertal Friesian heifers. J. Anim. Sci. 73, 3574-3584. [ Links ]
 Correspondence:
Correspondence:
E-mail: crbailey@utm.edu


{kind=link}
{kind=link}
{kind=link}
{kind=link}