










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.2 Pretoria Feb. 2008
SHORT COMMUNICATION
Mapping QTL for fatty acid composition that segregates between the Japanese Black and Limousin cattle breeds
N.O.M. TshipulisoI; L.J. AlexanderII; T.W. GearyII; W.M. SnellingIII; D.C. RuleIV; J.E. KoltesV; B.E. MoteV; M.D. MacNeilII
IGenetics Department, University of the Free State, Bloemfontein 9300; & Agricultural Research Council, Private Bag X2, Irene 0062, South Africa
IIUSDA Agricultural Research Service, Miles City, Montana 59301, USA
IIIUSDA Agricultural Research Service, Clay Centre, Nebraska 68933, USA
IVDepartment of Animal Science, University of Wyoming, Laramie 82071, USA
VDepartment of Animal Science, Iowa State University, Ames 50011, USA
ABSTRACT
The objective of this study was to search for quantitative trait loci (QTL) that segregate between Japanese Black and Limousin cattle breeds and affect relative amounts of saturated (SFA), mono-unsaturated (MUFA) and poly-unsaturated (PUFA) fatty acids. Six F1 Japanese Black-Limousin cross bulls were joined with 121 F1 females over a three-year period to produce 328 F2 progeny. Calves, aged from 450 to 641 d (average 561 d), were harvested using standard industry procedures. After aging, 2.54 cm thick steaks were cut from the posterior end of the wholesale rib, frozen at -20 °C, and held for determination of fatty acid composition. Percentages of the individual fatty acids were classified into percentages of SFA, MUFA and PUFA. Two hundred seventeen microsatellite markers covering the 29 bovine autosomes were assayed and QTL were identified by least squares regression. Genome-wise significant QTL with additive effects on SFA (-0.61 ± 0.19%), MUFA (0.93 ± 0.19%) and PUFA (-0.52 ± 0.11%) were observed near the centromere of BTA2. Also observed were five QTL indicative of dominance effects on: MUFA (BTA9, 119 cM, -1.87 ± 0.72%; BTA22, 47 cM, 1.85 ± 0.60%) and PUFA (BTA9, 54 cM, -1.49 ± 0.42%; BTA10, 38 cM, 1.20 ± 0.35%; and BTA15, 14 cM, 1.11 ± 0.36%). Based on these results, we conclude that it may be possible to improve the healthfulness of beef by manipulating fatty acid composition using genetic markers and appropriate crossbreeding systems.
Keywords: Beef, fatty acids, Japanese Black, Limousin, quantitative trait loci, Wagyu
Meat contains a mixture of saturated (SFA), mono-unsaturated (MUFA) and poly-unsaturated (PUFA) fatty acids. Fatty acid composition in beef has received considerable interest in view of its implications for human health and meat quality characteristics (Wood et al., 2004). While unsaturated fats are beneficial when consumed in moderation, high levels of saturated fat are associated with increased serum low-density lipoprotein cholesterol concentrations and pose a risk factor for coronary heart disease (Department of Health, 1994; Katan et al., 1994, 2000). In addition to the human health implications of fatty acid composition, beef with the most desirable flavour has a lower percentage of SFA and PUFA and a higher percentage of MUFA in the muscle fat (Melton et al., 1982).
Based on experimental comparisons of Limousin and Wagyu germplasm it was concluded that these breeds differ markedly with respect to fat deposition (Mir et al., 2002, Pitchford et al., 2002). Beef from the Wagyu breed, Japanese Black, is known for its extensive intramuscular fat or marbling. In contrast, beef from Limousin is lean with little marbling. Further, Japanese Black beef has a greater absolute level of MUFA and a greater ratio of MUFA relative to SFA than other breeds (e.g. Sturdivant et al., 1992; May et al., 1993; Mir et al., 2000). These differences provide an opportunity to identify quantitative trait loci (QTL) that affect carcass quality and fatty acid composition, to enhance meat palatability and minimize human health implications. Thus, the objective of this study was to search for QTL segregating in a population derived from a cross between Japanese Black and Limousin and affecting relative amounts of SFA, MUFA and PUFA. Further details of this project and QTL affecting carcass traits, meat quality, and absolute amounts of various fatty acids were presented previously by Alexander et al. (2007a; b).
Six F1 Japanese Black-Limousin cross bulls were joined with 121 F1 females over a three-year period to produce 328 F2 progeny. Calves were reared by their dams until weaning at approximately 175 d of age (SD = 14 d). Each year, before harvest and within sex, calves were randomly assigned to a harvest date in groups of 8 to 11 hd per d. After weaning, the calves were managed in a two-phase system: a growing phase with diet composition of 50 to 54% DM, 14.4 to 15.6% CP, and 4.44 to 4.94 MJ/kg of NEg, and a finishing phase with diet composition of 68 to 70% DM, 11.6 to 13.4% CP, and 5.27 to 5.48 MJ/kg of NEg. The finishing diet was fed for a minimum of 113 d until the calves were harvested. Within year and sex, groups of calves were harvested at two to three week intervals. Thus, the final group harvested each year had been consuming the finishing diet for at least 210 d. Calves, aged from 450 to 641 d (average 561 d), were transported to the abattoir the afternoon before harvest, held overnight with water and without feed, and harvested the next morning using standard industry procedures. To insure the ethical treatment of animals, all protocols involving the use of animals were approved by the Fort Keogh Livestock and Range Research Laboratory Animal Care and Use Committee. Two days post mortem, the wholesale rib was removed, vacuum packaged, and aged for 14 d at 2 °C. After aging, a three-rib section and four 2.54 cm thick steaks were cut from the posterior end of the wholesale rib, then individually vacuum packaged, frozen at -20 °C, and held for further analyses.
Two steaks were transported to University of Wyoming for determination of fatty acid composition as described by Rule et al. (2002). Briefly, the entire core of the Longissimus dorsi was sampled (i.e., devoid of trim fat and extraneous muscles) by dicing the muscle into 1.0 cm cubes while the muscle was semi-frozen, and weighed into pre-weighed, plastic cups with perforated lids. All samples were freeze-dried (Genesis 25 freeze dryer, The VirTis Co., Gardiner, NY), and then ground and homogenized using a home-style electric coffee grinder. Samples were packed into 20 mL plastic vials and sealed to inhibit exposure to air, and then stored at -80 °C until analyzed for fatty acids, which occurred within 2 to4 wks of freeze drying. Approximately 150 mg of dried muscle were weighed, in duplicate, into 16 mm x 125 mm screw-capped tubes that contained 1.0 mg of tridecanoic acid as internal standard, and then subjected to direct saponification as described by Rule et al. (2002). Samples were reacted with 4.0 mL of 1.18 M KOH in ethanol at 90 °C along with frequent vortex-mixing (two to three times per minute for 3 sec each time) until the sample was completely dissolved, except for insoluble collagen that appeared as a white powder in suspension upon mixing. After about 45 min, tubes were cooled, 2.0 mL water were added and non-saponified lipids extracted with 2.0 mL of hexane and removed. One millilitre of concentrated HCl was added to the original tubes and fatty acids extracted in 2.0 mL of hexane for fatty acid methyl ester (FAME) preparation, which was carried out according to Rule et al. (2002) using methanolic HCl as a catalyst. Analysis of conjugated linoleic acid (CLA) is hampered by use of acid catalysts because of partial geometric isomerization of cis-9, trans-11 CLA to trans-9, trans-11 CLA (Yamasake et al., 1999) and degradation of CLA to allylic methoxy artifacts (Kramer et al., 1997). However, Murrieta et al. (2003) demonstrated that dietary treatment effects on CLA in ovine muscle were maintained when acid catalysts were used for FAME preparation, despite up to 20% loss of cis-9, trans-11 CLA. Preparation of FAME from nonesterified free fatty acids (NEFA) requires the use of the acid catalyst because alkaline catalysts do not react with NEFA to form fatty acid methyl esters (Christie, 1982). For the current study, freeze dried muscle samples were chosen at random from approximately 5% of the samples for FAME preparation using methanolic KOH, which does not affect CLA proportions. We did not observe losses of CLA in the samples analyzed (data not shown). Generally, we have observed either minimal or no loss of CLA in samples containing low concentrations (about 0.5 mg per 100 mg of total fatty acids) of this fatty acid. Quantities of the various fatty acids were summarized by calculating relative amounts of SFA, MUFA and PUFA.
DNA was extracted from semen of sires of the F1 bulls and from white blood cells of the F1 bulls, F1 females, and F2 calves using standard protocols (e.g. Ausubel et al., 1994). Touchdown PCR was performed in MJ Research (Waltham, MA, USA) thermocyclers as described in http://biolibrary.licor.com/htdocs/RnP/LabPrim.jsp. All genotypes were collected on a LiCor 4200 DNA Analysis System (Lincoln, NE, USA). Genotypes were independently scored by two individuals. PCR was repeated for discrepancies that could not be resolved. Anomalous genotypes were detected using GenoProb (Thallman et al., 2001a,b). Genetic linkage maps were constructed using CRI-MAP (Green et al., 1990; http://compgen.rutgers.edu/multimap/crimap/). Two hundred seventeen markers covering the 29 bovine autosomes were chosen from http://www.marc.usda.gov/genome/cattle/cattle.html based on marker position, suitability for multiplex reactions, ease of scoring and number of alleles. Distance between markers was less in regions containing previously detected QTL for the amounts of various fatty acids.
Quantitative trait loci were identified by least squares regression using the F2 option of the QTL Express software for analysis of quantitative trait data from outbred populations (Seaton et al., 2002). An F-statistic profile was generated at 1-cM intervals for each chromosome. Effects of one additive QTL and one QTL with additive and dominance effects were modelled in separate analyses. The phenotypes were simultaneously adjusted for fixed effects of year and sex, and covariate age at harvest. Thus, given the random assignment of animals to harvest dates and the statistical model used to analyze the data, inference here is to an age-constant endpoint. Observed significance levels were adjusted to a genome-wide basis following Cheverud (2001). Ninety-five percent confidence intervals (CI) for the QTL map locations were calculated as described by Darvasi & Soller (1997). The hypothesis of two QTL versus one QTL was also tested for each phenotype - chromosome combination, again using QTL Express.
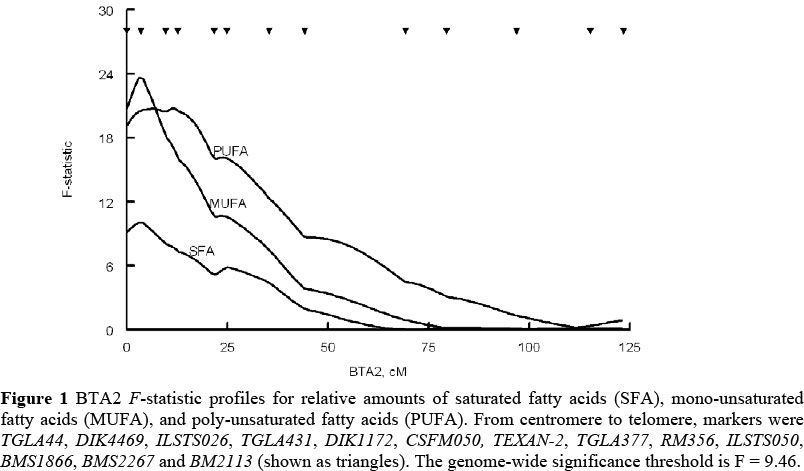
Genome-wise significant QTL with additive effects on SFA (-0.61 ± 0.19%, P = 0.04) at 4 cM (CI = 54 cM), MUFA (0.93 ± 0.19%, P <0.01) also at 4 cM (CI = 24 cM), and PUFA (-0.52 ± 0.11%, P < 0.01) at 11 cM (CI = 28 cM) were observed situated between markers DIK4469 and ILISTS026 on BTA2 (Figure 1). Substitution of an allele from Japanese Black for one from Limousin decreased percentages of SFA and PUFA and increased MUFA content. This QTL was also identified by Alexander et al., (2007b) as affecting fatty acid content, marbling, and flavour. Previous studies have positively associated desirable flavour with the MUFA oleate (Dryden & Marchello, 1970; Westerling & Hendrick, 1979; Melton et al, 1982).
Growth differentiation factor 8, or myostatin is co-located with the observed QTL on BTA2 and has profound effects on muscularity and beef composition (McPherron et al., 1997; Smith et al., 1997). Raes et al. (2001) suggested the polyunsaturated/saturated fatty acid ratio increased with low intramuscular fat content, likely because of the decreased saturated fatty acids associated with decreased intramuscular adipose tissue. Several mutations in myostatin cause exceptional muscle development or muscle hypertrophy which is commonly referred to as double muscling (McPherron & Lee, 1997). However, neither Limousin nor Japanese Black are thought to be double muscled and the classical phenotype was not observed in the F2 generation. Sequencing the myostatin gene revealed five of the F1 sires were heterozygous for the previously reported (Sellick et al., 2007) transversion (g.433C>A) resulting in an amino acid change of phenylalanine to leucine at position 94 of the myostatin protein.
Two QTL indicative of dominance effects were observed on BTA9. A QTL located at 54 cM (CI = 39 cM) affected PUFA (-1.49 ± 0.42%, P < 0.01). The second QTL affected MUFA (-1.87 ± 0.72%, P = 0.09) and was located at 119 cM (CI = 67 cM). As the total of SFA, MUFA, and PUFA is 100%, these results were potentially indicative of two QTL on BTA9. However, the two QTL model was only nominally better than the single QTL model for MUFA (P = 0.04). In each case, substitution of an allele from Japanese Black for one from Limousin indicated a decrease in the proportion of the respective group of fatty acids. Three additional QTL that were also suggestive of dominance effects on the relative amounts of fatty acids were also detected. QTL affecting PUFA content were observed at 38 cM (CI = 62 cM) on BTA10 (1.20 ± 0.35, P = 0.02) and at 14 cM (CI = 59 cM) on BTA15 (1.11 ± 0.36, P <0.01). Finally, a QTL affecting MUFA content was found at 47 cM (CI = 59 cM) on BTA22 (1.85 ± 0.60, P = 0.02). To the best of our knowledge, this is the first report of QTL with dominance effects on the fatty acid composition of beef. Other results from this experiment (Alexander et al., 2007b), indicate QTL with additive effects on the quantities for saturated, mono-unsaturated and poly-unsaturated fatty acids in steaks from the Longissimus dorsi. However, only the QTL on BTA2 reported here were coincident with those previously reported.
Excess dietary saturated fatty acids are thought to predispose people to several cardiovascular diseases (e.g., Department of Health, 1994; Katan et al., 1994; 2000). Results of this study indicate that the relative amounts of SFA, MUFA, and PUFA are under a degree of genetic control. Thus, it may be possible to manipulate fatty acid composition using genetic markers and appropriate specialized crossbreeding systems in order to improve healthfulness of beef.
Acknowledgements
The authors thank L. French, V. Leesburg, C. Murrieta, K. Neary and H. Stroh for excellent technical assistance. We also thank J.J. Reeves and C.T. Gaskins at Washington State University for initiating this population. USDA-ARS is an equal opportunity/affirmative action employer. All agency services are available without discrimination. This research was conducted under cooperative agreements between USDA-ARS, Montana Agric. Exp. Sta., and the ARC. Mention of a proprietary product does not constitute a guarantee or warranty of the product by USDA, Montana Agric. Exp. Sta., ARC, or the authors and does not imply its approval to the exclusion of other products that also may be suitable.
References
Alexander, L.J., Geary, T.W., Snelling, W.M. & MacNeil, M.D., 2007a. Quantitative trait loci with additive effects on growth and carcass traits in a Wagyu-Limousin F2 population. Anim. Genet. 38, 413-416. [ Links ]
Alexander, L.J., MacNeil, M.D., Geary, T.W., Snelling, W.M., Rule, D.C. & Scanga, J.A., 2007b. Quantitative trait loci with additive effects on palatability and fatty acid composition of meat in a Wagyu-Limousin F2 population. Anim. Genet. 38, 413-416. [ Links ]
Ausubel, F., Brent, M.R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. & Struhl. K., 1994. Current Protocols in Molecular Biology 1. John Wiley & Sons, Inc. New York, USA. [ Links ]
Cheverud, J.M., 2001. A simple correction for multiple comparisons interval mapping genome scans. Heredity 87, 52-58. [ Links ]
Christie, W.W., 1982. The preparation of derivatives of lipids. In: Lipid Analysis, 2nd ed. Pergamon Press, New York, USA. [ Links ]
Darvasi, A. & Soller, M., 1997. A simple method to calculate resolving power and confidence interval for QTL map location. Behavior Genet. 27, 125-132. [ Links ]
Department of Health, 1994. Nutritional Aspects of Cardiovascular Disease. Report of the Cardiovascular Review Group of the Committee on Medical Aspects of Food Policy. Report on Health and Social Subjects 46. London: HMSO. [ Links ]
Dryden, F.D. & Marchello, J.A., 1970. Influence of total lipid and fatty acid composition upon the palatability of three bovine muscles. J. Anim. Sci. 31, 36-41. [ Links ]
Green, P., Falls, K. & Crooke, S., 1990. Documentation for CRI-MAP. Ver2.4. Washington School of Medicine, St. Louis, MO, USA. [ Links ]
Katan, M.D., 2000. Nutritional interventions: the evidence. Proc. Nutr. Soc. 59, 417-418. [ Links ]
Katan, M.B., Zock, P.L. & Mensink, R.P., 1994. Effects of fatty acids on blood lipids in humans: an overview. Am. J. Clin. Nutr. 60, 1017S-1022S. [ Links ]
Kramer, J.K.G., Fellner, V., Dugan, M.E.R., Sauer, F.D., Mossoba, M.M. & Yurawecz, M.P., 1997. Evaluating acid and base catalysts in the methylation of milk and rumen fatty acids with special emphasis on conjugated dienes and total trans fatty acids. Lipids 32, 1219-1228. [ Links ]
May, S.G., Sturdivant, C.A., Lunt, D.K., Miller, R.K. & Smith, S.B., 1993. Comparison of sensory characteristics and fatty acid composition between Wagyu crossbred and Angus steers. Meat Sci. 35, 289-98. [ Links ]
McPherron, A.C. & Lee, S.J., 1997. Doubling muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA. 94, 12457-12461. [ Links ]
McPherron, A.C., Lawler, A.M. & Lee, S.J., 1997. Regulation of skeletal muscle mass in mice by a new TGF-ß super family member. Nature 387, 83-90. [ Links ]
Melton, S.L., Amiri, M., Davis, G.W. & Backus, W.R., 1982. Flavor and chemical characteristics of ground beef from grass-, forage- grain- and grain finished steers. J. Anim. Sci. 55, 77-87. [ Links ]
Mir, P.S., Mir, Z., Kuber, P.S., Gaskins, C.T., Martin, E.L., Dodson, M.V., Calles, J.A.E., Johnson, K.A., Busboom, J.R., Wood, A.J., Pittenger, G.J. & Reeves, J.J., 2002. Growth, carcass characteristics, muscle conjugated linoleic acid (CLA) content, and response to intravenous glucose challenges in high percentage Wagyu, Wagyu x Limousin, and Limousin steers fed sunflower oil-containing diets. J. Anim. Sci. 80, 2996-3004. [ Links ]
Mir, Z., Paterson, L.J. & Mir, P.S., 2000. Fatty acid composition and conjugated linoleic acid content of intramuscular fat in crossbred cattle with and without Wagyu genetics fed a barley based diet. Can. J. Anim. Sci. 80, 195-197. [ Links ]
Murrieta, C.M., Hess, B.W. & Rule, D.C., 2003. Comparison of acidic & alkaline catalysts for preparation of fatty acid methyl esters from ovine muscle with emphasis on conjugated linoleic acid. Meat Sci. 65, 523-529. [ Links ]
Pitchford, W.S., Deland, M.P.B., Siebert, B.D., Malau-Aduli, A.E. & Bottenma, C.D.K., 2002. Genetic variation in fatness and fatty acid composition of crossbred cattle. J. Anim. Sci. 80, 2825-2832. [ Links ]
Raes, K., De Smet, S. & Demeyer, D., 2001. Effect of double-muscling in Belgian Blue young bulls on the intramuscular fatty acid composition with emphasis on conjugated linoleic acid and polyunsaturated fatty acids. Anim. Sci. 73, 253-260. [ Links ]
Rule, D.C., Broughton, K.S., Shellito, S.M. & Maiorano, G., 2002. Comparison of muscle fatty acid profiles and cholesterol concentrations of bison, beef cattle, elk, and chicken. J. Anim. Sci. 80, 1202-1211. [ Links ]
Seaton, G., Haley, C.S., Knott, S.A., Kearsey, M. & Visscher, P.M., 2002. QTL Express: mapping quantitative trait loci in simple and complex pedigree. Bioinformatics 18, 339-340. [ Links ]
Sellick, G.S., Pitchford, W.S., Morris, C.A., Cullen, N.G., Crawford, A.M., Raadsma, H.W. & Botterma, C.D.K., 2007. Effect of myostatin F94L on carcass yield in cattle. Anim. Genet. 38, 440-446. [ Links ]
Smith, T.P.L., Lopez-Corrales, N.L., Kappes, S.M. & Sonstegard, T.S., 1997. Myostatin maps to the interval containing the bovine mh locus. Mamm. Genome 8, 742-744. [ Links ]
Sturdivant, C.A., Lunt, D.K., Smith, G.C. & Smith, S.B., 1992. Fatty acid composition of subcutaneous and intramuscular adipose tissues and M. longissimus dorsi of Wagyu cattle. Meat Sci. 32, 449-458. [ Links ]
Thallman, R.M., Bennett, G.L., Keele, J.W. & Kappes, S.M., 2001a. Efficient computation of genotypes probabilities for loci with alleles. I. Allelic peeling. J. Anim. Sci. 79, 26-33. [ Links ]
Thallman, R.M., Bennett, G.L., Keele, J.W. & Kappes, S.M., 2001b. Efficient computation of genotype probalities for loci with many alleles. II. Iterative method for large, complex pedigrees. J. Anim. Sci. 79, 34-44. [ Links ]
Westerling, D.B. & Hedrick, H.B., 1979. Fatty acid composition of bovine lipids as influenced by diet, sex and anatomical location and relationship to sensory characteristics. J. Anim. Sci. 48, 1343-1348. [ Links ]
Wood, J.D., Richard, R.I., Nute, G.R., Fhisher, A.V., Campo, M.M., Kasapidou, E., Sheard, P.R. & Enser, M., 2004. Effect of fatty acids on meat quality: a review. Meat Sci. 66, 21-32. [ Links ]
Yamasake, M., Kishihara, K., Ikeda, I., Sugano, M. & Yamada, K., 1999. A recommended esterification method for gas chromatographic measurement of conjugated linoleic acid. J. Am. Oil Chem. Soc. 76, 933-938. [ Links ]
 Correspondence:
Correspondence:
Email: mike.macneil@ars.usda.gov


{kind=link}