










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.2 Pretoria Feb. 2008
Effect of sage extract (Salvia officinalis) on growth performance, blood parameters, oxidative stress and DNA damage in partridges
S. YurtsevenI; M. ÇetinI; T. ŞengülI; B. SögütII
IHarran University, Agriculture Faculty, Department of Animal Science, Sanliurfa, Turkey
IIYüzüncü Yil University, Agriculture Faculty, Department of Animal Science, Van, Turkey
ABSTRACT
This study was performed to evaluate the effect of different doses of sage extract on the growth and blood parameters, oxidative stress and DNA damage in partridges. In total, 252 day-old partridges (Alectoris chukar) were used. The birds were divided into four groups: 0.1% flavomycin was included in the diet of the control group (I) while 2.5, 5.0 and 7.5 mL sage extract/kg were included in the diets of treatment groups II, III and IV, respectively. At the end of the experiment no significant differences between treatments were observed in live weight, live weight gain, feed consumption, feed conversion ratio, carcass weight and survival rate. In addition, blood analyses indicated that the differences between groups in the amounts of plasma cholesterol, triglyceride high density lipoprotein (HDL), very low density lipoprotein (VLDL) and alkaline phosphates (ALP) were not significantly different. Furthermore, treatments did not affect total sulphydryl (SH) and lipid hydroperoxide (LOOH) concentrations, total antioxidant response or the total oxidant status (TOS). However, sage extract significantly decreased DNA damage in a linear, dose-dependent manner whilst the antibiotic, flavomycin, elevated the oxidative stress index (OSI) and resulted in DNA damage. It was concluded that supplementing sage extract in the partridge diet at the doses studied does not lead to a negative effect on the growth performance of these birds.
Keywords: Salvia officinalis, Alectoris chukar, growth performance, blood parameters, DNA damage
Introduction
The use of antibiotics as growth promoters in poultry diets has been prohibited by the European Union in 2002 due to the risk of residues, side effects and the promotion of bacterial resistance to antibiotics. This has lead to a reduction in their use in poultry diet in other countries (Hinton, 1988; Narayanankuti et al., 1992; Ceylan et al., 2003). Consequently, research has been focused on the inclusion of natural plant extracts in diets of birds as alternatives to antibiotics to combat disease and enhance the performance of the birds. Positive results have been reported (Jamroz & Kamel, 2002; Parlat et al., 2005).
Sage plants belong to the Lamiaceae family and are known to be inhibitors of inflammation, suppressors of blood glucose levels and strong antioxidants. Salvia officinalis L. is a native aromatic and medicinal plant, and grows in countries bordering the Mediterranean Sea (Cuvelier et al., 1994; Miliauskas et al., 2004). Rosmarinic and carnosic acids, which have antioxidant properties, are present at high concentrations in the extract of sage plants (Cuvelier et al., 1994). Studies on the leaves, roots and/or water soluble extractions of S. officinalis L. have shown that they contain 1 - 2.5% volatile fatty acids, saponins, diterpenes, flavonoids, phenolic acids, salviatannins, resin and oestrogenic substances (Dogan, 2004; Slamenova et al., 2004; Lima et al., 2005). Experiments examining the effect of the essential oil from this species on rats have been done, but little information is available on their effects in poultry (Lima et al., 2004). The use of higher doses of S. officinalis L. is not recommended because its essential oil contains tannins, toxic thujone and eugenol compounds (Cuppet & Hall, 1998; Dogan, 2004; Lima et al., 2004). However, the dietary level at which sage extract becomes toxic to poultry has not been established.
The use of antibiotics and exposure to synthetic materials may result in the release of free radicals in the body and that can cause cancer. Free radicals occur due to metabolic processes in body tissues and their abundance can be controlled by ensuring an optimal antioxidative status of the body. However, under some conditions such as poor hygiene and disease, oxidative stress cannot be controlled by the body. Under these conditions, DNA damage can lead to changes in the genetic code in cells, mutation and cell death (Floyd, 1990). According to Vujosevic & Blagojevic (2005) the essential oil of sage and its fractions contain substances with antimutagenic potential against ultraviolet-induced and possibly spontaneous mutations. For this reason, sage extracts have the capacity to help an organism's antioxidant systems, and prevent diseases. In this role, plant extracts could be used as a natural and alternative means of preventing oxidation.
The objective was to determine the effects of including sage extract in the diet of growing partridges on their growth, survival, blood parameters, oxidative stress and DNA damage.
Materials and Methods
Dried sage was ground in a knife mill to particle sizes of 1 - 3 mm, and was separated into size fractions using sieves. A hydroalcoholic extract was prepared using food grade ethanol and deionised water (96 and 4%, respectively). The mixture was placed in a sealed glass vial in a heated water-bath shaker for extraction. After extraction the glass vial was removed and left to cool for 10 min. The mixture was vacuum filtered and the solid residue and hydroalcoholic extract collected. Evaporation of the hydroalcoholic solvent under reduced pressure gave an extract of 15% dry sage which contained 7% rosmarinic acid, 10.6% carnosic compounds and 1.73% essential oil.
A study was conducted using 252 partridges (Alectoris chukar) consisting of equal numbers of males and females. They were kept together in mixed-sex pens during the experiment. Day-old partridges were weighed and divided into groups on a weight basis to form four treatment groups (n = 63), each with three replications (cages) of 21 chicks per cage. Body weights were equated and divided into groups at the beginning of the experiment.
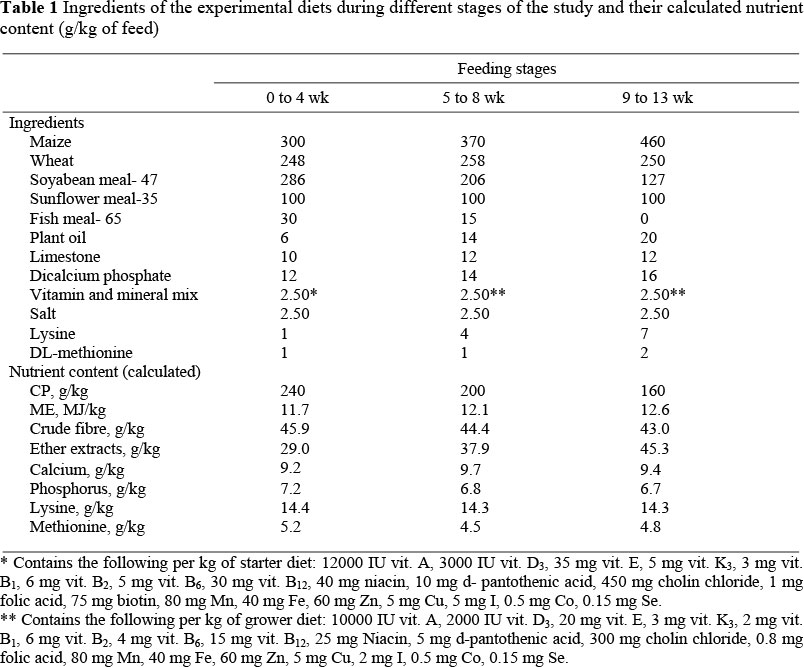
The experiment lasted three months, between May and June. During the first week of life, the birds were reared in brooders before being placed in floor pens with wood shaving as litter. The light program was 23.5 h light until seven days of age and then 18 h light : 6 h dark. The treatment diets were fed ad libitum and the birds had free access to water. Feed formulation and calculated analysis are presented in Table 1.
The control group received the basal diet with 0.1% flavomycin whilst treatment groups II, III and IV received the basal diet with the inclusion of 2.5, 5.0 or 7.5 mL sage extract/kg, respectively. Information on the effects of sage extract on poultry performance is limited, but Galobart et al. (2000) found that low levels of the oil (2 mL/kg diet or less) had only a small effect on animal performance. Consequently, higher doses were selected in the present study. Treatment diets were prepared by adding one kg/ton of pre-diluted sage extract to the basal diet. Live weight (LW) and feed consumption (FC) of the partridges were recorded weekly for the first month and then every second week until the end of the experiment. At the end of the trial, 27 birds from each group were slaughtered to determine carcass weight (CW). These birds were also used for the collection of blood samples which were harvested and analyzed in the biochemistry laboratory of the University of Harran.
After overnight fasting, peripheral blood samples (total 6 mL) were collected from the antecubital vein into heparinised tubes, stored at 10 °C in the dark to prevent further DNA damage and processed within 2 h. Mononuclear leukocyte isolation for the comet assay was performed, using Histopaque 1077 (Sigma). The remaining blood was centrifuged at 1500 * g for 10 min to obtain plasma. The separated plasma was stored at -80 °C until further analysis of lipid and other oxidative stress parameters.
The comet assay was performed as described by Singh et al. (1988), with the following modifications: The images of randomly chosen nuclei (50 nuclei from each of two replicate slides) were analyzed visually for each subject. Each image was classified according to the intensity of the fluorescence in the comet tail and was given a value of 0, 1, 2, 3 or 4 (from undamaged DNA, class 0, to maximally damaged DNA, class 4), so that the total score of a slide could be between 0 and 400 arbitrary units (AU).
Plasma lipid hydroperoxides (LOOH) were evaluated by an automated xylenol orange method based on that described by Jiang et al. (1991) in which an acidic medium, Fe++, is oxidized to Fe+++ by lipid hydroperoxides. The resulting values were measured in µmol/L.
The measurement of total antioxidant response (TAR) reflects the antioxidative status of the plasma. The TAR of the plasma was determined using a novel automated measurement method, developed by Erel (2005). In this assay a standardized solution of Fe2+-o-dianisidine complex reacts with a standard solution of hydrogen peroxide via a Fenton-type reaction, producing hyroxyl radicals. This potently reactive oxygen oxidizes the reduced colourless o-dianisidine to yellow-brown coloured dianisidyl at low pH values. The results of this procedure were expressed as mmol Trolox equivalent/L. Total peroxide concentrations were determined using the "FOX2" method (Miyazawa, 1989) with minor modifications. The FOX2 test system is based on the oxidation of a ferrous ion to ferric by various types of peroxides contained within the plasma samples, to produce a coloured ferric-xylenol orange complex whose absorbance can be measured. The results were expressed as mmol H2O2/L Trolox equivalent/L. The oxidative stress index (OSI) was determined by dividing TAR by the total oxidant status (TOS) (TAR/TOS; Erel, 2005).
The enzymatic activity of caeruloplasmin was measured according to the method published by Erel (1998) in which ferrous ions are oxidized to ferric ions via caeruloplasmin ferroxidase activity. The oxidized ion amount is related to the enzymatic activity and the values are expressed in U/L. Free sulphydryl groups (-SH) in the serum samples were assayed according to the method of Ellman (1959) as modified by Hu et al. (1993). The levels of triglyceride (TG), total cholesterol (TC), high density lipoprotein (HDL), low density lipoprotein (LDL) and alkaline phosphate (ALP) were determined using commercially available assay kits (Abbott, Chicago, IL, USA) with an Abbott Aeroset auto-analyzer (Chicago, IL, USA).
Data were analyzed with one-way analyses of variance and significant differences between means were determined by the Duncan's multiple range test (SPSS, 1999). When the F-test was significant, single degree of freedom orthogonal contrasts were used to determine linear, quadratic and cubic effects of the level of sage extract. Because no biologically significant cubic effects were detected, only the results of linear and quadratic contrasts are reported in Table 2, and again, since no biologically significant quadratic and cubic effects were detected, only the results of linear contrasts are reported in Tables 3 and 4.
Results
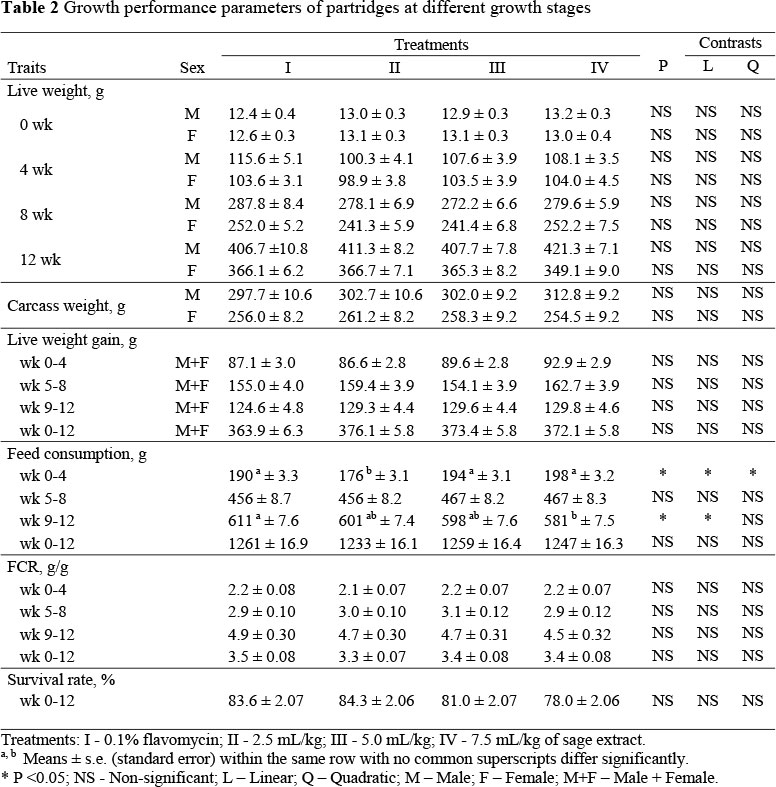
The LW, CW, live weight gain (LWG), FC, feed conversion ratio (FCR) and survival rate of partridges during the experiment are presented in Table 2.
The live weights and carcass weights data from male and female birds were analyzed separately, but data from the sexes were combined for the analysis of live weight gains. There were no significant differences between the treatments in LW, LWG, CW, FC, FCR and survival rate. However, feed consumption was reduced (P <0.05) in a linear, dose dependent manner by the inclusion of sage extract between weeks 9 - 12 of age.
Live weights were not significantly affected by the treatments and remained similar at the end of the trial period, ranging from 406.7 to 421.3 g for males and 349.1 to 366.7 g for females. The different sage extract doses also did not significantly affect LWG during the period 0 - 12 weeks of age. The lowest and highest LWG in the treatment groups were 363.9 and 376.1 g, respectively. The CW of male and female partridges were not significantly different at the end of the treatment period.
Mean FC differed (P <0.05) between weeks 0 - 4 and 9 - 12 but was similar between weeks 5 - 8 and 0 - 12. The mean FC ranged from 1233 to 1261 g. No significant treatment effects on FCR were observed which ranged from 3.3 to 3.5 during the full experimental period (weeks 0 - 12 wk; Table 2). The level of sage extract also did not affect the survival rate which was recorded as 83.6, 84.3, 81.0 and 78.0% during weeks 0 - 12 in groups I, II, III and IV, respectively. Treatment and sex did not significantly interact to affect LW, LWG, FC or FCR.
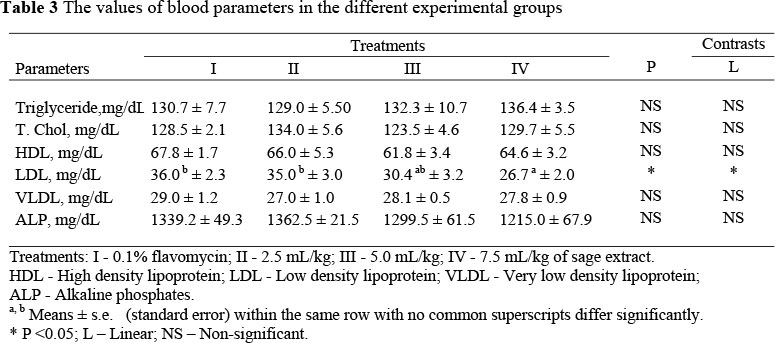
The blood analyses showed that the treatment imposed, did not significantly affect the levels of triglyceride, total cholesterol, HDL, VLDL or ALP. However, the LDL rate was lower (P <0.05) in the treatment groups than in the control group (P <0.05), and, within the supplemented groups, the level of LDL declined linearly with stepwise increases in extract dose. The values of some blood parameters are shown in the Table 3.
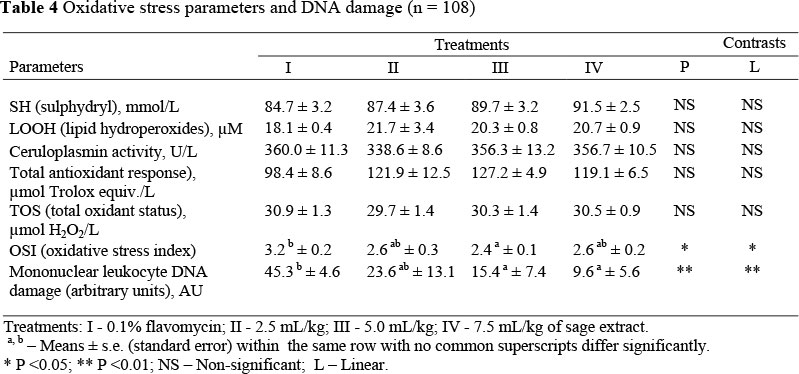
No significant differences between treatments were observed in SH, LOOH, caeruloplasmin activity, TAR or TOS as oxidative stress parameters. However, OSI values were reduced significantly. Although not significant, the lowest TAR value was observed in the antibiotic supplemented group. The highest level of DNA damage was also detected in this group (Table 4), whilst significant decreases in DNA damage were suggested to occur as a result of supplementation with the extract. The higher the dose of extract, the greater was the beneficial effect of its inclusion (P <0.01, Figure 1).
Discussion
Information on the effects of sage extract on poultry is limited and no advice is available regarding suitable dose levels. The minimum inhibitory concentration (MIC) value of sage oil is > 7.5 iiL/mL for many bacteria (Schwiertz et al., 2006). The reason for using sage extract doses of greater than 2 mL/kg was to eliminate bacterial action and to allow the expression of the antioxidant properties of this supplement. The doses used in this study did not significantly affect the performance of the partridges. As such there were no treatment effects on LW, LWG, CW, FC, FCR or survival rate during weeks 0 - 12 of age. A significant and linear reduction in FC due to sage extract occurred in weeks 9 to 12. This may have been due to the strong smell and bitter taste of the extract since these are properties known to characterize antioxidants in sage plants. However, these characteristics did not significantly affect the performance of the birds.
The results from this study indicated that sage extract had a positive effect on antioxidant status in the body, as confirmed by the reduction in OSI value, an indicator of oxidative stress. Kuzmenko et al. (1999) have previously reported that a diet supplement with 6% carnosic acid resulted in a significant reduction in lipid peroxide in the liver.
In the present study it appears that the sizeable antioxidant effect of sage extract did not originate from protein oxidation and reduced lipid peroxidation since the concentration of thiol groups (SH) and lipid peroxide (LOOH), their indicator, were unaffected by the extract. In addition, the level of fat (30 - 40 g/kg) in the diet fed to the groups was not high enough to absorb a high level of lipids. Furthermore, there were no significant differences between groups in caeruloplasmin activity. The mechanism driving the absorption of phenolic compounds in sage and/or other plant extracts has not yet been adequately determined in either poultry or mammals. However, it has been suggested that the process operates through affecting antioxidant enzymes, especially in the liver. Lima et al. (2005) speculated that supplemented sage extract in the diet of rats caused significant increases in the activity of glutathione-S-transferase and glutathione reductase antioxidant enzymes in the liver. In vivo and in vitro observations of liver cells support these findings. Researchers underlined that the impact of sage extract is derived from compounds in its essential oil. Lopez-Bote et al. (1998) indicated that supplemented sage extract in the diet of the broilers prevented lipid oxidation in the carcass for a prolonged period of time, and so it helped to extend the shelf-life of meat.
Synthetic antioxidants have thus far been preferred as they are effective and inexpensive, but recently it has been shown that they can cause cancer. Using tocopherols as natural antioxidants instead of synthetics has not proved popular since their action is short-lived (Akgül, 1989; Akgül & Ayar, 1993). In the current study the weak plasma antioxidant effect of sage extract tended to reduce DNA damage in supplemented birds, where one of the main causes of DNA damage is oxidative stress. Other reasons for DNA damage are different chemical compounds and medicines. Partridges may offer a good model in which to observe the plasma antioxidant effect of sage extract if they are under significant stress. The linear dose-response relationship in which a reduction in DNA damage resulted from the use of sage oil indicates that the extract thus has the potential to reduce oxidation, DNA damage and, through this, cancer.
The most important anti-pathogenic effects of antibiotics as growth factor are to destroy bacterial DNA. However, they are also known to destroy the DNA of the host. For instance, it has been determined that penicillin, used widely, and part of the biggest antibiotic group, causes a 50% increase in DNA damage in lymphocytes, which are the defence system in children (Dündaröz et al., 2001). In addition, it has been reported that some antibiotics inhibit the action of the products of cell DNA polymerase enzymes (Huynh et al., 1987; Neftel et al., 1987). Observing the highest DNA damage in the group supplemented with flavomycin in the present study is an indicator of this effect (Figure 1). Additionally, it was observed that the antioxidative defence system was weakened in the antibiotic group. Similarly, Sirvydis et al. (2003) reported that broilers supplemented with plant extracts instead of flavomycin had a better performance. However, even though antioxidant effects of the sage extract were observed, hypocholesterolemic effects were not.
Conclusion
In conclusion, the present study shows that sage extract included as a growth factor in the diet of partridges does not lead to negative effects on performance. The extract significantly reduced DNA damage in leukocyte cells, but did not affect lipid profiles or oxidative stress parameters. The levels of sage extract used in this experiment were not toxic, but using higher levels of sage extract could be toxic.
References
Akgül, A., 1989. The antioxidant effects of spices. Doga-Tr. J. Agric. Forest. 13, 11-24. [ Links ]
Akgül, A. & Ayar, A., 1993. The antioxidant effects of domestic spices. Doga-Tr. J. Agr. Forest. 17, 1061-1068. [ Links ]
Ceylan, N., Ciftci, I. & Ilhan, Z., 2003. The Effects of some alternative feed additives for antibiotic growth promoters on the performance and gut microflora of broiler chicks. Turk J. Vet. Anim. Sci. 27, 727-733. [ Links ]
Cuppet, S.L. & Hall, C.A., 1998. Antioxidant activity of labiate. In: Advances in Food and Nutrition Research. Academic Pres. London. pp. 245-271. [ Links ]
Cuvelier, M.E., Berset, C. & Richard, H., 1994. Antioxidant constituents in sage (Salvia officinalis). J. Agric. Food Chem. 42, 665-669. [ Links ]
Dogan, Y., 2004. A source of healing from past to today: Salvia officinalis. J. Nature Environ. Culture, No. 3, Izmir. [ Links ]
Dündaröz, R., Ulucan, H., Denli, M., Karapinar, K., Aydin, H.I. & Baltaci, V., 2001. Evaluation of DNA damage using the comet assay in children on long-term benzathine penicillin for secondary prophylaxis of rheumatic fever. Pediatrics Int. 43, 276-280. [ Links ]
Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 82, 70-77. [ Links ]
Erel, O., 1998. Automated measurement of serum ferroxidase activity. Clin. Chem. 44 (11), 2313-2319. [ Links ]
Erel, O., 2005. A new automated method to measure total antioxidant response against potent free radical reactions. Clin. Biochem. 37, 112-119. [ Links ]
Floyd, R., 1990. Role of oxygen free radicals in carcinogenesis and brain ischemia. Faseb. J. 4, 2587-2597. [ Links ]
Galobart, J., Barroeta, A.C., Baucells, M.D., Codony, R. & Ternes, W., 2000. Effect of dietary supplementation with rosemary extract and a-tocopheryl acetate on lipid oxidation in eggs enriched with ω3-fatty acids. Poult. Sci. 80, 460-467. [ Links ]
Hinton, M.H., 1988. Antibiotics, Poultry production and public health. World's Poult. Sci. 44, 67-69. [ Links ]
Hu, M.L., Louie, S., Cross, C.E., Motchnik, P. & Halliwell, B., 1993. Antioxidant protection against hyochlorous acid in human plasma. J. Lab. Clin. Med. 121 (2), 257-262. [ Links ]
Huynh, U.D., Neftel, K.A., Spadari, S. & Hubscher, U., 1987. Betalactam antibiotics interfere with eukaryotic DNA-replication by inhibiting DNA polymerase alfa. Nucleic Acids Res. 15, 495-506. [ Links ]
Jamroz, D. & Kamel, C., 2002. Plant extracts enhance broiler performance. In: Non-Ruminant Nutrition: Antimicrobial agents and plant extract on immunity, health and performance. J. Anim. Sci. 80, (E. Suppl 1), p. 41. [ Links ]
Jiang, Z.Y., Wollard, A.C.S. & Wolff, S.P., 1991 Lipid hyroperoxides measurement of Xylenol orange, comparison wit TBA assay and iodometric method. Lipids 26, 853-856 [ Links ]
Kuzmenko, A.I., Morozova, R.P., Nikolenko, I.A., Donchenko, G.V., Richheimer, S.L. & Bailey, D.T., 1999. Chemiluminiscence determination of the in vivo and in vitro antioxidant activity of RoseOx and carnosic acid. J. Photochem. Photobiol. 48, 63-67. [ Links ]
Lima, C.F., Andrade, P.B., Rosa, M.S., Fernandes, M.F. & Wilson, C.P., 2005. The drinking of Salvia officinalis infusion liver antioxidant status in mice and rats. J. Ethnopharm. 97, 383-389. [ Links ]
Lima, C.F., Carvalho, F., Fernandes, E., Bastos, M.L., Santos-Gomes, P.C., Fernandes, Ferreria, M. & Pereira-Wilson, C., 2004. Evaluation of toxic/protective effects of essential oil of Salvia officinalis on freshly isolated rat hepatocytes. Toxicology in Vitro 18, 457-465. [ Links ]
Lopez-Bote, C.J., Gray, J.I., Gomaa, E.A. & Flegal, C.J., 1998. Effect of dietary administration of oil extracts from rosemary and sage on lipid oxidation in broiler meat. Br. Poult. Sci. 2, 235-240. [ Links ]
Miliauskas, G., Venskutonis, P.R. & Van Beek, T., 2004. Screening of radical scavenging activity of some medicinal and aromatic plant extract. Food Chem. 85, 231-237. [ Links ]
Narayanankutty, K., Ramakrishnan, A. & Visvannath, A., 1992. Efficacy of virginamycin as growth promoter in commercial broiler chicks. J. Vet. Anim. Sci. 23, 96-97. [ Links ]
Neftel, K.A. & Hubscher, U., 1987. Effects of beta-lactam antibiotics on proliferating eucaryotic cells. Antimicrob. Agents Chemother. 31, 1657-1661. [ Links ]
Parlat, S.S., Yildiz, A.O., Olgun, O. & Cufadar, Y., 2005. Usage of oregano essential oil (Origanum vulgare L.) extract for growth stimulant antibiotics in quail rations. Selcuk Univ., J. Fac. Agric. 36, 7-12. [ Links ]
Schwiertz, A., Duttke, C., Hild, J. & Müller, H.J., 2006. In vitro activity of essential oils on microorganisms isolated from vaginal infections. Int. J. Aromather. 16, 169-174. [ Links ]
Singh, P.N., McCoy, M.T., Tice, R.R. & Schneider, E.L., 1988. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell. Res. 175, 184-191. [ Links ]
Sirvydis, V.H., Bobiniene, R., Priudokiene, V. & Vencius, D., 2003. Phytobiotics add value to broiler feed. World Poult. 19 (19), 16-17. [ Links ]
Slamenova, D., Masterova, I., Labaj, J., Horvathova, H., Kubala, P., Jakubikova, J. & Wsolova, L., 2004. Cytotoxic and DNA-damaging effects of diterpenoid quinones from the roots of Salvia officinalis L. on colonic and hepatic human cells cultured in vitro. Basic Clin. Pharmacol. Toxicol. 94, 282-290. [ Links ]
SPSS, 1999. SPSS Applications guide (Base 10.0). SPSS Inc., Chicago, IL., USA. [ Links ]
Vujosevic, M. & Blagojevich, J., 2005. Antimutagenic effects of extracts from sage (Salvia officinalis) in mammalian system in vivo. Arch. Biol. Sci., Belgrade 57, 163-172. [ Links ]
 Correspondence:
Correspondence:
E-mail: tsengul2001@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}