










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.2 Pretoria Feb. 2008
Heritability estimates and correlations between production and reproductive traits in Lori-Bakhtiari sheep in Iran
M. Vatankhah; M.A. Talebi
Department of Animal Science, Agriculture and Natural Resources Research Centre, P.O. Box 415, Shahrekord, Iran
ABSTRACT
Heritablities and correlations were estimated between lamb body weight at different ages and reproductive traits in the Lori-Bakhtiari sheep breed. Data and pedigree information for Lori-Bakhtiari sheep used in this study were 5826 records of body weight of lambs from 240 sires and 1627 dams, and 5741 records of reproductive traits from 1797 ewes collected from 1989 to 2006 from a Lori-Bakhtiari flock at the Shooli station in Shahrekord. The lamb traits investigated were weights at birth (BWT), weaning (WWT) and at six months of age (WT6). The reproductive traits were conception rate (CR), litter size at birth per ewe lambing (LSB/EL), litter size at weaning per ewe lambing (LSW/EL), litter size at birth per ewe exposed to a ram (LSB/EE), litter size at weaning per ewe exposed (LSW/EE), total litter weight at birth per ewe lambing (TLWB/EL), total litter weight at weaning per ewe lambing (TLWW/EL), total litter weight at birth per ewe exposed (TLWB/EE) and total litter weight at weaning per ewe exposed (TLWW/EE). (Co)Variance components and genetic parameters were estimated using the restricted maximum likelihood procedure applying three multi-trait animal models. The estimates of direct heritability for lamb body weights were 0.31 ± 0.02 at birth, 0.10 ± 0.02 at weaning and 0.19 ± 0.02 at six months of age, and estimates of maternal heritability were 0.22 ± 0.02, 0.16 ± 0.02 and 0.04 ± 0.02, respectively. The estimates of heritability for reproductive traits varied from 0.02 ± 0.01 for CR to 0.23 ± 0.01 for TLWB/EL, and repeatability estimates for reproductive traits ranged from 0.11 for CR to 0.28 for LSB. Genetic correlations among lamb body weight and various reproductive traits were positive and varied from 0.06 to 0.98. Phenotypic correlations were lower than genetic correlations (0.01 to 0.20). The estimates of genetic correlations between lamb body weight with CR and litter size at birth and at weaning were low to moderate, while those between lamb body weight with TLBW and TLWW per ewe lambing and per ewe exposed were positive and very high. Thus, lamb body weight at weaning or at six months of age could be considered as selection criteria to indirectly improve reproductive traits in Lori-Bakhtiari sheep because the heritability of lamb body weight was more than that of reproductive traits and can be measured at an early stage in the life of the animal.
Keywords: Genetic correlations, early growth traits, reproductive traits
Introduction
Improved ewe productivity is a major objective in the sheep industry in Iran. Several traits have been used as indicators of ewe productivity. Litter size at birth is directly related to ovulation rate, though selection for this trait would not be enough to increase lamb production since it does not include lamb survival and weight at weaning. Litter size at weaning includes survival of lambs at weaning but not weight (Snyman et al., 1997). On the other hand, total litter weight of lambs weaned per ewe exposed combines the ewe's fertility, survival rate and growth performance of the lambs from birth to weaning. Therefore, this trait is the most important factor in determining ewe productivity and the economic efficiency of a lamb production enterprise (Boujenane et al., 1991). Genetic improvement of ewe productivity could be achieved either by direct selection for reproductive traits or by indirect selection for correlated traits that have high and positive genetic correlations with these traits. Direct selection for reproductive traits would be beneficial if these traits have high heritabilities. Estimates of heritability for reproductive traits in the literature are generally low (Fogarty, 1995; Safari et al., 2005). Another difficulty in direct selection of reproductive traits is the measurement of these traits, which can be done only at a relatively late stage in life. Therefore, early selection for these traits based on the animal's own performance is not possible (Snyman et al., 1998). Snyman et al. (1997; 1998) indicated that indirect selection for total weight of lambs weaned, based on the early recorded body weight traits of lambs could be more effective than direct selection in Afrino and Merino sheep. Hanford et al. (2002; 2003) suggested that selection based on weaning performance over a long period could result in a moderate positive response in litter size at birth and weaning, lamb weights at birth and weaning, but not in wool traits. The Lori-Bakhtiari sheep breed is one of the most common native breeds in the south western part of Iran (the Zagros Mountains), with a population of more than 1.7 million. The animals are kept mostly in villages under semi-intensive systems. The Ministry of Jahad-Agriculture in Iran has found it important to increase the efficiency of sheep production, because the output of sheep in this system is low. Due to the fact that reproductive traits are the most important traits in all sheep production systems (Gallivan, 1996; Vatankhah, 2005), increasing the efficiency of sheep production (output) by improving reproductive traits is economically important. To construct the breeding programmes for improvement of reproductive traits in sheep, heritability of reproductive traits, and genetic and phenotypic correlations of these traits with early growth traits of lambs should be estimated. This study was conducted to obtain such estimates for Lori-Bakhtiari sheep.
Materials and Methods
The data set used in this study consisted of 5826 records of birth weight, 5418 records of weaning weight, 4234 records of body weight at six months of age of lambs born to 240 sires and 1627 dams, and 5741 records of reproductive traits from 1797 ewes collected from 1989 to 2006 inclusive, from a Lori-Bakhtiari research flock at the Shooli station in Shahrekord. The flock was subjected to different selection criteria, generally related to increasing weaning weight (growth traits, total weaning weight per ewe exposed or Kleiber ratio) during this period. The means and measures of variations for the various traits used in the analyses are shown in Table 1.
The flock was managed under a semi-migratory or village system. From December to May the flock was usually kept at the station and was fed lucerne, barley and wheat stubble indoors. The rest of the year the sheep grazed range and cereal pastures. The breeding period extended from late August to late October (20 -25 ewes were assigned randomly to one ram) and consequently, lambing started in late January. Lambs suckled their dams and from 15 days of age had free access to a creep feed and were weaned at an average age of 90 ± 5 days. After weaning the male and female lambs were separated. Ewe lambs were kept on a pasture of cultivated lucerne, while the ram lambs were kept indoors and up to six months of age received a maintenance and growth diet ad libitum. The diet consisted of 45% lucerne hay, 39% barley, 7% beet pulp, 8% cottonseed meal, 1% salt and mineral supplements, and contained 135 g crude protein and 10.46 MJ metabolizable energy/kg.
Body weights of the lambs were recorded at birth (BWT), weaning (WWT) and at six months of age (WT6). Reproductive traits were conception rate (CR, measured as 1 or 0, for a ewe exposed to a ram that did or did not conceptive, respectively), litter size at birth per ewe lambing (LSB/EL, the number of fully formed lambs born per ewe lambing), litter size at weaning per ewe lambing (LSW/EL, the number of lambs alive at weaning), litter size at birth per ewe exposed (LSB/EE = CR x LSB/EL), litter size at weaning per ewe exposed (LSW/EE = CR x LSW/EL), total litter weight at birth per ewe lambing (TLWB/EL), total litter weight at weaning per ewe lambing (TLWW/EL), total litter weight at birth per ewe exposed (TLWB/EE = CR x TLWB/EL) and total litter weight at weaning per ewe exposed (TLWW/EE = CR x TLWW/EL).
The GLM procedure of SAS (1996) was applied to identify important fixed effects to be considered in the final model. The statistical model included year of lambing, age of dam (2 - 6 and > 7), sex of lamb (male and female), type of birth (single and multiple) and age (in days) as covariates for lamb weights at weaning and six months of age. For reproductive traits the factors included were year of ewe lambing and age of ewe (age of lamb was included as a covariate in the models used to analyze TLWW/EL and TLWW/EE). All the effects were significant (P <0.05) and hence they were included in the animal model. (Co)Variance components and correlations were estimated from an animal model in a bivariate analysis using the restricted maximum likelihood method (WOMBAT program of Meyer, 2006). Three different animal models were used. Tests of significance of each random effect were performed using log likelihood ratio tests after including each random effect (excluding residual) to the fixed effects model. An effect was considered significant when its inclusion in the model caused a significant increase in the log likelihood. A Chi-square distribution for a = 0.05 and appropriate degrees of freedom (1) were used as the critical test statistics. When -2 times the difference between log likelihood was greater than the critical value the inclusion of the effect was considered significant. When differences between log likelihoods were not significant the model with the fewest random effects was chosen. The following bivariate animal models were fitted:
yi= Xibi+ Ziai+ ei (M1)
yi= Xibi+ Ziai+ Wipei+ ei (M2)
yi= Xibi+ Ziai+ Wipei+Mimi+ ei (M3)
where, yi, bi, ai, pei, mi and ei are the vectors of observations, fixed effects, direct additive genetic effects, permanent environmental effects due to repeated records for reproductive traits, maternal additive genetic effects for lamb traits and residual random effects for the ith trait, respectively. Incidence matrices Xi, Zi, Wi and Mi related the observations of the ith trait to the respective fixed effects, additive genetic effects, permanent environmental effects and maternal genetic effects, respectively. The average information (AI) REML algorithm was use to maximize the likelihood (convergence criterion was 10-8) and additional restarts were performed until no further improvement in log likelihood occurred.
Results and Discussion
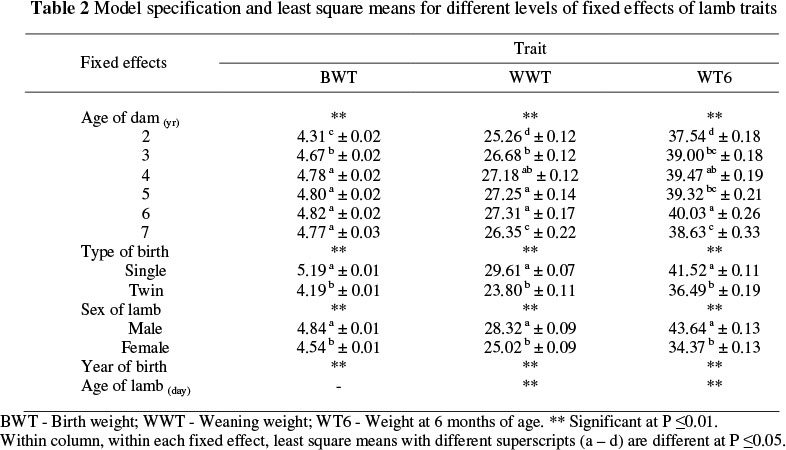
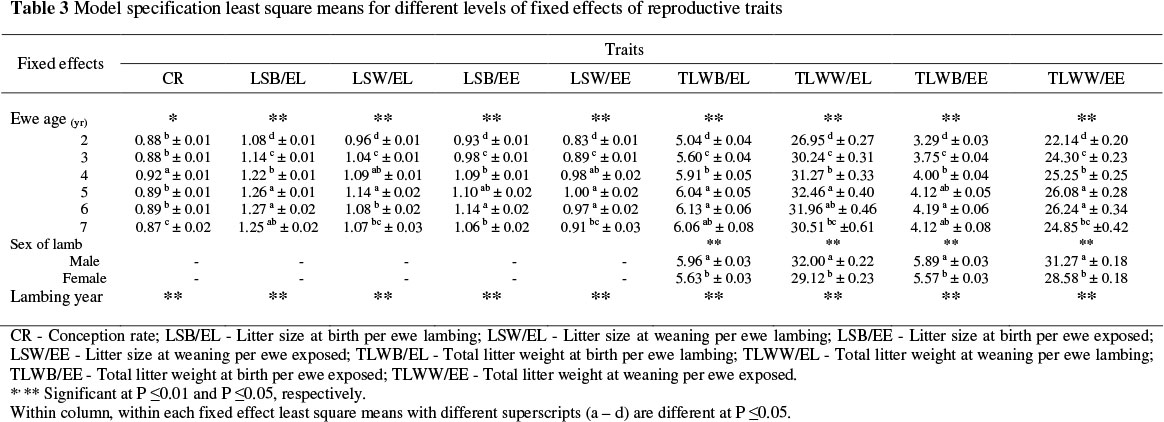
Model specification and least square means for different levels of fixed effects of lamb traits and reproductive traits are shown in Tables 2 and 3. The least square means of lamb traits increased with age of dam, and the highest means occurred in the 6-year old dam group. At all ages single-born and male lambs were heavier than multiple-born and female lambs. The least square means of reproductive traits increased with age of the ewe, the highest occurring in four to six years old ewes. Male lambs had a higher total litter weight at birth and weaning than female lambs (Table 3). Due to variations in climatic conditions, year of lamb birth and ewe lambing were highly significant on all lamb and reproductive traits.
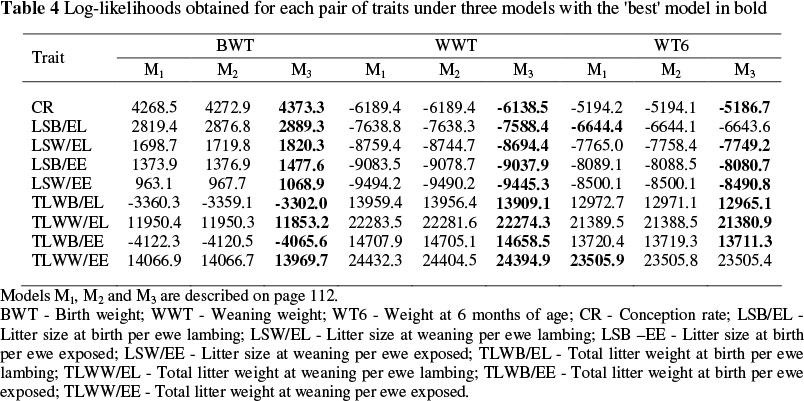
The log-likelihood obtained for each pair of traits under three different statistical models and the most appropriate model (in bold) from log-likelihood ratio tests are presented in Table 4. Model 3 which included a maternal genetic effect for lamb traits as well as a permanent environmental effect for reproductive traits was the most appropriate model for all sets of traits except for LSB/EL and TLWW/EE with weight at six months of age.
Estimates of direct heritability, maternal heritability, environmental variance as proportion of phenotypic variance and repeatability for lamb body weight and reproductive traits are given in Table 5. The estimate of direct heritability for BWT (0.31) was higher than those weighted means of heritability reported in the literature (Safari et al., 2005). Direct heritability at weaning (0.10) and six months of age (0.19) are in accordance with those reported in the literature (Fogarty, 1995; Safari et al., 2005). The lower direct heritability estimates in weight at three and six months of age compared with birth weight may be attributed to increased phenotypic variance due to the variation in mother's milk and environmental conditions (feed) which lambs received. The maternal heritability estimates of lamb body weight were lower than direct heritability estimates except for body weight at three months of age. The maternal heritability estimates of lamb body weight decreased with the age of the lamb, partly due to decreasing dependence of the lamb on mother's milk and partly due to the increasing additive genetic variance caused by a probable increase in expression of genes with age. These estimates and their trends with age were similar to the findings of Snyman et al. (1995), Näsholm & Danell (1996) and Matika et al. (2003).
The heritability estimate for CR (0.02) was lower than weighted means reported in the literature, but within the range of that of Fogarty (1995) and Safari et al. (2005). Conception rate is of great economic importance. However, because its heritability estimate is quite low, the genetic progress through within flock selection would be low. The estimated heritabilities for LSB/EL and LSW/EL (0.10) are similar to some estimates in the literature (Fogarty, 1995; Rosati et al., 2002). The heritability estimates for LSB/EE (0.02) and LSW/EE (0.01) are lower than the average of those reported in the literature (Fogarty, 1995; Safari et al., 2005). Therefore, improving multiple births in Lori-Bakhtiari sheep by genetic means would be slow. The heritability estimate for TLWB/EL was 0.23. Rosati et al. (2002) reported that the direct and maternal heritability for this trait were respectively 0.40 and 0.34. On the other hand, the corresponding estimate reported for Turkish Merino was lower at 0.05 (Ekiz et al., 2005). The heritability estimate of TLWW/EL (0.15) was higher than the weighted mean (0.11) reported by Safari et al. (2005). The TLWW/EL compared to the other reproductive traits could be considered the best criterium for selection because it measures the overall productivity of the ewes in terms of weights of lambs produced per parity, even though it does not take into account CR. The heritability estimate for TLWB/EE (0.16) was higher than the estimate (0.13) reported by Rosati et al. (2002). The heritability estimate of TLWW/EE was 0.12. The mean of heritability estimates reported for this trait is 0.13 (Fogarty, 1995; Safari et al., 2005). The trait has a low heritability estimate, possibly due to environmental effects such as creep feeding from 30 days of age until weaning. The TLWW/EE could be considered for selection purposes because it measures total productivity of ewes for lamb-meat production for a breeding year. The estimated fraction of variance due to permanent environmental effects for CR, LSB/El, LSW/EL, LSB/EE and LSW/EE was higher than heritability, while, for the other traits these estimates were lower than the heritability estimates.
Repeatability estimates for reproductive traits obtained in the current study ranged from 0.11 for CR to 0.28 for LSB and were similar to previously reported estimates (Fogarty, 1995). The heritability estimates for lamb body weight and reproductive traits obtained in this study indicated that firstly, lamb body weight traits gave higher responses to selection unlike reproductive traits, because they showed higher heritabilities. Secondly, selection based on lamb body weight gave faster genetic progress due to the fact that the body weight of lambs can be measured at an early stage in life, thus reducing generation interval.
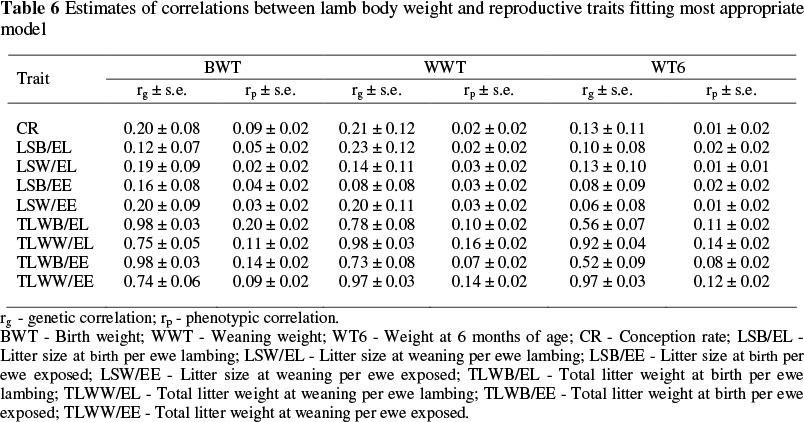
The estimates of correlations between lamb body weight and reproductive traits obtained from fitting the most appropriate model are presented in Table 6. Genetic correlation coefficients among lamb body weight and reproductive traits were positive and ranged from low (0.06) to very high (0.98). Birth weight (BWT) had a positive and low genetic correlation with CR, LSB/EL, LSW/EL, LSB/EE and LSW/EE and indicated that selection to improve BWT would result in low genetic response in conception rate and traits related with litter size at birth and weaning per ewe lambing and per ewe exposed. Similar to current estimates, Bromley et al. (2001) reported low to moderate genetic correlation between BWT and litter size at birth (-0.01 to 0.26) and between BWT and litter size at weaning (-0.37 to 0.01) for Columbia, Polypay, Rambouillet and Targhee sheep breeds. Hanford et al. (2002) reported a correlation of 0.10 between BWT and litter size at birth and 0.00 between BWT and litter size at weaning in a Columbia sheep breed. The estimates of genetic correlation between BWT and litter size at birth and weaning in the current study were similar to average estimates reported by Safari et al. (2005) but smaller than the averages reported by Fogarty (1995). Estimates of genetic correlation between BWT and TLBW/EL, TLWW/EL, TLBW/EE and TLWW/EE in the current study were positive and ranged from 0.74 to 0.98. Hence, selection based on BWT could lead to a high improvement in total litter weight at birth and weaning per ewe lambing and per ewe exposed. Bromley et al. (2001) reported -0.22 to 0.28 for the genetic correlation between BWT and total litter weight weaned in Columbia, Polypay, Rambouillet and Targhee sheep breeds, and unlike in the current study, these results indicate that selection to improve BWT would have little influence on genetic response in total litter weight weaned.
The estimates of genetic correlations between weaning weight (WWT) with CR, LSB/EL, LSW/EL, LSB/EE and LSW/EE obtained in the current study ranged from 0.08 to 0.23, while estimates of genetic correlations between WWT with TLBW/EL, TLWW/EL, TLBW/EE and TLWW/EE were positive and ranged from 0.73 to 0.98. Estimates of genetic correlation between WWT and reproductive traits in the literature are very variable. These estimates range from 0.07 to 0.40 for genetic correlation between WWT and litter size at birth (Al-Shorepy & Notter, 1996; Olivier et al., 2001; Hanford et al., 2002; Matika et al., 2003; Hanford et al., 2003; Van Wyk et al., 2003), from 0.15 to 0.43 for genetic correlation between WWT and litter size at weaning (Hanford et al., 2002; Hanford et al., 2003; Van Wyk et al., 2003), and from 0.10 to 0.75 for genetic correlation between WWT and total litter weight at birth and weaning (Ap-Dewi et al., 2002; Van Wyk et al., 2003; Safari et al., 2005). Estimates of genetic correlation between WWT with CR, LSB and LSW per ewe lambing and per ewe exposed obtained in this study were in the range of estimates in the literature. However, the positive and high estimates of genetic correlations between WWT with total litter weight at birth and weaning per ewe lambing and per ewe exposed in the current study were higher than values reported in the literature. These differences between results obtained in this study and the literature could be attributed to the rate of twinning in different breed of sheep. In the Lori-Bakhtiari sheep twinning rate is low. Therefore, total lamb weaning weight per ewe that lambed and per ewe exposed, are correlated to lamb body weight, unlike that in prolific breeds.
The estimates of genetic correlations between weight at six months of age (WT6) with CR, LSB/EL, LSW/EL, LSB/EE and LSW/EE obtained in the current study were low (0.06 to 0.13) but estimates of genetic correlations between WT6 with TLBW/EL, TLWW/EL, TLBW/EE and TLWW/EE were positive and high (0.52 to 0.97). These estimates indicate that similar to BWT and WWT, selection based on WT6 would have little influence on genetic response in CR, LSB and LSW per ewe lambing and per ewe exposed, but total litter weight at birth and weaning per ewe lambing and per ewe exposed increased due to correlated responses. The results of Safari et al. (2005) are in accordance with the results of the current study, in that estimates of genetic correlations between post-weaning weight with litter size were low to moderate and between post-weaning weight with total litter weight high.
The estimates of phenotypic correlations between lamb body weight at different ages and reproductive traits in this study were lower than the genetic correlations, and ranged from 0.01 to 0.20. These estimates are similar to those reported in the literature (Fogarty, 1995; Safari et al., 2005).
Conclusions
The heritability estimates obtained for lamb body weight and reproductive traits in the present investigation indicated that response to selection for traits of lamb body weight were higher than reproductive traits. The estimates of genetic correlations between lamb body weight traits and various reproductive traits were positive and ranged from low to high. Estimates of genetic correlations between weaning weight and lamb body weight at six months of age with reproductive traits, especially total litter weight weaned per ewe exposed as net reproductive (ewe productivity), were very high and positive, suggesting that selection based on increasing weaning weight or weight at six months of age may increase genetic merit in ewe productivity traits. Therefore, weaning weight or weight at six months of age could be considered as selection criteria to indirectly improve the reproductive traits (ewe productivity) in breeds of sheep such as Lori-Bakhtiari sheep with low twinning rates.
References
Al-Shorepy, S.A. & Notter, D.R., 1996. Genetic variation and covariation for ewe reproduction, lamb growth and lamb scrotal circumference in a fall-lambing sheep flock. J. Anim. Sci. 74, 1490-1498. [ Links ]
Ap-Dewi, I., Saatci, M. & Ulutas, Z., 2002. Genetic parameters of weights, ultrasonic muscle and fat depths, maternal effects and reproductive traits in Welsh Mountain sheep. Anim. Sci. 74, 399-408. [ Links ]
Boujenane, I., Kerfal, M. & Khallouk, M., 1991. Genetic and phenotypic parameters for litter traits of D'man ewes. Anim. Prod. 52, 127-132. [ Links ]
Bromley, C.M., Van Vleck, L.D. & Snowder, G.D., 2001. Genetic correlations for litter weight weaned with growth, prolificacy and wool traits in Columbia, Polypay, Rambouillet and Targhee sheep. J. Anim. Sci. 79, 339-346. [ Links ]
Ekiz, B., Özcan, M. & Yilmaz, A., 2005. Estimates of phenotypic and genetic parameters for ewe productivity traits of Turkish Merino (Karacabey Merino) sheep. Turk. J. Vet. Anim. Sci. 29, 557-564. [ Links ]
Fogarty, N.M., 1995. Genetic parameters for live weight, fat and muscle measurements, wool production and reproduction in sheep: A review. Anim. Breed. Abstr. 63, 101-143. [ Links ]
Gallivan, C., 1996. Breeding objectives and selection indexes for genetic improvement of Canadian sheep. Ph.D. thesis. University of Guelph. 174 pp. [ Links ]
Hanford, K.J., Van Vleck, L.D. & Snowder, G.D., 2002. Estimates of genetic parameters and genetic change for reproduction, weight, and wool characteristics of Columbia sheep. J. Anim. Sci. 80, 3086-3098. [ Links ]
Hanford, K.J., Van Vleck, L.D. & Snowder, G.D., 2003. Estimates of genetic parameters and genetic change for reproduction, weight, and wool characteristics of Targhee sheep. J. Anim. Sci. 81, 630-640. [ Links ]
Matika, O., Van Wyk, J.B., Erasmus, G.J. & Baker, R.L., 2003. Genetic parameter estimates in Sabi sheep. Livest. Prod. Sci. 79, 17-28. [ Links ]
Meyer, K., 2006. WOMBAT- A program for mixed model analyses by restricted maximum likelihood. User notes, Animal Genetics and Breeding Unit, Armidale. 55 pp. [ Links ]
Näsholm, A. & Danell, O., 1996. Genetic relationships of lamb weight, maternal ability and mature ewe weight in Swedish fine wool sheep. J. Anim. Sci. 74, 329-339. [ Links ]
Olivier, W.J., Snyman, M.A., Olivier, J.J., Van Wyk, J.B. & Erasmus, G.J., 2001. Direct and correlated responses to selection for total weight of lamb weaned in Merino sheep. S. Afr. J. Anim. Sci. 31, 115-121. [ Links ]
Rosati, A., Mousa, E., Van Vleck, L.D. & Young, L.D., 2002. Genetic parameters of reproductive traits in sheep. Small Rumin. Res. 43, 65-74. [ Links ]
Safari, E., Fogarty, N.M. & Gilmour, A.R., 2005. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest. Prod. Sci. 92, 271-289. [ Links ]
SAS, 1996. Statistical Analysis Systems user's guide (Release 6.11). SAS Institute Inc., Cary, North Carolina, USA. [ Links ]
Snyman, M.A., Erasmus, G.J., Van Wyk, J.B. & Olivier, J.J., 1995. Direct and maternal (co)variance and heritability estimates for body weight at different ages and fleece traits in Afrino sheep. Livest. Prod. Sci. 44, 229-235. [ Links ]
Snyman, M.A., Olivier, J.J., Erasmus, G.J. & Van Wyk, J.B., 1997. Genetic parameters for total weight of lamb weaned in Afrino and Merino sheep. Livest. Prod. Sci. 48, 111-116. [ Links ]
Snyman, M.A., Cloete, S.W.P. & Oliver, J.J., 1998. Genetic and phenotypic correlations of total weight of lamb weaned with body weight, clean fleece weight and mean fibre diameter in three South African Merino flocks. Livest. Prod. Sci. 55, 157-162. [ Links ]
Vatankhah, M., 2005. Defining a proper breeding scheme for Lori-Bakhtiari sheep in village system. Ph.D. thesis. University of Tehran, Iran. 207 pp. [ Links ]
Van Wyk, J.B., Fair, M.D. & Cloete, S.W.P., 2003. Revised models and genetic parameter estimates for production and reproduction traits in the Elsenburg Dormer sheep stud. S. Afr. J. Anim. Sci. 33, 213-222. [ Links ]
 Correspondence:
Correspondence:
E-mail: vatankhah_mah@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}