










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 n.2 Pretoria Feb. 2008
Sprout selection and performance of goats fed Acacia karroo coppices in the false thornveld of the Eastern Cape, South Africa
C.C. Nyamukanza; P.F. Scogings
Department of Agriculture, University of Zululand, Private Bag X1001, KwaDlangezwa 3886, South Africa
ABSTRACT
Acacia karroo Hayne is the dominant invading species in semi-arid savannas of South Africa and is an ecological threat of our modern era. This study investigated the preference and intake rates by goats when fed A. karroo coppice sprouts of different basal diameter sizes, viz. 3 mm, 4 mm and 5 mm. A study was also conducted to determine the digestibility and performance of goats when fed A. karroo coppices and commercial feed pellets. Nitrogen content varied among the sprout sizes. The larger sprouts had the highest nitrogen content, but the acid detergent fibre, neutral detergent fibre and hemicellulose did not differ among sprout sizes. There were no significant differences in preference of the different sprout sizes. Intake rate of the sprout sizes was significantly different; small sprouts were consumed at the highest intake rate compared to medium size sprouts, but not compared to the larger sprouts. Nguni goats had higher bite rates compared to Boer goats. However, intake rates of the Boer goats and the Nguni goats were not significantly different. Goats fed A. karroo had a higher average daily gain compared to the goats fed the pellets. Average daily gain differed between the two goat breeds. Apparent digestibility of dry matter and crude protein for A. karroo and pellets also differed. It is concluded that both Boer and Nguni goats have the potential to utilize smaller A. karroo sprouts. These animals can therefore potentially be used to control bush encroachment.
Keywords: Browse, bush encroachment, herbivory, intake rate, preference, digestibility
Introduction
Goats are important browsers in the Eastern Cape Province, which keeps nearly half of the goat population of South Africa (Dziba et al., 2003). Although goats are important for meat, milk and hide production (Devendra & Burns, 1993), they may be a major cause of habitat destruction if poorly managed (Scogings, 1998).
Goats have the ability to adapt to all climatic conditions and are important from an ecological perspective in controlling bush encroachment. Bush encroachment constitutes a major management problem in savannas (Mahanjana & Cronje, 2000; Ward, 2005). Persistent invasion of the grass sward by the bush component is caused by various factors, including low fire frequency, under-utilisation of the bush component, and overgrazing of the herbaceous component, which together tip competitive interactions in favour of encroachment by woody species (Smit et al., 1999). Goats can effectively control sprouts and regrowth following mechanical control (Du Toit, 1972; Strang, 1974; Trollope, 1974; Trollope et al., 1989).
Leguminous forages and fodder trees are an important source of feed for animals in tropical areas (Goromela et al., 1997; Aganga et al., 2000). The major limitation to the quality of these feeds is the presence of secondary plant compounds such as tannins, which can suppress feed intake and utilization by animals (Thi Mui et al., 2002). Goats are known for their ability to consume tannin-containing brush and have the ability to utilize forage and shrub species that contain tannins and phenolic compounds at levels that prohibit their selection as feeds by sheep and cattle (Aganga et al., 1998).
Acacia karroo Hayne (Fabaceae = Leguminosae) trees are abundant and able to grow in severe and dry conditions (Abdulrazak et al., 2001). It is important to know the preference of goat breeds for the different sizes of coppice sprouts in order to control the coppicing A. karroo trees, the main woody invader in the Eastern Cape. This information can be used to determine when, and with which breed, the sprouts can be effectively controlled. These inherent preferences partly reflect different morphologies and anatomies of these animal types, which influence their ability to prehend different plants or plant parts and to detoxify plant allelochemicals (Olson, 1999).
The objectives of this study were, firstly, to determine patterns of diet selection and intake rates by Nguni and Boer goats of A. karroo coppices of different sizes in a controlled experiment, and secondly, to evaluate the effect of A. karroo coppices on digestibility and growth of goats.
Materials and Methods
The study was conducted at the Fort Hare Research Farm near Alice, in the Eastern Cape Province of South Africa. The research site is at an altitude of 500 - 620 m above sea level. The region is predominantly savanna and consists of subtropical thicket vegetation dominated by deciduous woody shrubs shorter than 1.5 m although the woody layer can reach up to 5 m (Scogings et al., 1996). The vegetation is representative of Acocks (1975) 'False Thornveld of the Eastern Cape' and ranges from grassland to dense, clumped, medium-short, evergreen thicket (Dziba, 2000). Acacia karroo occurs in almost pure stands as an open to dense short tree savanna and is the dominant woody species. The vegetation type is thus variable, ranging from diverse grasslands with A. karroo invasion to dense thickets of A. karroo scrub and depauperated, overgrazed grasslands.
Annual rainfall is extremely variable both temporally and spatially and the mean annual rainfall at the research farm was 639 mm from 1970 to 1998, with a coefficient of variation of 22% (Dziba et al., 2003). Most (70%) of the rainfall occurs in summer, from October to April. In this period there is a bimodal distribution, with a major peak in March and a lesser one in October. The mean daily summer temperatures range from 18 - 21 °C and the mean daily winter temperatures range from 10 - 13 °C.
Sprout preference and intake rates of goats were determined using the dominant invading A. karroo. Goats used in this study included the Nguni and the Boer since they are farmed together in the Eastern Cape grass-bush savannas. Six non-lactating mature female Nguni goats aged between three and four years and with an average weight of 38.2 ± 3.5 kg were used in the study. The other six non-lactating mature female Boer goats were aged between three and five years and had an average weight of 53.6 ± 3.4 kg. The animals were dewormed and placed in individual pens where they were allowed a conditioning period to habituate them to the feed, environmental conditions and experimental procedure. In the first phase of habituation, animals were given two weeks to adapt to the pens and the feed, which was lucerne from the farm and 'ram, lamb and ewe' pellets from Epol (Pty) Ltd., Berlin, South Africa. The pellets contained, on a dry matter (DM) basis, 130 g protein/kg, 25 g fat/kg, 140 g crude fibre/kg, 15 g calcium/kg, 3 g phosphorus/kg, 10 g urea/kg, 5 g/kg of other non-protein nitrogen sources and 10 000 IU vitamin A/kg DM. In the second phase, the animals were given a two week adaptation period. Animals were run through the exact procedure of the experiment to identify any potential problems of handling and measurement.
On each morning of the experiment three different sizes of sprout material were measured and cut from the coppicing trees felled three months previously. The diameters were 3 mm, 4 mm and 5 mm because they were representative of what was available on the coppices at the time of the experiment. The sprouts were kept in water to reduce transpiration loss. Some sprouts were kept as controls to correct for weight loss due to transpiration. Ten samples of each size of sprout were placed into cooler boxes packed with ice blocks and then later freeze dried before chemical analysis.
In the experiment, each individual goat was transferred from its pen to an adjacent feeding pen where it was offered the three different sprout sizes. These sprouts were mounted 30 cm apart on a crescent-shaped artificial foraging board allowing the animal access to the feed material without necessarily moving its whole body (Dziba et al., 2003). The goats were deprived of food in the morning before the experiment to make sure they had an appetite. The experiment was run for 18 days. No sprouts of the same size appeared in one position more than twice per goat to control possible effects of position on the foraging board. A 6x6 Latin Square design was used to ensure that animals would have an equal opportunity to select from all options on offer.
The animals were allowed to consume the offered A. karroo sprouts for a maximum three minutes to ensure that foraging behaviour was not influenced by depletion of leaves on the branch (Green, 1984). If an animal prematurely stopped foraging before the three minutes was up, then 20 seconds after the animal stopped the trial was ended.
In each feeding bout (when a goat was offered all three sprout diameters simultaneously), the diameter of the sprout that was first bitten was recorded, and the number of bites, sprout mass before and after feeding, and the duration of the feeding bout were recorded. For each sprout diameter in each feeding bout, the ratio of the proportion of total ingested mass to the proportion of total available mass was calculated as a preference index (PI) (Chesson, 1983). If PI for a particular sprout size was less than 1 then that size was avoided and if PI was greater than 1 for a particular size then that size was preferred.
To calculate bite rate (bites/s), the number of bites counted during feeding and the time, in seconds (s), spent feeding were used. The difference between the initial branch mass and the residual branch mass (after correcting for transpiration loss) was divided by the number of bites to give bite size (g/bite). Intake rates (g/s) were estimated as the product of bite rate and bite size.
In the growth trial 12 non-lactating goats, which were used in the preference trial, were used in a completely randomized experiment replicated three times. Due to sickness one Nguni goat had to be removed from the study. The goats were drenched against internal parasites and dipped to control external parasites.
The goats were randomly placed in metabolism crates and randomly assigned to the dietary treatments, i.e., lucerne/A, karroo and lucerne/pellets. The goats were given an adaptation period of two weeks. The diets for each goat were calculated using values of metabolisable energy (ME) according to NRC (1981) recommendations for body weight (BW). The animals were fed DM at levels of approximately 3% of their body weight.
The animals were weighed before the trial, during the habituation period and every day during the first week of the trial. They were then weighed randomly during the course of the trial and then every day during the last week of the trial until the end of the trial. Feed samples were taken every day and pooled per animal over the collection period to be analysed. Refusals were also taken daily from each pen and pooled into weekly samples to be analysed. Refusal values were used to correct for the feed values consumed by the animal. The experimental period lasted a total of 31 days.
A study was also conducted to determine the digestibility of 4, karroo when fed to the two goat breeds. The study was incorporated into the performance study and ran concurrently with the performance study. The performance study was a 5-week experiment in which two digestibility trials of five days were incorporated. The animals were fed twice a day, in the morning and late in the afternoon with fresh 4, karroo coppices and lucerne and the control group was fed with pellets and lucerne. The animals were fed DM at levels of approximately 3% of their body weight. Water was accessible every time.
The metabolism crates allowed for the collection of faeces. Faeces were emptied daily in the morning and the total mass of the removed faeces for each animal was recorded. Ten per cent of the daily faeces voided was sampled and weighed before and after drying for DM determination. The feed, refusal and faecal samples were pooled per animal over the collection period and ground through a 1 mm screen and then stored in sealed plastic containers for analysis.
Sprout samples for the preference study and feed and faecal samples for the growth study were analysed for DM, crude protein (CP), neutral detergent fibre (NDF) and acid detergent fibre (ADF). Hemicellulose was calculated by difference between ADF and NDF. Nitrogen (N) was determined using the Kjeldahl method and CP was calculated as N x 6.25 (AOAC, 1990). Acid detergent fibre and NDF were analysed as free ash fractions by the sequential procedure of Goering & Van Soest (1970) using the ANKOM200/220 Fiber Analyser (ANKOM Corporation® Technology, Fairport, NY).
Analysis was done using the general linear model procedure of SPSS Version 12 (SPSS, 2003) statistical software and with SYSTAT 10 (SPSS, 2000). The data were checked for compliance with the assumptions of parametric statistical analysis. A one-factor ANOVA was done for the chemical data and a two-factor ANOVA was done for the preference and intake rate data. The LSD test was used to compare the means. A nested design was used for analysis of the performance data, to overcome pseudoreplication (Underwood, 1997), with diet nested within breed (Nguni goats and Boer goats).
Results
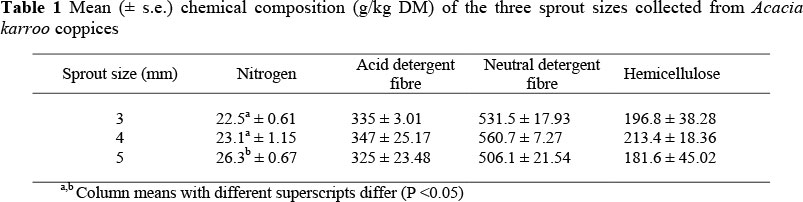
The chemical composition of the A. karroo sprouts is summarized in Table 1. Hemicellulose, NDF and ADF did not vary with sprout diameter (P >0.05), but N increased with an increase in sprout diameter (P <0.05) (Table 1). For the performance trial, the chemical composition for A. karroo was as follows: N = 23.7 ± 0.60 g/kg, NDF = 587.5 ± 8.93 g/kg, ADF = 399.1 ± 12.12 g/kg and hemicellulose = 188.4 ± 15.22 g/kg. For the pellets (according to the manufacturer): N = 20.8 g/kg, NDF = 200 g/kg and ADF = 58.0 g/kg.
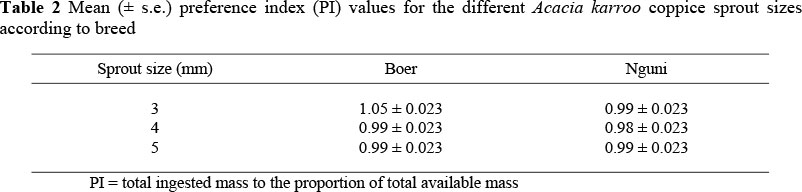
The interaction between breed and sprout size was not significant (P >0.05) in terms of PI. Neither the effect of breed nor sprout size had a significant effect on PI (Table 2).
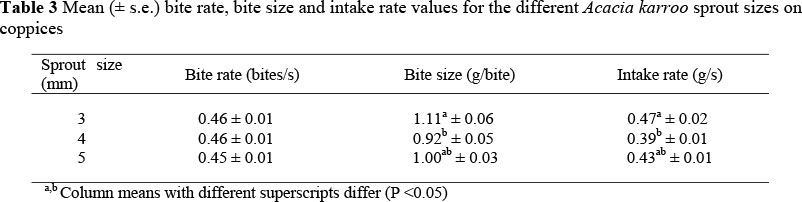
Goats ate the browse material on offer until the end of the trial period of three minutes. On average, 65 ± 0.5% (IMAGEMAQUI ± s.e., n = 1296) of each sprout was consumed, regardless of size. There were no significant differences in bite rates among the different sprout sizes (Table 3). However, Nguni goats had higher (P <0.05) bite rates (0.47 ± 0.01 bites/s) compared to the Boer goats (0.45 ± 0.01 bites/s). The interaction between breed and sprout size was not significant for bite rate.
Bite size differed among the different sprout sizes (P <0.05) (Table 3). Goats achieved the highest bite sizes from the 3 mm sprout size followed by the 5 mm and the 4 mm sprout sizes. Bite size was not, however, significantly different for the two goat breeds. The mean bite size for the goats was 1.01 ± 0.03 g/bite. The interaction of breed and sprout size was also not significant for bite size.
Intake rates for the goats differed among the different sizes of the sprouts (P <0.05). Intake rates were higher for the 3 mm sprouts than for the 4 mm sprouts (Table 3). There were no significant breed effects on overall intake rates and the size of the sprout most frequently bitten first (P >0.05). A total of 38% of the 5 mm sprouts, 35% of the 4 mm sprouts and 27% of the 3 mm sprouts were bitten first.
Goats fed A. karroo tended to have a higher average daily gain (ADG) compared to the goats fed the pellets (P = 0.061) (Table 4). The average daily gain of the two goat breeds did not differ significantly (P >0.05).
The apparent digestibility of DM for the pellets was 85.8 ± 3.64% and for A. karroo it was 63.4 ± 3.88% and the apparent digestibility of CP for the pellets was 88.2 ± 0.69% and for A. karroo it was 51.7 ± 0.59%. A significant difference in apparent DM digestibility of A. karroo and pellets (P <0.05) was observed, but not in apparent CP digestibility (P >0.05). There were no significant (P >0.05) differences in apparent digestibility of DM and hemicellulose for the two goat breeds when fed A. karroo (Table 5). However, breed effects were significant (P <0.05) in apparent digestibility of CP, NDF and ADF. These values were higher for the Boer goats than the Nguni goats (Table 5).
Discussion
It was noted that the larger sprouts (5 mm) had a higher N content than the smaller sprouts. The larger sprouts are faster growing than the smaller ones within a coppice and have a competitive advantage in acquiring nutrients. The larger sprouts had higher N compared to the smaller sprouts because of higher photosynthetic activity. This means higher chloroplast and related cofactors and enzymes such as rubisco which may account for 20 - 40% of a leaf s organic N (Loomis, 1997; Proe et al., 2002).
The study also showed no significant differences in the PI of all the sprout sizes. The goats did not prefer any particular sprout size. Boer goats were expected to prefer larger sprouts than the Nguni goats because of their body and mouth sizes. However, our results showed that there was no difference in the two breeds' preference for the sprout sizes. The range in sprout sizes was probably too small to detect differences in PI.
The bite sizes and intake rates for the three different sprout sizes were significantly different. Bite size was highest for the smallest 3 mm sprouts and subsequently intake rate was also high, compared to the other larger sprouts due to the differences in biomass density of each sprout size.The differences in bite size and intake rate are due to physical as well as structural properties of the vegetation (Illius et al., 1999).
The higher ADG of goats fed A. karroo coppices compared to those fed pellets was probably due to the high N content of A. karroo. The lower weight gains obtained in this study could be ascribed to the goats being older and the lower weight gains observed for the Nguni goats are due to the smaller size of the Nguni goat breed.
Nutritive value is determined by the chemical composition of the material, digestibility of nutrients and efficiency of utilization of the nutrients (Van Soest, 1994). Scogings & Macanda (2005) reported values of N content of A. karroo coppices which were similar to values obtained in this study. The NDF levels in this study, however, were much higher than those of A. karroo coppicing trees reported by Scogings & Macanda (2005), who reported concentrations of approximately 39%. The NDF fraction is correlated to forage intake by ruminants. The high levels of fibre in the feed could have had an effect on the low average daily gains of the animals reported in the present study due to lignification of the fibre component. An increase in ADF and NDF usually decreases digestibility, intake and animal performance (Van Soest, 1994). Danckwerts (1989) reported that the animal and the properties of the feed determine animal performance.
The apparent CP digestibility for the A. karroo coppices was similar to other browse species whose apparent digestibility of CP ranged from 50-57% (Salem et al., 2005). Aganga et al. (1998) reported DM digestibility of 55.9% for mature A. karroo which was lower than values reported for this study. The reduced CP digestibility values can be attributed to secondary compounds (Van Soest, 1994) and the stage of growth of the browse species.
Browse plants have the potential to provide enough nutrients to goats despite the presence of phenolic compounds since goats are able to utilize proteins from browse plants (Aganga & Monyatsiwa, 1999). Acacia karroo coppices are capable of supplying nutrients to goats and goats are able to utilize the nutrients as can be observed in the chemical composition of the A. karroo coppices and the LWG of the goats. Further research on the effect of A. karroo on the performance of goats is necessary if goats are to play an integral part in controlling the invading browse species, A. karroo.
Conclusion
Acacia karroo sprouts have good potential as a goat diet and both breeds have tended to utilise better the smaller sprouts. Continuous browsing by goats of such sprouts when young will reduce the amount of regrowth of coppices of A. karroo, which in turn can halt encroachment of the species in semi-arid areas of South Africa.
Acknowledgements
The research was supported by National Research Foundation grant GUN2050690. Iain Gordon (CSIRO) and Alan Duncan (Macaulay Institute) provided valuable advice on the experimental design. Iain Gordon, Luthando Dziba, Michael Undi, Thomas Papachristou, Ignatius Nsahlai, Wellington Ekaya and Craig Morris kindly provided useful comments.
References
Abdulrazak, S.A., Nyangaga, J. & Fujihara, T., 2001. Relative palatability to sheep of browse species, their in sacco degradability and in vitro gas production characteristics. Asian-Aust. J. Anim. Sci. 14, 1580-1584. [ Links ]
Acocks, J.P.H., 1975. Veld Types of South Africa, Mem. Bot. Surv. South Africa. 2nd ed. vol. 40. National Botanical Institute, Pretoria. [ Links ]
Aganga, A.A. & Monyatsiwa, C.B., 1999. Use of browses (Terminalia serecia, Combretum apiculatum or Euclea schimperi) as a supplement for growing Tswana goats. Tropic. Anim. Health Prod. 31, 295-305. [ Links ]
Aganga, A.A., Adogla-Bessa, T., Omphile, U.J. & Tshireletso, K., 2000. Significance of browses in the nutrition of Tswana goats. Arch. Zootec. 49, 469-480. [ Links ]
Aganga, A.A., Tsopito, C.M. & Adogla-Bessa, T., 1998. Feed potential of Acacia species to ruminants in Botswana. Arch. Zootec. 47, 659-668. [ Links ]
AOAC, 1990. Official Methods of Analysis (15th ed.). Association of Official Analytical Chemists, Inc., Arlington, V.A., USA. [ Links ]
Chesson, J., 1983. The estimation and analysis of preference and its relationship to foraging models. Ecology 64, 1297-1304 [ Links ]
Danckwerts, J.E., 1989. Animal performance. In: Veld Management in the Eastern Cape. Eds. Danckwerts, J.E. & Teague, W.R., Government Printer, Pretoria, South Africa. [ Links ]
Devendra, C. & Burns, M., 1993. Goat production in the tropics. Commonwealth Agric. Bureaux, London. [ Links ]
Du Toit, P.F., 1972. The goat in a bush-grass community. Proc. Grassld. Soc. Sth. Afr. 7, 44-50. [ Links ]
Dziba, L.E., 2000. Diet selection and foraging efficiency of boer goats and nguni goats in the False Thornveld of the Eastern Cape, South Africa. MSc (Agric) thesis, University of Fort Hare, Alice, South Africa. [ Links ]
Dziba, L.E., Scogings, P.F., Gordon, I.J. & Raats, J.G., 2003. Effects of season and breed on browse species intake rates and diet selection by goats in the False Thornveld of the Eastern Cape, South Africa. Small Rumin. Res. 47, 17-30. [ Links ]
Goering, H.K. & Van Soest, P.J., 1970. Forage fibre analysis. Agriculture Handbook No. 379. Agriculture Research Station, Washington, D.C., USA. [ Links ]
Goromela, E.H., Ledin, I. & Uden, P., 1997. Indigenous browse leaves as supplements to dual purpose goats in central Tanzania. Livest. Prod. Sci. 47, 245-252. [ Links ]
Green, R.F., 1984. Foraging rules for optimal foragers. Am. Nat. 123, 30-40. [ Links ]
Illius, A.W., Gordon, I.J., Elston, D.A. & Milne, J.D., 1999. Diet selection in goats: a test of intake-rate maximization. Ecology 88, 1008-1018. [ Links ]
Loomis, R.S., 1997. On the utility of nitrogen in leaves. Proc. Natl. Acad. Sci. USA.94, 13378-13379. [ Links ]
Mahanjana, A.M. & Cronje, P.B., 2000. Factors affecting goat production in a communal farming system in the Eastern Cape region of South Africa. S. Afr. J. Anim. Sci. 30, 149-154. [ Links ]
NRC., 1981. Nutrient Requirements of Goats: Angora, Dairy, and Meat Goats in Temperate and Tropical Countries. National Academy Press, Washington, D.C., USA. [ Links ]
Olson, B.E., 1999. Manipulating diet selection to control weeds. In: Grazing Behaviour of Livestock and Wildlife. Eds. Launchbaugh, K.L., Sanders, K.D. & Mosley, J.C., Idaho Forest, Wildlife and Range Exp. Sta. Bull. 70, Univ. Idaho, Idaho. pp. 36-44. [ Links ]
Proe, M.F.,Griffiths, J.H. & Craig, J., 2002. Effects of spacing, species and coppicing on leaf area, light interception and photosynthesis in short rotation forestry. Biomass and Bioenergy 23, 315-326. [ Links ]
Salem, A.Z.M., Salem, M.Z.M., El-Adawy, M.M. & Robinson, P.H., 2006. Nutritive evaluations of some browse tree foliages during the dry season: Secondary compounds, feed intake and in vivo digestibility in sheep and goats. Anim. Feed Sci. Technol. 127, 251- 267. [ Links ]
SPSS, 2000. SYSTAT 10. SPSS Inc., Chicago. [ Links ]
SPSS, 2003. SPSS for Windows version 12.0.1, Chicago, Illinois, USA. [ Links ]
Scogings, P.F., 1998. Resistance of six savanna woody plant species to browsing by goats in the Eastern [ Links ]
Cape province of South Africa. Unpublished PhD thesis. University of Fort Hare, Alice, South Africa. [ Links ]
Scogings, P.F. & Macanda, M., 2005. Acacia karroo responses to early dormant season defoliation and debarking by goats in a semi-arid subtropical savanna. Plant Ecology 179, 193-206. [ Links ]
Scogings, P.F., Trollope, W.S.W. & O'Connor, T.G., 1996. The impact of browsing and burning on savanna vegetation. In: Rangelands in a Sustainable Biosphere, vol. 1. Ed. West, N.E., Society of Range Management, Denver. pp. 498-499. [ Links ]
Smit, G.N., Aucamp, A. & Richter, C.G.F., 1999. Bush encroachment: an approach to understanding and managing the problem. In: Veld Management in Southern Africa. Ed. Tainton, N.M., University of Natal Press, Pietermaritzburg, South Africa. [ Links ]
Strang, R.M., 1974. Some man-made changes in successional trends on the Rhodesian highveld. J. Appl. Ecol. 11, 249-263. [ Links ]
Thi Mui, N., Ledin, I., Uden, P. & Van Binh, D., 2002. Nitrogen balance in goats fed Flemingia (Flemingia macrophylla) and Jackfruit (Artocapus heterophyllus) foliage based diets and effect of a daily supplementation of polyethylene glycol (PEG) on intake and digestion in goats. Asian-Aust. J. Anim. Sci. 15, 699-707. [ Links ]
Trollope, W.S.W., 1974. Role of fire in preventing bush encroachment in the Eastern Cape. Proc. Grassld. Soc. Sth. Afr. 9, 67-72. [ Links ]
Trollope, W.S.W., Hobson, F.O., Danckwerts, J.E. & Van Niekerk, J.P., 1989. Encroachment of undesirable species. In: Veld Management in the Eastern Cape. Eds. Danckwerts, J.E. & Teague, W.R., Government Printer, Pretoria, South Africa. [ Links ]
Underwood, A.J., 1997. Experiments in Ecology. Cambridge University Press, Cambridge, UK. [ Links ]
Van Soest, P.J., 1994. Nutritional Ecology of the Ruminant, 2nd ed. Comstock, Cornell Univ. Press, Ithaca, N.Y., USA. [ Links ]
Ward, D., 2005. Do we understand the causes of bush encroachment in African savannas? Afr. J. Range Forage Sci. 22, 101-105. [ Links ]
 Correspondence:
Correspondence:
E-mail: cnyamuka@pan.uzulu.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}