










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.38 no.1 Pretoria Jan. 2008
Across flock genetic parameter estimation for yearling body weight and fleece traits in the South African Dohne Merino population
J.B. van WykI, # ; J.W. SwanepoelI; S.W.P. CloeteII, III; J.J. OlivierIV; G.J. DelportI, V
IDepartment of Animal, Wildlife and Grassland Science, University of the Free State, PO Box 339, Bloemfontein 9300, South Africa
IIDepartment of Animal Sciences, Stellenbosch University, Private Bag X1, Matieland 7602, South Africa
IIIInstitute for Animal Production: Elsenburg, Private Bag X1, Elsenburg 7607, South Africa
IVARC Livestock Business Division: Private Bag X5013, Stellenbosch 7599, South Africa
VAnimal Breeding Consultant, PO Box 440, Middelburg 5900, South Africa
ABSTRACT
Accurate genetic parameter estimates are needed upon which to perform multiple-trait across flock breed analyses. Genetic parameters for yearling body weight (BW), clean fleece weight (CFW) and mean fibre diameter (MFD) were estimated using records of 107 389 individuals (the progeny of 1 530 sires and 45 178 dams) collected between 1992 and 2004 in the South African Dohne Merino population. Fixed effects included in the model were flock-year-season-sex-management group (1 594 classes), type of birth (singles, multiples), age of dam (2 - 7+ years) and age at measurement, fitted as a linear covariate (385 ± 12 days). Six different single-trait animal models were fitted, where different combinations of the following random effects were fitted: direct additive, the sire-flock interaction, the sire-flock-year-season interaction (SFYS), the dam genetic effect, the direct-maternal correlation and the dam permanent environmental effect. These analyses were followed by a three-trait analysis structured according to the log likelihood ratios obtained for the single-trait analysis. This analysis allowed the calculation of relevant correlations among traits together with their respective standard errors. Direct heritability estimates from the three-trait analysis were 0.17 for BW, 0.19 for CFW and 0.45 for MFD. Maternal heritability estimates were 0.01 for BW and 0.006 for CFW, with corresponding dam permanent environmental ratios of respectively 0.03 and 0.02. The genetic correlation between animal effects amounted to 0.48 for BW. Derived proportions of the total phenotypic variance due to SFYS were 0.02, 0.02 and 0.02, respectively. Genetic correlations of BW with CFW and MFD were 0.11 and 0.13, respectively, and of CFW with MFD 0.16. It was concluded that the inclusion of some form of a genotype by environmental interaction as part of the national evaluation is essential, although it controlled only a modest portion of the overall phenotypic variation.
Keywords: Direct heritability, maternal effects, genotype x environment interaction, correlations.
Introduction
The Dohne Merino breed is a synthetic breed established, ca. 1939 in the Eastern Cape Sourveld region of South Africa, from a cross between the Merino and the then German Merino (presently known as the SA Mutton Merino). The original intention was to develop a hardy and versatile genotype that would adapt well to semi-intensive farming practices in this region (Kotzé, 1951). This new breed was also to cope with the seasonal undersupply of nutrients during winter. It was named after the Dohne Research Institute where it was developed. The Dohne Merino has since proved itself as an extremely adaptable dual-purpose breed with easy-care properties and an ability to thrive under widely divergent conditions. This has resulted in a sustained growth in numbers, and expansion to other areas in South Africa. Increasing numbers of Dohne Merinos have also been exported recently to other major sheep producing countries, where it is believed that they would have an important role to play in the further development of the sheep industry in those countries.
Since introduction of a performance recording scheme for wool-sheep in South Africa ca. 1980 (Erasmus & Hofmeyr, 1984), recording has been compulsory for all registered Dohne Merino breeders. A sire referencing scheme for the Dohne Merino stud industry was introduced in 1992. Initially, genetic evaluation was based on a single-trait sire model using parameters estimated by Delport & Botha (1994). Since 1996 similar single-trait animal models (Delport, unpublished) were used for analyzing data from the sire referencing scheme and flocks of interested breeders. An evaluation of all recorded animals was introduced in 2005, again using single-trait animal models. Our aim was to construct models and estimate necessary parameters for yearling body weight (BW), clean fleece weight (CFW) and mean fibre diameter (MFD) of South African Dohne Merino sheep with the goal of assisting the Dohne Merino Breeders' Association in embarking on an across-flock multiple-trait national genetic evaluation.
Materials and Methods
The data consisted of records from purebred Dohne Merino sheep from 97 studs situated throughout South Africa. Modelling of systematic effects can be problematic when environmental conditions fluctuate from year to year and from region to region. Nevertheless the breeders applied the same set of record keeping guidelines set by the Dohne Merino Breeders' Association. In short, the management guidelines for record keeping prescribed by the Dohne Merino Breeders' association are as follows (McMaster, 2000): Weights are recorded on-farm by breeders. At the age of 12/14 months animals are weighed and shorn for performance testing. After shearing, the fleece of each individual animal is weighed and a fleece sample is sent to one of the wool testing facilities in South Africa for analysis. Since 2005 the ARC Small Stock Improvement Scheme used the raw data to do performance testing and calculate within-flock indices and individual breeding values. This information is returned to the breeder to complement selection decisions. The complete data set, after editing, comprised 107 389 records, the progeny of 1 530 sires and 45 178 dams, collected between 1992 and 2004. Contemporary groups were defined as flock-year-season-sex-management group (FYSSM). Records of sires with fewer than 25 progeny in the data set and flocks without genetic linkage (at least two sires) were excluded from the data. Contemporary groups with less than 10 records were also excluded.
Production traits analyzed included yearling (actual age ± s.d. = 385 ± 12 days) BW, CFW and MFD. Fixed effects included in the model were FYSSM (1 594 classes), type of birth (singles, multiples), age of dam (2 - 7+ years) and age at measurement fitted as a linear covariate. All these effects were significant (P <0.01) and retained in subsequent analyses. ASREML (Gilmour et al., 2002) was used to estimate (co)variance components for each trait. Random terms were added to the fixed effect model, resulting in different single-trait animal models with various combinations of random effects for each trait. These included direct and maternal genetic effects and the covariance between them, permanent environmental effects due to the dam and either sire-flock or sire-flock-year-season effects. The different single-trait models that were fitted are summarized in Table 2. The order in which random effects were entered into this hierarchy has been decided a priori and models were compared in the following order (Table 2): 1) 1 vs. 2-testing c2sf; 2) 1 vs. 3 - testing c2sfys and when c2sfys had a higher contribution; 3) 3 vs. 4 - testing m2; 4) 4 vs. 5 - testing ram; 4) 5 vs. 6 - testing c2pe.
Tests of significance for each random effect were performed using the log likelihood ratio test (LRT) after including each random effect (excluding the residual) to the fixed effects model. An effect was considered significant when its inclusion in the model caused a significant increase in the log likelihood ratio. A chi square distribution for α = 0.05 and one degree of freedom was used as the critical test statistic (3.841). When -2 times the difference between log likelihoods was greater than the critical value the inclusion of the effect was considered significant. After the most appropriate single-trait models for the different traits were determined, estimates obtained from the single-trait analyses were used as starting values in the three-trait analyses. The most appropriate multiple-trait analysis incorporated the "best" models from the single trait analyses, and allowed the calculation of relevant correlations among traits together with their respective standard errors.
Results and Discussion
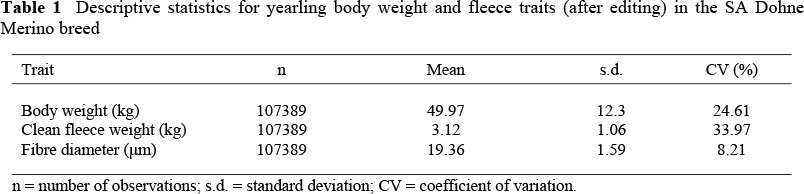
Coefficients of variation for the quantitative production traits were high, ranging from 24.6% for live weight to 34.0% for greasy fleece weight. Coefficients of variation for weight traits (Table 1) were considerably higher than the weighted means calculated by (Safari et al., 2005) mostly from experimental flocks. Coefficients of variation below 10% are commonly found for fibre diameter while more variation is expected in the other production traits. Corresponding coefficients of variation in the seven Australian Merino research resource flocks used for across flock analyses ranged from 18% to 31% for CFW, from 16.7% to 22.2% for yearling BW and from 8.7 % to 11.1% for MFD (Safari et al, 2007a).
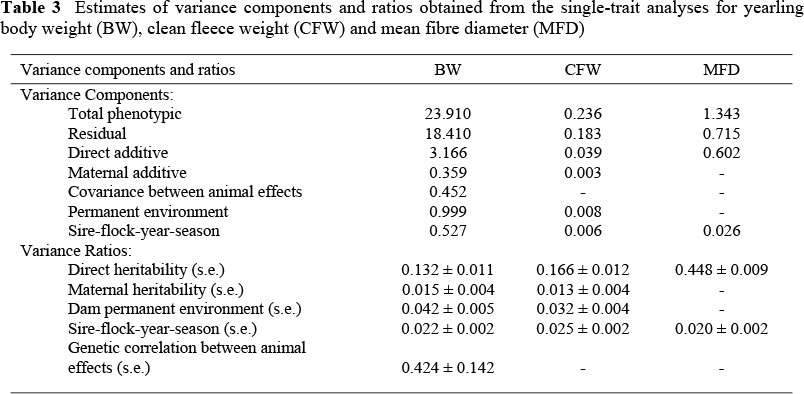
According to the LRT from single-trait analyses (Table 2) the most appropriate model for BW included both direct and maternal additive genetic effects, the covariance between animal genetic effects, permanent environmental effects due to the dam, and sire-flock-year-season (SFYS) as random factors. The appropriate model for CFW was similar except for the exclusion of the covariance between animal effects. The model with SFYS as the only additional random term to the direct genetic effect was found to be superior for MFD.
The derived direct heritability (h2) estimates were considerably lower than means derived for wool sheep, from the literature by Safari et al. (2005). Respective h2 estimates reported in the latter review amounted to 0.30 for BW, 0.28 for CFW and 0.59 for MFD. No apparent reason for this discrepancy can be provided, except that many of the references cited in the review of Safari et al. (2005) reported on data from experimental flocks where more control arguably existed over managerial regimes as compared to industry data. However, it needs to be stated that the present h2 estimate for MFD accorded very well with estimates of 0.44 - 0.47 derived from a breed analysis on Merino sheep in South Africa (Olivier et al., 1998).
Expressed relative to the total phenotypic variation, maternal genetic effects (m2) in the single-trait analyses amounted to 1.5% for BW and to 1.3% for CFW (Table 3). These estimates are consistent with the lower ranges of comparable estimates in the literature. Previous estimates of m2 on Merino types ranged from 0.01 to 0.15 for body weight and from 0.03 to 0.17 for clean fleece weight (see summary from the literature by Cloete et al., 2002). Corresponding estimates for the dam permanent environment (c2pe) were 4.5% for BW and 3.2% for CFW. It is noteworthy that the estimates of m2 were lower at ~0.015 than that of c2pe in the current study, and it is tempting to consider these effects as unimportant. However, maternal genetic effects were generally more important than maternal permanent environmental effects in the review of Safari et al. (2005). Estimates for c2 generally were negligible or non-existent. The same general result was reported in the subsequent paper of Safari et al. (2007b) where data from a number of Australian Merino resource flocks were pooled in a single analysis involving > 110 000 records. The fact that the present data were obtained over a relatively short period of time may have contributed to the contrasting result in the present study, making it difficult to partition the maternal variance into its genetic and permanent environmental components. The correlation between animal effects for BW was similar in sign to that derived by Safari et al. (2005) from literature values. However, the magnitude of the present estimate (0.42) was about double that of the derived mean (0.18). Safari et al. (2007b) accordingly reported direct-maternal correlations of 0.07 for yearling live weight and 0.25 for hogget live weight. Sire-flock-year-season was fitted as an additional random effect and contributed between 2.0% and 2.5% to total phenotypic variance in the respective traits. Comparable results were reported by Olivier et al. (1998) for CFW and MFD in a breed analysis of South African Merinos, although the SFYS variance ratio was substantially higher at 0.07 to 0.08 for BW. Results reported for CFW (0.03 - 0.04) and MFD (0.02) were more in line with those derived in the present study.
The most appropriate three-trait model consisted of the following effects that were indicated by the LRT in the single trait analyses (see Table 2 for definition of variance ratios):
BW= h2 + c2sfys + m2 + ram + c2pe
CFW= h2 + c2sfys + m2 + c2pe
MFD=h2 + c2sfys
Parameter estimates fitting these models in the three trait analyses are presented in Table 4. The most striking feature of these results is the fact that estimates of h2 for BW and CFW were somewhat higher than those obtained from single-trait analyses, while the estimate for MFD remained constant. The other variance ratios (m2, c2pe and c2sfys) were generally somewhat lower for BW and CFW than in the single-trait analyses. Some of the variation in these effects was thus seemingly repartitioned towards the direct, additive variance component in the three-trait analysis.
The direct heritability estimate from the three-trait analysis was calculated at 0.17 for yearling BW (Table 4). This estimate is within the range reported by Safari et al. (2005) but lower than the weighted mean of 0.31 and 0.41 for wool and dual-purpose breeds, respectively. The only two previously animal model estimates for BW of Dohne Merino sheep were 0.32 for the SA Dohne Merino breed (Delport, unpublished) and 0.24 ± 0.06 for the Kromme Rhee Dohne Merino nucleus flock (Cloete et al., 1998). Both previous estimates were higher than the present value. This is probably due to different and more sophisticated models used in the latter and/or a lower level of control of on-farm performance recording implemented in the Dohne Merino industry.
The weighted mean heritability in the case of CFW was 0.28 in wool breeds (Safari et al., 2005). At 0.19, the estimate of the present study is somewhat lower than this weighted mean. Estimates of respectively 0.25 and 0.35 derived for Dohne Merinos by Delport (unpublished) in the Dohne Merino breed and by Cloete et al. (1998) for the Kromme Rhee Dohne Merino nucleus flock were also somewhat higher than the present estimate.
The heritability estimate for MFD (0.45) was lower than the weighted means from various studies (Fogarty, 1995; Safari et al., 2005). However, it was somewhat higher than the value of 0.39 previously estimated in the Dohne Merino breed (Delport, unpublished). The animal model estimate reported by Cloete et al. (1998) on the Kromme Rhee Dohne Merino nucleus flock amounted to 0.43, which accorded very well with the present estimate.
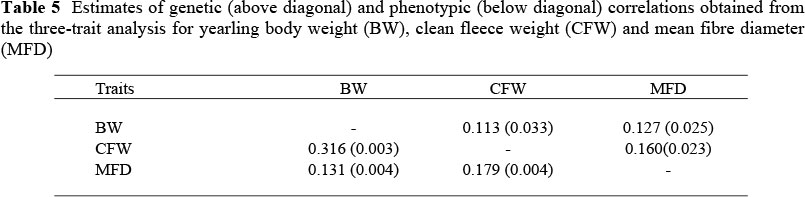
Genetic correlations (Table 5) suggested a relatively low (but positive) genetic correlation between BW and CFW. The comparable value derived from literature values by Safari et al. (2005) was about twice as high at 0.24. The earlier review by Fogarty (1995) reported variable weighted mean genetic correlations of BW with MFD (-0.11 for yearling weight and 0.10 for hogget weight), which were more consistent with the present estimate. Genetic correlations of BW with MFD (0.13) and of CFW with MFD (0.16) were slightly higher, but still below the mean values of respectively 0.20 and 0.28 derived by Safari et al. (2005) from a comprehensive list of literature sources. The phenotypic correlations were comparable with the genetic correlations pertaining to MFD, but the phenotypic correlation of BW with CFW was substantially higher than the genetic correlation.
Conclusions
Estimates of phenotypic and genetic parameters constantly need to be updated and refined to facilitate accurate multiple-trait breeding value predictions in all breeds. The present study is the first published multiple-trait evaluation of the South African Dohne Merino breed. A significant sire-flock-year-season effect for all traits suggests that inclusion of some form of a genotype by environmental interaction be included in the national evaluation for Dohne Merinos. Significant maternal effects (genetic and permanent environmental) also imply that the inclusion of maternal ability in the national evaluation should be considered. The comparatively low genetic correlations among body weight, clean fleece weight and fibre diameter may be an indication that wool quality as well as wool quantity might not be compromised as a correlated response to selection for meat production traits in the Dohne Merino.
Acknowledgements
Funding for this study was partly provided by the National Research Foundation. The authors wish to thank the Dohne Merino Breeders' Association for kind permission to use the data.
References
Cloete, S.W.P., Greeff, J.C. & Lewer, R.P., 2002. Direct and maternal (co)variances for hogget live weight and fleece traits in Western Australian Merino sheep. Aust. J. Agric. Res. 53, 271-279. [ Links ]
Cloete, S.W.P., Scholtz, A.J. & Aucamp, B.B., 1998. Environmental effects, heritability estimates and genetic trends in a Western Cape Dohne Merino nucleus flock. S. Afr. J. Anim. Sci. 28, 185-590. [ Links ]
Delport, G.P. & Botha, A.F., 1994. Reducing fibre diameter variation and coarse edge in Dohne Merinos -Preliminary results. Proc. 5th World Congr. Gen. Appl. Livest. Prod. 19, 417-420. [ Links ]
Erasmus, G.J. & Hofmeyr, J.H., 1984. The development of performance recording programmes for woolled sheep in South Africa. Proc. 2nd World Conf. Sheep Beef Cattle Breed. pp. 252-256. [ Links ]
Fogarty, N.M., 1995. Genetic parameters for live weight, fat and muscle measurements, wool production and reproduction in sheep: a review. Anim. Breed. Abstr. 63, 101-143. [ Links ]
Gilmour, A.R., Gogel, B.J., Cullis, B.R., Welham, S.J. & Thompson, R., 2002. ASREML User Guide Release 1.0. VSN International Ltd, Hemel Hempstead, HP11es, UK. [ Links ]
Kotzé, J.J.J., 1951. The development of a mutton-wooled sheep for the Sour-grassveld area. Farming in South Africa. Reprint no. 26, pp. 110-113. [ Links ]
McMaster, J.C., 2000. Prestasietoetsing van Kleinvee. Agricultural Research Council (ARC). pp. 1-21. [ Links ]
Olivier, J.J., Yurgens, Y., Mostert, B.E. & Erasmus, G.J., 1998. Across-stud evaluation of Merinos in South Africa. Proc. 6th World Cong. Gen. Appl. Livest. Prod. 24, 11-14. [ Links ]
Safari, E., Fogarty, N.M. & Gilmour, A.R., 2005. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest. Prod. Sci. 92, 271-289. [ Links ]
Safari, E., Fogarty, N.M., Gilmour, A.R., Atkins, K.D., Mortimer, S.I., Swan, A.A., Brien, F.D., Greeff, J.C. & van der Werf, J.H.J., 2007a. Across population genetic parameters for wool, growth, and reproduction traits in Australian Merino sheep. 1. Data structure and non-genetic effects. Aust. J. Agric. Res. 58, 169-175. [ Links ]
Safari, E., Fogarty, N.M., Gilmour, A.R., Atkins, K.D., Mortimer, S.I., Swan, A.A., Brien, F.D., Greeff, J.C. & Van der Werf, J.H.J., 2007b. Across population genetic parameters for wool, growth, and reproduction traits in Australian Merino sheep. 1. Estimates of heritability and variance components. Aust. J. Agric. Res. 58, 177-184. [ Links ]
# Corresponding author. E-mail: vanwykjb.sci@ufs.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}