










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.35 no.1 Pretoria 2005
Simulated natural lighting and constant 14-hour photoperiods for broiler breeders during the rearing period, and interactions of lighting with body weight
P.D. Lewis#; M. Ciacciariello; M. Nonis; R.M. Gous
Animal and Poultry Science, University of KwaZulu-Natal, Pietermaritzburg 3209, South Africa
ABSTRACT
Spring-hatched broiler breeders housed in non-lightproof facilities are frequently maintained on long daylengths to prevent precocity, but controlled-feeding and delayed dissipation of photorefractoriness retard sexual maturation. Trials were conducted to determine the appropriate growth for birds maintained on 14-h photoperiods, and to compare responses with naturally increasing and decreasing daylengths. Birds maintained on 14-h photoperiods matured about 5 d earlier and produced 2 more eggs to 60 weeks for each 10-d advance in age at 2.1 kg. However, despite requiring 0.2 kg less feed to reach 2.1 kg, accelerating growth resulted in 0.5 kg more feed being consumed to sexual maturity. The accelerated-growth birds produced more eggs that were too small for hatching and more double-yolked eggs during the early weeks of production. The data suggest that profitability is maximised when constant-14-h birds reach 2.1 kg at about 18-19 weeks. There were no significant differences in maturity between conventionally-grown birds given increasing or decreasing daylengths or those held on 14-h photoperiods, though there was a tendency for the constant 14-h birds to have later maturity, to produce 6-7 fewer eggs to 60 weeks, and to consume more feed per egg, resulting in a lower economic margin than naturally lighted birds. The wisdom of rearing Spring-hatched broiler breeders on long photoperiods to avoid precocity is therefore questionable. Birds allowed faster growth, generally matured about three weeks earlier than conventionally-grown birds, but matured five weeks earlier when given an increasing lighting regimen that started from 10 h rather than 11 h.
Keywords: Broiler breeders, body weight, natural light, constant photoperiods, sexual maturity, egg production
Introduction
Ad libitum-fed egg-type hybrids reared on increasing photoperiods mature precociously, produce an increased number of small eggs, and have a propensity for prolapsus. Although extensive and open-sided housing are not normally recommended for rearing replacement pullets, in some parts of the world their use is unavoidable. In such situations, supplementation of natural lighting with artificial illumination to create a daylength equal to the longest natural daylength expected during the rearing phase has long been the standard recommendation to avoid the adverse consequences of precocity (Morris, 1968). However, whilst this may be sound advice for ad libitum-fed egg-type hybrids, it might not necessarily be the correct strategy for rearing control-fed broiler breeders. Gous & Cherry (2004) and Lewis et al. (2003; 2004) produced evidence that broiler breeding stock still exhibit photorefractoriness, and that rearing on long photoperiods delays its dissipation and retards sexual maturity. In conventionally grown broiler breeders, rearing on constant long days retards sexual maturity by three to four weeks, compared with the normal practice of rearing on short days and photostimulating at about 20 weeks, which increases the cost of rearing birds to point of lay, reduces egg numbers to a given age and increases egg weight.
The principal down-side of rearing egg-type pullets on increasing daylengths is the reduction in egg weight that emanates from advanced maturity. However, this should not be a problem for broiler breeders, and, indeed, might even be a benefit. Data from Lewis et al. (2003) showed that pullets grown to typically recommended broiler breeder body-weight targets do not start to respond to an increment in photoperiod until after 10 weeks of age, and, as a flock, are not fully photosensitive until about 18 weeks. It follows, therefore, that the risks involved in rearing broiler breeders on naturally increasing daylengths might be minimal, especially in comparison with egg-type pullets. Throughout the history of the broiler breeder industry, age at sexual maturity (ASM) has been reported to be closely linked to prepubertal growth rate (e.g. Isaacks et al., 1960; Lee et al., 1971; Pym & Dillon, 1974; Blair et al., 1976; Leeson & Summers, 1983; Yu et al., 1992; Yuan et al., 1994; Nøddegaard et al., 2000), and so a relaxation of feed control might be a way of cancelling out the retardational influences of rearing on constant long days.
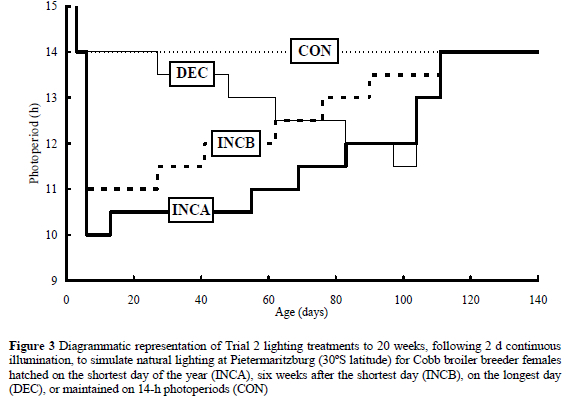
This paper describes two trials that were conducted to assess the effects of growth rate when rearing broiler breeders on constant 14-h photoperiods, and to quantify the risks of rearing on naturally changing daylengths. In the first trial, two opposing hypotheses were tested, (1) that accelerated growth would counter the delaying effect of rearing on constant long days, and (2) that slower growth would ensure that the body weight at a given rate of egg production would match that recommended for normally reared broiler breeders. In the second trial, birds were reared on accelerated and standard growth curves, and, within each group, were given a lighting regimen that simulated natural daylengths for birds hatched on the shortest day (INCA) or six weeks after the shortest day (INCB), simulated natural daylengths for birds hatched on the longest day of the year (DEC), or maintained on 14 h to 20 weeks (CON).
Materials and Methods
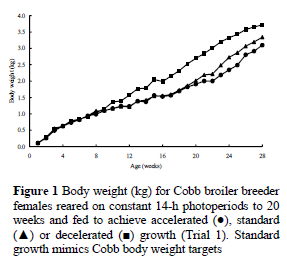
In Trial 1, 1500 Cobb broiler breeder females were randomly allocated at 1 d of age to three light-proof floor-rearing rooms (10.5 birds/m2), with each room divided into two pens by a wire partition, and each pen holding birds that were fed to achieve one of the three body weight profiles in Figure 1 (nominally, accelerated, standard and decelerated growth). All rooms were given an initial 2 d of continuous illumination before being permanently changed to a 14-h photoperiod (07.00-21.00), the longest daylength at a 30° latitude (e.g. Pietermaritzburg), through to the end of the trial at 57 weeks. Illumination was from four 60 W incandescent lamps in each room, and mean illuminance at a height of 30 cm was 26 ± 2.8 lux. At 15 weeks, a total of 990 birds were transferred to 18 floor pens in an open-sided house (six pens of 55 birds for each growth treatment, stocked at 5.2 birds/m2). Natural light was supplemented with artificial illumination from a single 15 W compact fluorescent lamp located centrally in each pen to maintain a 14-h photoperiod.
All birds were fed a 210 g CP/kg, 12.4 MJ AME/kg crumbled diet ad libitum until transfer to controlled-feeding and a 175 g CP/kg, 11.9 MJ AME/kg pelleted diet scattered on the litter floor from three weeks. At six weeks, the diet was changed to a 140 g CP/kg, 11.5 MJ AME/kg, and finally to a 145 g CP/kg, 11.5 MJ AME/kg layer diet from 5 eggs/100 bird.d (%) egg production onwards. Daily feed allocation was progressively increased to a peak of 170 g/bird/day between 30 and 32 weeks, depending on rate of lay, before being reduced by 5 g/bird for each subsequent 5% fall in rate of lay.
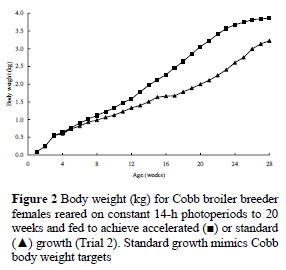
The birds in Trial 2 were housed in the same rearing and laying facilities as those in the first trial, except that they were placed in eight rather than three rooms (4 birds/m2) during the rearing phase, and there were two growth curves, designated accelerated or standard (Figure 2), with all rooms containing a pen of each. Two rooms were allocated to each of the four lighting schedules shown in Figure 3 (30-min changes were dictated by time clock design), and in the rooms that simulated natural lighting, 30 min of dim light (1.2 ± 0.18 lx) were given before and after the main photoperiod to mimic morning and evening civil twilight at 30° latitude. All groups were transferred to 16-h photoperiods when moved to the adult facilities at 20 weeks. Dietary specifications and feeding schedules were similar to Trial 1, with the exception that the daily feed allocation was maximised at 165 g/bird at 30 weeks.
Egg numbers were recorded daily in both trials, and eggs bulk-weighed to pen daily in the first trial and three times per week in Trial 2. Sexual maturity was defined as the second of two days in which the total number of eggs laid in a pen first equalled or exceeded the mean number of birds in that pen over the two days. Data from the first trial were subjected to a 1-way ANOVA using a model from Statistix, version 7 (Analytical Software, 2000), and data from the second trial were subjected to a 2-way ANOVA, with lighting and growth (the lighting x growth interaction was not significant at P < 0.05 for any parameter, and was dropped from the model) as variables.
Results
All data for Trial 1 are presented in Table 1. Accelerating growth, which required a 40% increase in feed allocation to 22 weeks, significantly advanced ASM by 12 d, and slowing growth, which saved about 10% feed to 22 weeks, significantly delayed ASM by 9 d. The accelerated birds produced 11 more eggs to 60 weeks than the decelerated birds (P < 0.05), with the standard group intermediate between, but not significantly different from, the other two groups. However, there was less than three eggs difference between the best (decelerated, 121.9) and worst (accelerated, 119.1) growth-treatments when egg numbers for the 25 weeks immediately following ASM were compared (P = 0.481). Mean egg weights for the three growth groups were not significantly different from each other. However, a regression of mean egg weights for the 18 individual pens on ASM showed that it increased by 0.3 g for each 10-d delay in ASM (P = 0.022, Slope SE = 0.01, SD = 0.56), and the three treatment egg-weight means were in the order of their ASM. Notwithstanding that the overall production of double-yolked eggs was minimal, there were no significant differences between treatments, with means of 0.98, 0.93 and 0.65 for the accelerated, conventional and decelerated growth groups, respectively. With significant differences in egg numbers, but similar egg weights, it is obvious that the total egg output for the groups would mirror the differences in egg numbers, with the accelerated growth producing 670 g more egg mass (P = 0.005) than the decelerated group, with standard birds again intermediate, but not significantly different. Despite the accelerated-growth group maturing 12 d earlier than the standard group, it weighed 200 g significantly heavier at, and consumed 2.7 kg more feed to, sexual maturity. Conversely, the decelerated-growth group weighed 140 g less at, and consumed 500 g less feed to, ASM, despite maturing 9 d later. The higher feed intake of the accelerated-growth group during the rearing period resulted in them requiring 14 g more feed (includes feed consumed in the rearing period) to produce an egg, and 0.2 g significantly more feed to produce 1 g of egg than either of the other two groups. There were no significant differences in body weight at 57 weeks of age.
n Trial 2, ASM for the accelerated-growth birds on the INCB, DEC and CON lighting treatments was 22-27 d earlier, resulting in the production of six to eight more eggs to 60 weeks, than for normally-grown birds, but for the INCA birds, faster growth resulted in a 34-d advance in ASM and the production of 19 more eggs (Table 2). However, when the effects of ASM were removed, there was no significant difference between the growth treatments for the number of eggs laid in the ensuing 29 weeks (accelerated growth: 134.3, standard growth: 136.9). Whereas, across all lighting treatments the mean egg weight for normally grown birds was 1.2 g heavier than for accelerated-growth birds (P = 0.014), the difference was 2.6 g for INCA birds (Table 2). The accelerated groups laid significantly more double-yolked eggs (1.39/bird) than the conventionally-grown groups (0.53/bird), and laid more eggs weighing < 50 g during the first 4 weeks of production (Figure 4). Nevertheless, superior egg numbers for the accelerated groups resulted in them yielding 0.5 kg more egg mass output to 60 weeks (P = 0.134) than the conventionally-grown (Table 2). The accelerated-growth birds used 0.9 kg less feed to reach 2.1 kg (at 103 d) than the normally grown birds (at 145 d), but consumed 1.8 kg more feed to ASM. The extra feed consumed to ASM resulted in the accelerated-growth birds being 0.25 to 0.50 kg heavier at sexual maturity than the conventionally-grown birds on the same lighting regimen (P < 0.001). The differences in feed intake to ASM automatically resulted in significantly higher total intakes to 60 weeks for the accelerated-growth birds, and the need for 20 g more feed to produce an egg and 0.4 g more per 1 g of egg (P < 0.05) than the conventionally-grown groups. Their body weight on depletion at 60 weeks was also significantly heavier than for the conventionally grown birds (Table 3).
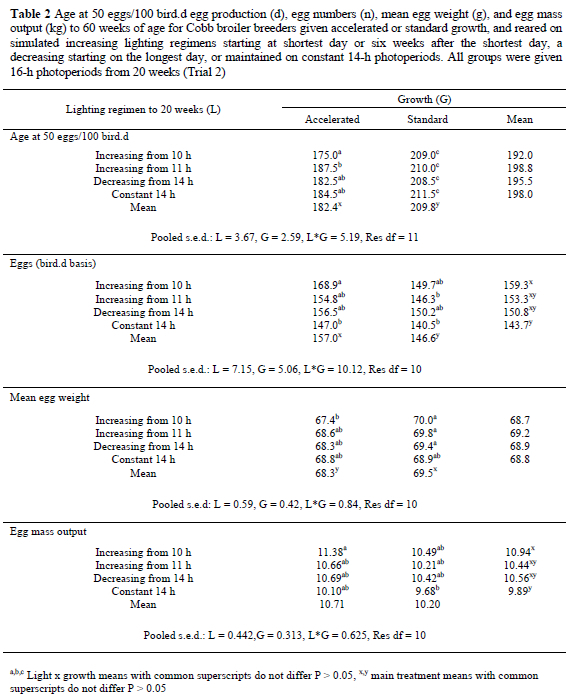
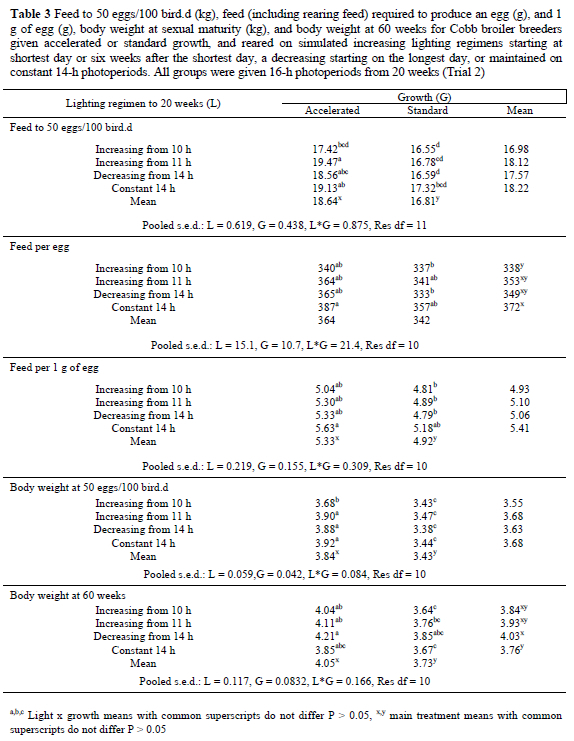
Lighting did not significantly affect ASM, but, within each growth-group, birds maintained on 14-h photoperiods produced between seven and 16 fewer eggs, and 0.5 to 1.0 kg less egg mass to 60 weeks than the other lighting regimens, with both significantly different for INCA (Table 2). There were no significant effects on mean egg weight or the production of double-yolked eggs. Even though lighting did not significantly affect cumulative feed intake to sexual maturity, the poorer egg production resulted in the birds maintained on 14-h photoperiods using 19 to 34 g more feed to produce an egg, and 0.3 to 0.5 g more feed to produce 1 g of egg than the other birds (Table 3). Body weight at sexual maturity was similar for all lighting groups, but the CON birds gained only 80 g during the laying cycle, compared to 400 g for the DEC birds, to become significantly lighter at 60 weeks. The INCA (+ 290 g) and INCB (+ 250 g) groups were intermediate between, but not significantly different from, the DEC and CON groups (Table 3).
There were no indications in either trial that mortality was affected by accelerating growth or by any of the lighting treatments.
Discussion
Data from both trials confirmed the earlier findings of Gous & Cherry (2004), Payne (1975) and Lewis et al. (2003; 2004) that rearing on constant long photoperiods to 20 weeks delays ASM compared with conventional broiler breeder lighting (Figure 4). However, it is clear that the extent to which ASM is retarded depends very much on growth rate during the rearing period. Taking the time to reach a body weight of 2.1 kg as an arbitrary measure of growth, meta-analyses of data from these two trials and laying data from the trial partially reported by Lewis et al. (2004), with significant differences of Trial 1 and Lewis et al. (2004) from Trial 2 removed by least squares analysis, were conducted to determine the effect of growth rate on ASM and other performance traits.
Sexual maturity for birds maintained on constant 14-h photoperiods occurred about 5 d earlier for each 10-d advance in age at 2.1 kg (Figure 5), with the regression described by the equation:
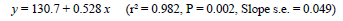
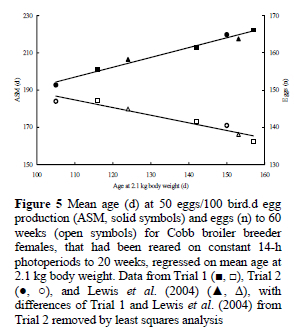
where y = ASM (d), and x = age at 2.1 kg body weight (d). This means that if birds are maintained on 14-h days during the rearing period, a 2.1-kg body weight needs to be reached about five weeks ahead of that recommended by the primary breeding company if 50% egg production is still to occur on the breeder's target of 26 weeks. The 0.53 rate of advance in ASM (d) achieved by accelerating growth (d) in these trials agrees remarkably well with the 0.55 figure reported by Leeson & Summers (1983) for birds reared on 8 h and transferred to 14-h daylengths at 2.1 kg. However, it is much larger than the 0.12 d/d observed by Yuan et al. (1994) for birds grown on various growth curves and photostimulated at the same age though at different body weights. Whereas the agreement of the current findings with Leeson & Summers (photostimulation at same weight) supports the suggestion of a threshold body weight and/or fat pool for the onset of lay in broiler breeders hypothesised by Bornstein et al. (1986), the contrast with the findings of Yuan et al. (1994), where body weight at photostimulation varied, shows that the body weight at photostimulation interacts with the age at which the light increment is given to influence ASM.
Although accelerating growth might resolve the problem of delayed ASM for birds maintained on long days during the rearing period, there is a price to be paid for using this technique. Notwithstanding that accelerating growth by five weeks reduced the feed required to reach 2.1 kg body weight by about 0.8 kg, the amount of feed consumed to ASM increased by almost 2 kg despite the 18-d earlier maturity (Figure 6). The regressions of cumulative feed intake to a 2.1-kg body weight and ASM are described by the equations:
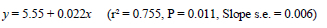
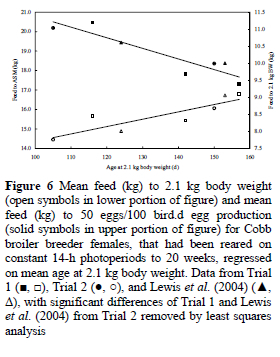
where y = feed to 2.1 kg BW (kg), and x = age at 2.1 kg body weight (d), and
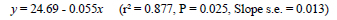
where y = feed to ASM (kg), and x = age at 2.1 kg body weight (d).
The lesser quantity of feed needed to reach 2.1 kg body weight when birds are given increased daily allocations is in agreement with the findings of Leeson & Summers (1983) and unpublished data from other work conducted at the University of KwaZulu-Natal.
The benefit of accelerating growth rate on eggs numbers to 60 weeks in the current trials was described by the equation (Figure 4):
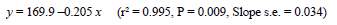
where y = eggs produced to 60 weeks (calculated on a bird.d basis), and x = age at 2.1 kg body weight (d). However, analyses of the total number eggs laid in a given period from ASM (174 d in Trial 1, and 202 d in Trial 2) showed that there were no significant differences between treatments in either trial, indicating that the differences in egg numbers within each trial were principally a consequence of the effect that the treatments had on ASM (Figure 5). This also concurs with other unpublished work conducted at KwaZulu-Natal. Although the above equations predict that a 35-d acceleration in growth to 2.1 kg body weight will result in an 18-d earlier ASM and about seven more eggs being laid to 60 weeks, the production of eggs averaging less than 50 g during the first few weeks of lay (Figure 7), and an increased production of double-yolked eggs (an extra 0.2 for each 10-d advance in ASM, P = 0.002), decreased the proportion of eggs suitable for hatching and partly negated the increase in total egg production.
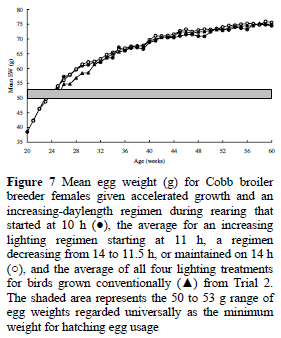
As a general rule, the value of a broiler breeder hatching egg is similar to the cost of 1 kg of feed, and so it is suggested that a useful indicator of gross margin might be the difference between the total feed consumed (kg) and the number of hatching-egg produced. This index conveniently provides an estimate of gross margin without the need for financial inputs, and can be universally applied. A regression of gross margin index on the age at 2.1 kg body weight for data from the current two trials and the experiment reported by Lewis et al. (2004) revealed a curvilinear response, and showed that, to maximise gross margin in birds reared on constant stimulatory photoperiods, 2.1 kg needs to be reached between 18 and 19 weeks (Figure 8). The regression was described by the equation:
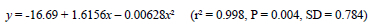
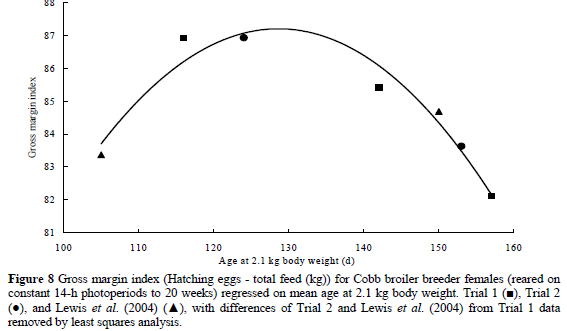
where y = gross margin index (hatching egg numbers (n) - total feed (kg), and x = age at 2.1 kg body weight (d). However, the optimal age to achieve a 2.1-kg body weight might be different when birds are given a conventional broiler breeder lighting programme, where photorefractoriness will be more quickly dissipated by the short days, and ASM advanced by photostimulation at about 20 weeks.
The most likely explanation for the 34-d difference in ASM between accelerated and conventionally grown birds exposed to the INCA regimen, but only 22-d difference when for the INCB birds (Table 3), is a difference in the rate at which photorefractoriness was dissipated. Shorter, less stimulatory daylengths and increased feed allocations both allow broiler breeders to become photoresponsive more rapidly than birds exposed to longer photoperiods or those given stricter feed control (Lewis et al., 2004). The difference between the two INCB groups suggests that the more generous feed allocation effected a 22-d advance in ASM, and the initial eight weeks of minimally stimulatory photoperiods (< 11-h) experienced by the INCA birds, in contrast with the INCB birds that had no exposure to < 11 h daylengths, accounted for a further 12 d of advanced maturity. Fuller et al. (1969) reported similar findings for White Plymouth Rock pullets, though without the comments on the dissipation of photorefractoriness. When reared on increasing daylengths (and so exposed initially to minimally stimulatory daylengths that would have facilitated photorefractoriness dissipation), birds fed a broiler starter diet ad libitum to six weeks matured 6.2 weeks earlier than others given a diet containing 33% less energy, but ASM was only 3.3 weeks earlier when birds were reared on decreasing daylengths (and would have experienced delayed photorefractoriness dissipation because of the initial exposure to stimulatory daylengths).
Although ASM for the DEC birds might have been expected to be delayed in comparison with those given constant or increasing daylengths, such a retardational effect would have been countered by the greater photostimulation experienced by these birds at 15 weeks when they were transferred from 11.5 to 13 h (Figure 3). These findings are in agreement with those of Brake et al. (1989) and Idris & Robbins (1994). However, similar ASM for birds exposed to naturally increasing and decreasing lighting treatments, be they simulated or actual, contrasts with a 14-d difference in ASM observed for broiler breeders hatched on the shortest or longest day and kept in non-lightproof houses in Spain at 42°N (Vandenberghe et al., 1999), and a 17-d difference reported by Lopez & Leeson (1992) for broiler breeders kept in open housing at between 20 and 25°N in Mexico. In these two reports, where birds were located at higher and lower latitudes than Pietermaritzburg, the data were for commercial flocks that had actually been hatched at different times of the year, and so exposed, not only to different daylength patterns, but also to different light intensity and temperature conditions, both of which are known to influence sexual maturation (Lopez & Leeson, 1992). In the current trials, only the lighting regimen was different, and this might explain the disagreement.
Although accelerating growth for birds maintained on 14-h photoperiods (2.1 kg at 116 vs. 142 d in Trial 1, 103 vs. 145 d in Trial 2) advanced ASM, enhanced egg numbers and increased total feed intake, and retarding growth (2.1 kg at 157 d in Trial 2) delayed ASM, reduced egg numbers, and reduced feed, intake neither strategy resulted in an improvement in gross margin (Figure 8). In the faster-growing birds, the superior egg production was insufficient to compensate for the extra feed that they required to achieve the accelerated growth, resulting in an inferior conversion of feed to egg - a finding in agreement with McDaniel (1983), and the feed saving achieved by slowing growth was insufficient to cancel the reduction in egg numbers that resulted from the delay in ASM.
The similar ASM, egg numbers, mean egg weights, and feed conversion efficiencies for the increasing- and the decreasing-daylength regimens grown conventionally in Trial 2, but reduced egg numbers and inferior feed utilisation for the birds maintained on 14 h to 20 weeks, indicate that the practice of rearing broiler breeders on constant long days to avoid the effects of precocity in open-sided or non-lightproof housing is both unnecessary and incorrect. Indeed, application of the gross index margin formula showed that maintaining birds on 14 h would result in at least a 6-unit reduction compared with any of the simulated natural-lighting regimens. However, the policy of rearing non-controlled environment broiler breeders on a constant daylength might still be well-founded for birds kept at low latitudes, where the longest anticipated natural daylength is only mildly stimulatory, or when other seasonally changing environmental factors, such as light intensity and temperature, adversely affect sexual maturation.
Acknowledgements
We wish to acknowledge the supply of day-old chickens and generous financial contributions from Cobb-Vantress, Siloam Springs, USA.
References
Analytical Software, 2000. StatistixVersion 7, Tallahassee, FL 32317, USA. [ Links ]
Blair, R., MacCowan, M.M. & Bolton, W., 1976. Effects of food regulation during the growing and laying stages on the productivity of broiler breeders. Br. Poult. Sci. 17, 215-223. [ Links ]
Bornstein, S., Plavnik, I. & Lev, Y., 1984. Body weight and/or fatness as potential determinants of the onset of egg production in broiler breeder hens. Br. Poult. Sci. 25, 323-341. [ Links ]
Brake, J., Garlich, J.D. & Baughman, G.R., 1989. Effects of lighting program during the growing period and dietary fat during the laying period on broiler breeder performance. Poult. Sci. 68, 1185-1192. [ Links ]
Fuller, H.L., Potter, D.K. & Kirkland, W., 1969. Effect of delayed maturity and carcass fat on reproductive performance of broiler breeder pullets. Poult. Sci. 48, 801-809. [ Links ]
Gous, R.M. & Cherry, P., 2004. The effects of body weight at, and lighting regimen and growth curve to, 20 weeks on laying performance in broiler breeders. Br. Poult. Sci. 45, 445-452. [ Links ]
Idris, A.A. & Robbins, K.R., 1994. Light and feed management of broiler breeders reared under short versus natural day length. Poult. Sci. 73, 603-609. [ Links ]
Isaaks, R.E., Reid, B.L., Davies, R.E., Quisenberry, J.H. & Couch, J.R., 1960. Restricted feeding of broiler type replacement stock. Poult. Sci. 39, 339-346. [ Links ]
Lee, P.J.W., Gulliver, A.L. & Morris, T.R., 1971. Restricted feeding of broiler breeder pullets during the rearing period and its effect on productivity and breeding. Br. Poult. Sci. 12, 499-510. [ Links ]
Leeson, S. & Summers, J.D., 1983. Consequence of increased feed allowance for growing broiler breeder pullets as a means of stimulating early maturity. Poult. Sci. 62, 6-11. [ Links ]
Lewis, P.D., Backhouse, D. & Gous, R.M., 2004. Constant photoperiods and sexual maturity in broiler breeder pullets. Br. Poult. Sci. 45, 557-560. [ Links ]
Lewis, P.D., Ciacciariello, M. & Gous, R.M., 2003. Photorefractoriness in broiler breeders: Sexual maturity and egg production evidence. Br. Poult. Sci. 44, 634-642. [ Links ]
Lopez, G. & Leeson, S., 1992. Manage broiler breeder pullets to reduce seasonal variations. Poult. Digest. September, 31-35. [ Links ]
McDaniel, G.R., 1983. Factors affecting broiler performance. 5. Effects of preproduction feeding regimens on reproductive performance. Poult. Sci. 62, 1949-1953. [ Links ]
Morris, T.R., 1968. Light requirements of the fowl. In: Environmental control in poultry production. Ed. T.C. Carter. Oliver & Boyd, Edinburgh. pp. 15-39. [ Links ]
Nøddegaard, F., Talbot, R.T. & Sharp, P.J., 2000. Effect of delayed step-up lighting on plasma luteinizing hormone and reproductive function in broiler breeders. Poult. Sci. 79, 778-783. [ Links ]
Payne, C.G., 1975. Day-length during rearing and the subsequent egg production of meat-strain pullets. Br. Poult. Sci. 16, 559-563. [ Links ]
Pym, R.A.E. & Dillon, J.F., 1974. Restricted food intake and reproductive performance of broiler breeder pullets. Br. Poult. Sci. 15, 245-259. [ Links ]
Vandenberghe, N., Ceular, A.L. & Moreno, A., 1999. Use of hatch date for broiler breeder production planning. Poult. Sci. 78, 501-504. [ Links ]
Yu, M.W., Robinson, F.E. & Robblee, A.R., 1992. Effect of feed allowance during rearing and breeding on female broiler breeders. 1. Growth and carcass characteristics. Poult. Sci. 71, 1739-1749. [ Links ]
Yuan, T., Lien, R.J. & McDaniel, G.R., 1994. Effects of increased rearing period body weights and early photostimulation on broiler breeder egg production. Poult. Sci. 73, 792-800. [ Links ]
# Corresponding author. E-mail: peter.lewis@dsl.pipex.com

