










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.34 no.5 Pretoria 2004
Protein supplementation, body condition and ovarian activity in goats -Pre-ovulatory serum profile of insulin
C.A. Meza-HerreraI, III, #; J.M. Sánchez S.I; J.G Chávez-PerchesII; H. SalinasIII; M. MelladoIV
IUnidad Regional Universitária de Zonas Aridas, Universidad Autônoma Chapingo, Mexico
IIRadiodiagnóstico y Ultrasonografía, Mexico
IIIInstituto Nacional de Investigaciones Forestales, Agricolas y Pecuarias, Mexico
IVUniversidad Autonoma Agraria Antonio Narro, Mexico
ABSTRACT
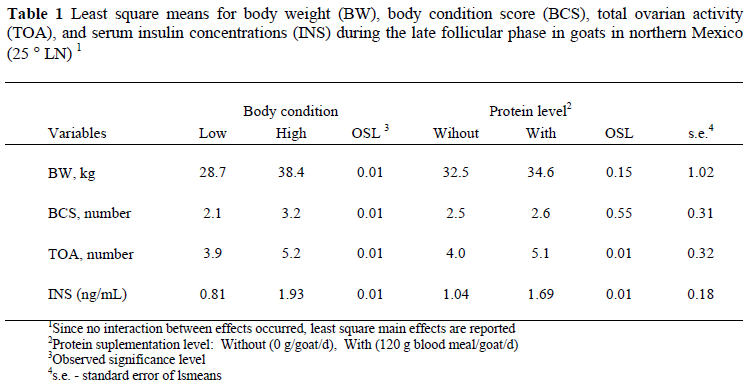
This study evaluated the effect of level of by-pass protein supplementation (PSL) and body condition (BC) on ovarian activity and serum insulin (INS) concentrations on does. Does (n = 32, 19 mo.) with either a low BC (LBC = 2.1 ± 0.3; n = 16; 28.8 ± 0.72 kg body weight, BW), or a high score (HBC = 3.2 ± 0.3; n = 16, 35.1 ± 0.72 kg body weight) received one of two levels of protein supplementation: Without supplemental protein (NPROT) or 120 g protein as blood meal/d (PROT) during a 40-d pre-ovulatory period. Once oestrous was synchronized, blood samples were collected during the mid-follicular phase at 60 min intervals during a 6-h period, to quantify serum INS concentration. During the late luteal phase (postovulation), total ovarian activity (TOA) was evaluated by transrectal ultrasonographic scanning, counting the total number of follicles (FT) and corpus lutea (CLT). While the HBC group had significantly more CLT than the LBC (2.81 ± 0.20 vs. 1.87 ± 0.20, respectively), no differences occurred between BC groups with respect to FT (2.43 ± 0.25 vs. 2.18 ± 0.25). While PSL affected CL and FT, favouring the PROT-goats (2.62 ± 0.21 vs. 2.06 ± 0.21, and 2.68 ± 0.25 vs. 1.92 ± 0.17, respectively), the HBC goats had significantly higher serum INS levels than the LBC group (1.92 ± 0.17 vs. 0.81 ± 0.17 ng/mL, respectively). The protein supplemented goats also displayed an increased serum INS concentration compared to the non-supplemented goats. A positive correlation was observed between serum INS concentrations and CLT (r = 0.46) and FT (r = 0.38) concentrations. Both the static effect (live weight body condition) and the dynamic effect (by-pass protein supplementation) of nutrition promoted a metabolic status characterized by high insulin levels and an increased ovarian activity.
Keywords: Goats, by-pass protein, body condition, ovarian activity, insulin
Introduction
While there is a paucity of data regarding the effect of protein supplementation on goats in different body conditions, a knowledge of nutritional and reproductive strategies can help to maximize overall production efficiency. Changes in metabolic fuel availability and associated endocrine status are important determinants in signalling the metabolic status to the reproductive axis. In fact, some metabolic hormones such as insulin could act as the signalling link between the metabolic status and reproductive function. Nutritional deficiencies or shortcoming on dietary protein are related in humans to low serum insulin levels and, consequently, hypoglycaemia (Picarel-Blanchot et al., 1995). In fact, animals in a negative energy balance are characterized by hypoinsulinaemia (Whitaker et al., 1993). In rats, insulin stimulates GnRH release, maintains normoglycaemia and provokes follicle-stimulating hormone (FSH) secretion and the pulsatility of the luteinizing hormone (LH), as well as luteal progesterone secretion (Arias et al., 1992). Nutritionally induced changes in the serum insulin level are strongly related to the IGF-1 and IGF-2 plasma concentrations, with increases in IGF-1, as well as to a concurrent increase in the steroidogenic ability of ovarian follicles (Spicer & Echternkamp, 1993). The aim of this study was to evaluate the effect of by-pass protein supplementation and body condition (BC) on ovarian activity and on serum insulin (INS) concentrations as well as their possible relationship in goats.
Materials and Methods
The study was carried out at the Southern Goat Research Unit of the Regional University Unit for Arid Lands-Chapingo Autonomous University, Bermejillo, Durango, Mexico. The unit is located at 103° 36' WL, 25° 53' NL, at 1, 117 meters above sea level. Annual averages for temperature and precipitation are 20 °C,and 242 mm, respectively. After a 40-d adaptation period, crossbred goats (Saanen-Alpine x Criollo, n = 32, 19 mo) were weighed at weekly intervals (BW, kg) and body conditions scored (BCS, 1 = very thin, 5 = very fat (Russel et al., 1969). The 16 heaviest goats received 1 kg lucerne hay (146 g CP/kg) per goat. The lighter 16 goats received 1 kg lucerne hay per goat plus 100 g of crushed maize, which was increased weekly up to 200 g at six weeks. Thereafter, the goats in the poor condition (LC, 28.4 ± 0.78 kg) and good condition (HC, 35.6 ± 0.78) groups received a basal diet of lucerne hay to supply 70% and 100% of their nutritional requirements, respectively. Both BW and BCS between groups differed (P < 0.01) at the end of the adaptation period.
Thereafter, the goats were randomly assigned, within body condition, to one of two bypass protein supplementation levels (PSL), viz. without protein (NPROT, 0 g/goat/d) or with protein (PROT, 120 g blood meal/goat/d) during 40-d pre-ovulation. The experimental design consisted of a 2 x 2 factorial arrangement of treatments, with two body condition levels (LC & HC) and two bypass protein supplementation levels (NPROT & PROT). The goats received a basal diet of lucerne hay (2.0% BW, 146 g CP/kg feed), mineral salts, water and shades during the whole experimental period (40 d). They were kept under natural photoperiod conditions prevailing in August and September at this latitude.
Oestrus was synchronized with two PGF2a injections, two days apart. A total of 16 goats was randomly selected for blood collection. Blood was collected via jugular venipuncture into 10 mL sterile vacutainer tubes (Corvac, Sherwood Medical St. Louis, MO) at 60 min intervals during a 6 h period. A total of seven samples per goat, 28 samples per treatment and 112 blood samples across the experiment was collected. Blood was allowed to clot for 30 min at room temperature and then centrifuged at 1500 g for 20 min. Serum was then decanted into polypropylene vials and stored at -20 °C. The INS concentration in the serum was determined by RIA with an intra-assay CV of 6.2% (Hoefler & Halford, 1987). During the late luteal post-ovulation phase, total ovarian activity (TOA) was evaluated through transrectal ultrasonographic scanning, using a 7.5 Mhz linear transducer (Toshiba Medical System, Ltd, Crawley, UK). The considered ovarian variables included the total number of follicles (TF), the total number of corpus lutea (CL) as well as total ovarian activity (TOA = TF + CL).
The variables BW, BC, and TOA were evaluated by ANOVA having a completely random design with a 2 x 2 factorial arrangement of treatments (Snedecor & Cochran, 1967). The serum insulin profile was analysed by split-plot ANOVA for repeated measures on animals across time (Gill & Hafs, 1971). In the case of significant treatment differences, comparisons between least square means were performed using the PDIFF option of the PROC GLM-SAS. Correlation analyses were conducted when appropriate, using the Pearson's Product Moment Test. All the analyses were done according to the procedures of SAS (Littlel et al., 1991).
Results and Discussion
While BC affected (P < 0.05) CLT in favour of the HBC compared to the LBC group (2.81 ± 0.20 vs. 1.87 ± 0.20, respectively), there were no differences in FT between the different BC groups (2.43 ± 0.25 vs. 2.18 ± 0.25, respectively). The number of CL and FT were both higher in the PROT goats than in the control (2.62 ± 0.21 vs. 2.06 ± 0.21, and 2.68 ± 0.25 vs. 1.93 ± 0.25, respectively). The HBC goats had the highest serum INS concentrations (1.9 ± 0.2 vs. 0.8 ± 0.2 ng/mL, respectively), and the supplemented goats recorded higher serum INS concentrations compared to the non-supplemented goats (Table 1). A positive correlation was observed between serum INS levels and CLT counts (r = 0.46; P = 0.06) and FT (r = 0.38; P = 0.10).
In this study, serum INS levels were affected by both BCS and the PSL. According to Peters & Mayer (1993) protein supplementation acts as a potent stimulator of INS secretion. Muturi et al. (2002) evaluated the effect of low and high levels of by-pass protein on plasma glucose, IGF-1 and INS concentrations. While no differences were observed on plasma IGF-1 levels, glucose concentrations differed (P < 0.05) at 60, 90 and 120 min. post-feeding, and high protein supplementation affected (P < 0.05) the plasma insulin concentrations. Animals in a negative energy balance displayed high blood levels of GH and NEFA's and low concentrations of IGF-1, insulin and glucose (Whitaker et al., 1993). Serum insulin concentration during mid-lactation in animals in a positive energy balance was close to 2.5 ng/mL, while in a negative energy balance during early lactation it was around 0.5 ng/mL. Therefore, glucose was shown to positively affect ovarian function, highlighting the role that insulin could play in that process as the main regulator of glucose levels and, in turn, in the process of ovarian follicular growth (Williams et al., 1997).
Almeida et al. (2001) reported increased ovulation rates in nutritionally restricted females who received exogenous insulin through promoting increases in the pulsatility of E2 and LH, diminishing follicular atresia, increasing the number of follicles of the pre-ovulatory pool and augmenting ovulation rate. In the long term, nutrition level determines both body weight and body condition, while in the short-term it is related to an increased level of nutrients at intracellular level, stimulating the release of gonadotropic hormones or acting directly in the ovary by increasing its metabolism and function (Cox et al., 1987).
Conclusions
Both the static (body weight, body condition) and the dynamic (by-pass protein supplementation) effects of nutrition in 19-mo goats generated a metabolic status characterized by high levels of serum insulin which in turn were positively correlated with increased ovarian activity. Results suggest that high serum insulin levels might have prevented follicular atresia and enhanced ovarian activity in both the good body conditioned and the by-pass protein supplemented goats, possibly in a route independent of increases in GnRH-gonadotropins. The better understanding of the interactions between nutrition and reproduction will contribute to enhance reproductive efficiency of breeding goat flocks through the development of optimal nutritional management strategies.
References
Almeida, F.R.C.L., Mao, J., Novack, S., Cosgrove, J.R. & Foxcroft, G.R., 2001. Effects of different patterns of feed restriction and insulin treatment during the luteal phase on reproductive, metabolic, and endocrine parameters in cyclic gilts. J. Anim. Sci. 79, 200-212. [ Links ]
Arias, P., Jarry, H., Leonhardt, S., Moguilevsky, J. & Wuttke, W., 1993. Estradiol modulates the LH release response to N-methyl-d-aspartate in adult female rats: studies on hypothalamic luteinizing hormone-releasing hormone and neurotransmitter release. Neuroendocrinology 57, 710-715. [ Links ]
Cox, N.M., Stuart, M.J., Althen, T.G., Bennet, W.A. & Miller, H.W., 1987. Enhancement of ovulation rate in gilts by increasing dietary energy and administering insulin during follicular growth. J. Anim. Sci. 64, 507-516. [ Links ]
Gill, J.L. & Hafs, H.D., 1971. Analysis of repeated measurements of animals. J. Anim. Sci. 33, 331. [ Links ]
Littel, R.C., Freund, R.J. & Spector, P., 1991. SAS System for Linear Models, (3rd ed.). SAS Institute INC, Cary, North Carolina., USA. [ Links ]
Muturi, K.N., Birnie, L.M., Struthers, J., Wheelhouse, N.M. & Lomax, M.A., 2002. The effect of rumen protected protein on plasma insulin, IGF-1 and glucose in sheep. University of Aberdeen. UK. http://www.bsas.org.uk/meetings/annlproc/PDF99/163.pdf [ Links ]
Peters, A.L. & Mayer, B.D., 1993. Protein and fat effects on glucose responses and insulin requirements in subjects with insulin-dependent diabetes mellitus. Am. J. Clin. Nutr. 58, 555-560. [ Links ]
Picarel-Blanchot, F., Alvarez, C., Bailbe, D., Pascual-Leone, A.M. & Portha, B., 1995. Changes in insulin action and insulin secretion in the rat after dietary restriction early in life: Influence of food restriction vs. low-protein restriction. Metabolism 44, 1519-1526. [ Links ]
Snedecor, G.W. & Cochran, W.G., 1967. Statistical Methods. (6th ed.). The Iowa State Univ. Press, Ames, USA. [ Links ]
Spicer, L.J., Alpizar, E. & Echternkamp, S.E., 1993. Effects of insulin, IGF-1 and gonadotrophins on bovine granulose cell proliferation, progesterone production in vitro. J. Anim. Sci. 71, 1232-12421. [ Links ]
Whitaker, D.A., Smith, E.J., Rosa, G.O.D. & Kelly, J.M., 1993. Some effects of nutrition and management on the fertility of dairy cattle. Vet. Rec. 133, 61-64. [ Links ]
Williams, S.A., Yaakub, H., O'Callaghan, D., Boland, M.P. & Scaramuzzi, R.J., 1997. Ovulation rate in ewes infused with glucose. J. Reprod. Fert. 109, 57-63. [ Links ]
# Corresponding author. E-mail: cmeza2000@hotmail.com


{kind=link}