










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.34 n.3 Pretoria 2004
Slaughter traits of Merino sheep divergently selected for multiple rearing ability
J.J.E. CloeteI, II,; S.W.P. CloeteI, III; L.C. HoffmanI; J.E. FourieIII
IDepartment of Animal Sciences, University of Stellenbosch, Private Bag X1, Matieland 7602
IICape Institute for Agricultural Training: Elsenburg, Private Bag X1, Elsenburg 7607
IIIElsenburg Agricultural Development Institute, Private Bag X1, Elsenburg 7607
ABSTRACT
Carcass data of 114 19-month-old Merino sheep descended from two lines that were divergently selected for maternal multiple rearing ability (H and L lines, respectively) were used. In study A only ram progeny of the same age were slaughtered, while ewes and rams of the respective selection lines were included in study B.Study A: Mean slaughter weight of H line rams was 12% heavier than that of L line contemporaries. A corresponding difference of 13% was found for carcass weight. Carcass component weights, body measurements, retail cut weights and eye-muscle areas were higher in the H line than in the L line, barring a few exceptions. Adjustment for the higher slaughter weight resulted in most of the line differences being eliminated, but the difference in the loin retail cut remained in favour of the H line.Study B: Mean slaughter and carcass weights of H line animals were respectively 7% and 11% heavier than that of L line contemporaries. Adjustment for the higher slaughter weights of H line animals resulted in most of the differences in the retail cut weight being eliminated, but the hindquarters of H line animals remained heavier than that of L line contemporaries. Line did not affect the moisture, protein, lipid or ash content of the M. longissimus dorsi toracis in study B. The effect of gender on the retail cut weights and proximate analysis was consistent with results from the literature. Selection for multiple rearing ability did not result in any unfavourable responses in retail cut weights or meat chemical properties. Carcasses in the H line, in fact, yielded better in the highly priced loin and hindquarters areas, independently of slaughter weight.
Keywords: Carcass composition, meat yield, proximate analysis, reproduction, retail cuts
Introduction
Reproduction of sheep plays a major role in the economic viability of the industry in South Africa (Olivier, 1999). Breeding is one of the avenues to be explored in a quest for an improved reproduction rate. Reproduction and survival rates are two important factors determining the efficiency of lamb production (Olivier et al., 1998). Prolificacy of the dam and the growth rate and slaughter quality of her offspring determine the efficiency of a lamb-meat production system (Visscher et al., 2000). The effect of the presence of the Booroola F gene on reproduction has been studied widely (Metherell, 1984; Ponzoni et al., 1985; Bradford & Quirke, 1986). The impact of reproduction on carcass parameters, however, received much less attention. Visscher et al. (2000) found that Merino crosses that include the Booroola F gene had bigger and fatter carcasses compared to their contemporaries without the gene. The consumer prefers lean meat at present, and an increase in fatness is thus not acceptable. Greeff et al. (2003) recently estimated genetic parameters for meat quality traits, as well as genetic correlations of meat quality with wool traits. The latter study was motivated against the increase of the importance of meat production in the traditional wool-producing Merino industry, and a lack of knowledge on the impact of improvements in the meat production capacity of Merinos on wool traits.
If economic production and consumption are striven for, attention has to be paid to meat quality (Sañudo et al., 1998). Jeremiah et al. (1997) also expressed the need for definite research to provide guidelines to optimise the balance between carcass weight, quantitative yield of retail cuts and meat quality. Meyer et al. (1998) studied the effects of the callipyge gene on lamb growth, carcass traits, muscle weight and meat characteristics. They found that lambs with the callipyge gene had higher growth rates and muscle weights, but that the shearing value of the meat was high, making it less acceptable to consumers.
It is evident that there is a lack of knowledge of the effects of selection for other traits on aspects of meat production. The effects of selection for an increased reproduction rate on carcass weight, mutton production, retail cut yield and chemical composition in Merino sheep has not yet been studied. This paper examines the effect of divergent selection for an increased reproduction rate on those traits in similar-aged Merino sheep.
Materials and Methods
Data of 114 19-month-old Merino sheep (40 ewes and 74 rams) were used in this study. The study was conducted in two sections over two years. Only rams (52) were slaughtered over a two-week period in study A. Twenty-two rams and 40 ewes were slaughtered over a three-week period in study B. The sheep were descended from two selection lines, a high (H) line and a low (L) line. Numbers within lines were 89 and 23 individuals, respectively. The lines have been divergently selected for maternal multiple rearing ability since 1986. Cloete et al. (2004) have described the selection procedure in detail. In brief, ewe and ram progeny of ewes rearing more than one lamb per joining (i.e. that reared twins at least once) were preferred as replacements in the H line. Descendants of ewes that reared less than one lamb per joining (i.e. that were barren or lost all lambs born at least once) were preferred as replacements in the L line (Cloete et al., 2004). The sheep used in this study were roughly subjected to four to five generations of selection.
All the sheep in the same study (A or B) were maintained in the same flock from birth until slaughter at 19-months of age. In study B, two flocks were maintained, separated on gender. Both lines were subjected to the same level of husbandry (e.g. parasite control) during this period. The sheep were shorn approximately 21 days prior to being slaughtered. Full body live weight was determined 24 hours prior to slaughtering (Hopkins et al., 1996). The sheep were slaughtered at a commercial abattoir using standard South African slaughter techniques, which included electrical stunning (4 seconds at 200 Volts) before being exsanguinated. The carcasses were subsequently hung to bleed out and then skinned. One day after slaughter the dressed carcasses were transported from the abattoir to a deboning facility, where they were kept for another 24 hours prior to deboning and sampling.
In study A recordings on the carcass included the weight of carcass components, cold carcass weight (after 24 hours in a cooler at 2 oC), the weight of the retail cuts and backfat depth. The latter was taken at a site 25 mm off the midline at the 13th rib (Gilmour et al., 1994). Carcass components that were weighed included the head, trotters and skin. Carcass length was measured on the hanging carcass from the anterior tip of the pubis bone to the front of the first rib. The leg circumference was measured at two points: the first leg circumference (1) was taken at the maximum circumference of a line passing over the distal end of the iliac wings of the pelvis and the most caudal point on the median line between the legs (Stanford et al., 1997), and the second leg circumference (2) was taken at the stifle (Oman et al., 1999). Hind leg length was measured from the inner edge of the proximal end of the tibia to the anterior tip of the pubis (Enright, 1990). The eye-muscle area was also measured at the 13th rib (Gilmour et al., 1994) by tracing the eye-muscle circumference onto wax paper. The silhouette of the eye muscle was then passed through a Li Cor LI3100 (1 mm2 resolution) for the determination of surface area. Each silhouette was measured five times. The five replicates were then used to calculate a mean for usage in the statistical analysis of the data.
After 48 hours in the cooler the carcasses were partitioned into different retail cuts, which were weighed separately. These cuts consisted of the neck, shoulder, chuck, flatrib, prime rib, loin and hindquarters. The neck was removed at the seventh cervical vertebrae (the point where the neck starts bending), the cut being made at right angles to the spine. Thereafter the hind legs were removed. This consisted of loosening the flanks on the inside of the legs (following the curve of the leg muscle) to an imaginary line perpendicular/vertical to the ilium (seen from the inside of the carcass). The leg was then removed by cutting along this line, just missing the ilium (through the last lumbar vertebrae). The rest of the carcass was then halved prior to being separated into trade cuts. The shoulder was removed by sawing along an imaginary line from the elbow joint to a point below the spinal column, between the fifth and sixth ribs. The carcass was then swivelled so that the spinal column was sawn through at right angles. The flank was removed by sawing from the M. obliquus abdominis internus parallel to the spine. The loin and rib were separated perpendicularly to the spinal column at the junction of the thoracic and lumbar vertebrae (Hoffman, 2000). All commercial cuts were weighed on a digital computing scale to the nearest gram.
In study B the same slaughtering procedures were used as in study A. Most of the recordings in this study were similar to those in Study A. In addition to these measurements the carcass depth and width were also taken on the hanging carcass (Kenney et al., 1995). After 48 hours the carcasses were cut into the different retail cuts, which were weighed separately (Hoffman, 2000). These cuts were the same as in study A, except for the hind legs, which were cut at the second last lumbar vertebrae and not the last. The hind leg length and circumference were also measured. The backfat depth was taken at a site 25 mm off the midline at the 13th rib. Individual eye-muscle areas were also determined as in study A.
The loin retail cut of 26 carcasses (6 H line and 6 L line rams; 7 H line and 7 L line ewes from study B) were randomly removed and taken to the laboratory for the measurement of proximate chemical analysis. The M. longissimus dorsi toracis was dissected, minced, freeze-dried and analysed for proximate chemical composition. The protein was measured by a FP-428 Nitrogen and Protein Determinator (Leco). Lipid (petroleum ether extraction) was measured according to AOAC (1995). Moisture content was determined by drying a sample (ca. 1.0 g) at 100 oC to a constant weight and ash content by cremation overnight at 500 oC (AOAC, 1995).
Prior to testing for differences of the various parameters between the two lines and sexes, an analysis of variance was done according to a 2 (lines; H and L) x 2 (birth type; singles and pooled multiples) factorial design in study A. Least squares procedures were used to account for uneven subclasses in an unbalanced design (Harvey, 1990). These methods have been successfully used over the past few decades for the analysis of unbalanced designs in animal breeding experiments (Harvey, 1982). Line did not interact (P < 0.05) with birth type in any of the studies and these results are not presented. There was also no significant interaction between sex and line in study B. It was found that the two lines diverged for line weight, H line animals becoming heavier and L line animals becoming lighter (Cloete & Olivier, 1998). Corresponding results were obtained in initial analyses during this study. In subsequent analyses, the higher mean slaughter weight of individuals in the H line was accounted for by analysis of covariance. Only the main effects of line (both studies) and sex (study B) are thus presented. The differences between the various parameters determined for the two lines and between sexes were tested separately by means of the null hypothesis (Ho) with Ho: μ = μo, and the alternative hypothesis (Hμ) being Hμ ≠μo. This was done by means of contrast analysis and estimated least square means as reported in the tables. Differences between the variables were accepted as being significant if the possibility of rejection of Ho was equal to or less than 5% (P < 0.05) between the two lines or sexes.
Results
Study A: The mean slaughter weight of the H line rams was 12% heavier (P < 0.01) than that of their L line contemporaries (Table 1). A corresponding difference (P < 0.01) of 13% was found for the mean carcass weight. Mean carcass component weights, retail cut weights and eye-muscle area were higher (P < 0.05) in the H line than in the L line, barring a few exceptions. Among these was a suggestion of a heavier mean skin weight (P = 0.20) in L line animals compared to H line contemporaries. Mean carcass length and leg circumference were higher (P < 0.01) in the H line than in the L line (Table 1).
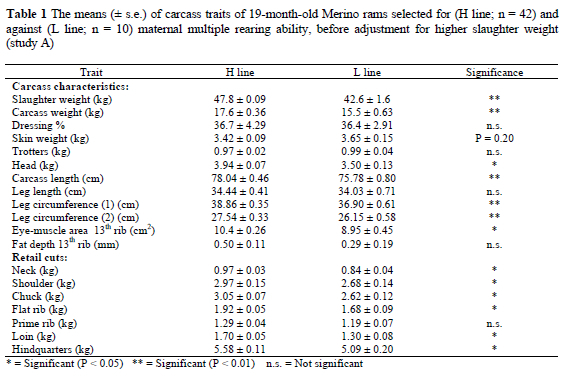
Adjustment for the higher mean slaughter weight of H line rams by analysis of covariance resulted in most of the line differences being eliminated (Table 2). The difference in the adjusted mean weight of the loin retail cut remained heavier in the H line after being adjusted for higher slaughter weight (P < 0.05). The mean skin weight was heavier (P < 0.01) in L line rams than in H line contemporaries after adjustment for slaughter weight. Rams in the L line also had heavier (P < 0.05) trotters than their H line contemporaries after adjustment for slaughter weight.

Study B: Since there were no significant sex x line interactions the data were presented as the main effects of line and sex, although the interaction was incorporated in the final analysis. The mean slaughter weight before adjustment for higher slaughter weight of H line animals was only 7% heavier (P = 0.05) than that of the L line contemporaries in this study (Table 3). A corresponding difference (P < 0.05) of 11% was found for carcass weight between the two lines.
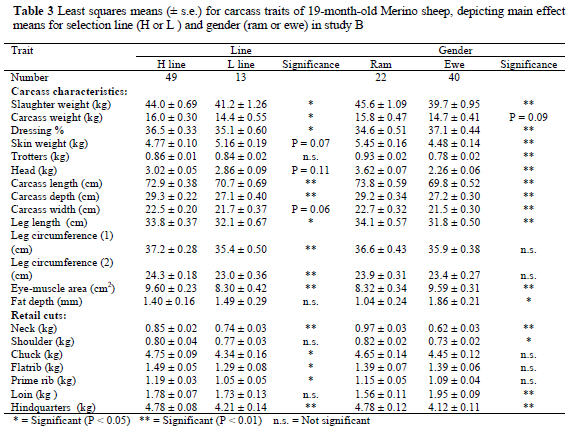
In study B most of the line differences corresponded to study A. There were no differences between lines for the weights of the trotters, as well as for the shoulder and loin retail cuts (Table 3). The mean carcass length (P < 0.01) and leg circumference (P < 0.01) were higher in the H line than in the L line. The mean leg length of the H line was longer than that of the L line animals (Table 3).
Mean slaughter weight of the rams was 15% heavier (P < 0.01) than that of ewes (Table 3). However, there was only a tendency for rams to have heavier carcasses than ewes (P = 0.09). The neck and shoulder retail cut weights were higher for rams than for ewes. No gender differences were found for the chuck, flatrib and prime rib retail cuts. The loin retail cut and eye-muscle area of the ewes were higher (P < 0.01) than those of the rams. The mean carcass length, depth and width of rams were higher (P < 0.01) than in ewes, but there were no differences in leg circumferences.
Adjustment for the higher mean slaughter weight of H line animals by analysis of covariance resulted in most of the line differences being eliminated (P > 0.05). The adjusted means for the parameters that still tended to differ (P < 0.10) between the two selection lines are shown in Table 4. The difference in the mean adjusted weight of the retail hindquarter cut remained significantly higher for the H line after being adjusted for higher slaughter weight (P < 0.01). The mean carcass length, depth (P < 0.01) and leg circumferences also remained higher (P < 0.05) in the H line after adjustment for slaughter weight. The adjusted mean skin weight (P < 0.01) was heavier in L line animals than in H line contemporaries, as in Study A.
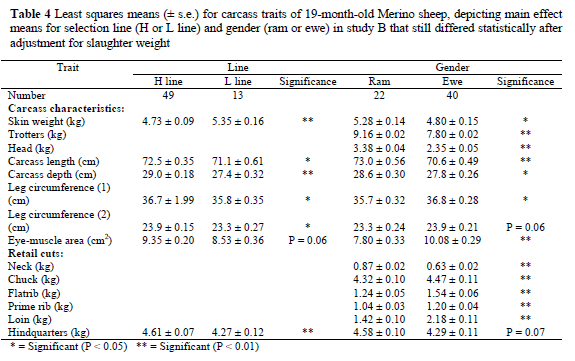
After adjustment for the higher mean slaughter weight of the rams by analysis of covariance, all the differences in carcass component weights remained significantly higher in the rams, the exception being leg circumference (1) that were higher (P < 0.05) in the ewes than in the rams. In all the adjusted mean retail cuts, weights were generally heavier (P < 0.05) in the ewes, except for the neck and hindquarters weights that were heavier (P < 0.05) in rams (Table 4).
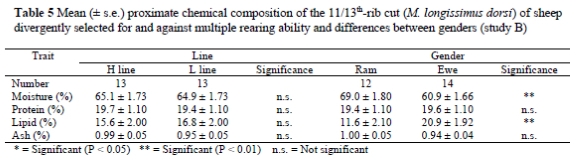
According to Table 5, there was no difference between the H and L line for mean proximate chemical composition of the M. longissimus dorsi toracis and subcutaneous fat (sampled from the 1st to the 6th lumbar vertebrae). The moisture content of the meat of rams was higher (P < 0.01) than that of ewes, and the lipid content of the meat from ewes was higher (P < 0.01) than that of rams.
Discussion
Previous research has shown divergent genetic trends for live weight in the H and L lines (Cloete & Olivier, 1998). It thus comes as no surprise to find significant differences in slaughter weight between lines. Most of the carcass component weights, retail cut weights and eye-muscle areas were consequently higher in the H line than in the L line. Animals from the H line also had a higher dressing percentage that L line contemporaries in both studies. This observation could possibly be related to the higher skin weight in the latter line, when related to slaughter weight. The heavier skin weight of the L line was probably related to skin thickness, which was shown to be thicker in the L line than in the H line (Cloete, 2002). Williams & Thornberry (1992) found that Merinos that were offspring from a high wool-producing flock had thicker skins in comparison with a low wool-producing flock. However, in this flock, there was no conclusive difference in genetic trends for two-tooth clean fleece weight between the H and L lines (Cloete & Olivier, 1998). Williams & Thornberry (1992) found a positive (P < 0.05), although small, relationship between skin thickness and wrinkling. Cloete & Olivier (1998) reported that the genetic change in fold score in the L line flock was positive (i.e. the animals became more developed), while H line animals became plainer. This could explain why L line animals had heavier skins than H line contemporaries when adjusted for slaughter weight.
The larger eye-muscle areas of the H line animals in both studies, and after adjustment for slaughter weight in study B suggested a higher muscularity in the H line. Eye-muscle area was found to be a significant predictor of leg muscle (Hopkins et al., 1992). Progeny of the H line had heavier hindquarter weights in study B after adjustment for slaughter weight. This finding, together with corresponding differences in leg circumference in favour of H line sheep in Study B after adjustment for slaughter weight, supports this line of reasoning. The differences between the results in the two parts of the experiment (heavier loins in the H line in Study A vs. heavier hindquarters in the H line in Study B) could be due to the difference in the partitioning of the retail cuts by different blockmen.
Chemical analysis of the M. longissimus dorsi toracis showed no differences between the selection lines. A previous study on a smaller sample from the same population and using samples from the M. semimembranosus similarly found no significant line differences (Hoffman et al., 2003).
The difference in dressing percentage between the rams and ewes could be due to the fact that the ewes were fatter than the rams, and the heads and trotters of the rams were heavier than those of the ewes. The average skin weight of rams was also higher than that of ewes. Ewes had larger eye-muscle areas than the rams after adjustment for slaughter weight. Fahmy et al. (1999) also found that ewes had heavier loin retail cuts and bigger eye-muscle areas than rams, although they worked on rams and ewes that were heterozygous or homozygous for the Callipyge gene. Jeremiah et al. (1997) correspondingly reported that rams had heavier neck retail cuts than ewes. This phenomenon of heavier forequarters in rams is typical of sexual dimorphism, and is expected when sheep are slaughtered after sexual maturity. The animals in these two studies were all 19 months old and therefore sexually mature.
Teixeira et al. (1996) stated that ewes were fatter than rams of the same age. Similar results were found in this study, as reflected by a higher backfat thickness and higher lipid content in the proximate chemical analysis of the M. longissimus dorsi toracis for ewes. Ram carcasses contained a higher percentage of moisture in the M. longissimus dorsi toracis than ewe carcasses, which agrees with the results of Kemp et al. (1976).
Conclusion
Selection for the ability to rear multiples thus resulted in higher carcass weights of H line animals at the same age as L line animals. This is an important observation from the producer's point of view, because of the obvious monetary advantages. Line differences in favour of the L line were in comparatively lower-priced carcass components, e.g. the skin (in both studies) and trotters (study A). The consistent differences in loin (study A) and hindquarter weight (study B) in favour of the H line suggested a higher percentage of the most valuable meat in this line. The outcome of this study indicates that selection for overall reproduction rate improved meat attributes, without any unfavourable correlated response. This is unlike the response to selection for highly fecund sheep, by using the Booroola gene, which led to an increased fat content (Visscher et al., 2000). The Callipyge gene correspondingly resulted in a marked improvement in muscling, but at the expense of tenderness (Meyer et al., 1998).
Acknowledgement
The financial contributions of the South African wool industry to the maintenance of the breeding flock, the Technology and Human Research for Industry Programme (THRIP) and the National Resources Foundation (NRF) of South Africa are gratefully acknowledged. We also wish to thank the personnel of Tommi's and Roelcor abattoirs for their friendly co-operation.
References
AOAC, 1995. Official methods of analysis (16th ed.). Association of Official Analytical Chemists International, Method 4.5.01 (920.39), Maryland, USA. [ Links ]
Bradford, G.E. & Quirke, J.F., 1986. Ovulation rate and litter size of Barbados, Targhee and crossbred ewes. J. Anim. Sci. 62, 905-909. [ Links ]
Cloete, J.J.E., 2002. Carcass traits in relation to genotype in sheep. M.Sc. (Agric.) thesis, University of Stellenbosch, South Africa. [ Links ]
Cloete, S.W.P., Gilmour, A.R., Olivier, J.J. & van Wyk, J.B., 2004. Genetic and phenotypic trends and parameters in reproduction, greasy fleece weight and live weight in Merino lines divergently selected for multiple rearing ability. Aust. J. Exp. Agric. 44, 745-754. [ Links ]
Cloete, S.W.P. & Olivier, J.J., 1998. Direct and correlated responses to divergent selection for multiple rearing ability in South African Merinos. Proc. S. Afr. Soc. Anim. Sci. 36, 65-68. [ Links ]
Enright, W.J., Quirke, J.F., Gluckman, P.D., Breier, B.H., Kennedy, L.G., Hart, I.C., Roche, J.F., Coert, A. & Allen, P., 1990. Effects of long-term administration of pituitary-derived bovine growth hormone and estradiol on growth in steer. J. Anim. Sci. 68, 2345-2350. [ Links ]
Fahmy, M.H., Gariepy, C. & Fortin, J., 1999. Carcass quality of crossbred lambs expressing the callipyge phenotype born to Romanov purebred and crossbred ewes. Anim. Sci. 69, 525-533. [ Links ]
Gilmour, A.R., Luff, A.F., Fogarty, N.M. & Banks, R., 1994. Genetic parameters for ultrasonic fat depth and eye muscle measurements in live Poll Dorset sheep. Aust. J. Agric. Res. 45, 1281-1291. [ Links ]
Greeff, J.C., Davidson, R. & Skerrit, J.W., 2003. Genetic relationship between carcass quality and wool production traits in Australian Merino rams. Proc. Assoc. Advmt Anim. Breed. Gen. 15, 330-333. [ Links ]
Harvey, W.R., 1982. Mixed model capabilities of LSML76. J. Anim. Sci. 54, 1279-1285. [ Links ]
Harvey, W.R., 1990. User's Guide for LSMLMW and MIXMDL. PC-2 version (Mimeograph: Columbus, Ohio, USA). [ Links ]
Hoffman, L.C., 2000. The yield and carcass chemical composition of impala (Aepyceros melampus), a southern African antelope species. J. Sci. Food Agric. 80, 752-756. [ Links ]
Hoffman, L.C., Schmidt, D., Muller, M.M., Cloete, J.J.E. & Cloete S.W.P., 2003. Sensory and objective quality characteristics of SA Merino sheep selected for and against reproductive fitness. S. Afr. J. Anim. Sci. 33, 52-64. [ Links ]
Hopkins, D.L., Gilbert, K.D., Pirlot, K.L. & Roberts, A.H.K., 1992. Elliottdale and crossbred lambs: growth rate, wool production, fat depth, saleable meat yield, carcass composition and muscle content of selected cuts. Aust. J. Exp. Agric. 32, 429-434. [ Links ]
Hopkins, D.L., Hall, D.G. & Luff, A.F., 1996. Lamb carcass characteristics 3. Describing changes in carcasses of growing lambs using real-time ultrasound and the use of these measurements for estimating the yield of saleable meat. Aust. J. Exp. Agric. 36, 37-43. [ Links ]
Jeremiah, L.E., Jones, S.D.M., Tong, A.K.W., Robertson, W.M. & Gibson, L.L., 1997. The influence of lamb chronological age, slaughter weight and gender on yield and cutability. Sheep Goat Res. J. 13, 39-46. [ Links ]
Kemp, J.D., Johnson, A.E., Stewart, D.F., Ely, D.G. & Fox, J.D., 1976. Effect of dietary protein, slaughter weight and sex on carcass composition, organoleptic properties and cooking losses of lamb. J. Anim. Sci. 42, 575-583. [ Links ]
Kenney, P.A., Goddard, M.E. & Thatcher, L.P., 1995. Genetic parameters for terminal sires estimated using data of progeny from Border Leicester x Merino ewes. Aust. J. Agric. Res. 46, 703-719. [ Links ]
Metherell, J.A., 1984. Management and breeding policies for the use of the Booroola F gene for increased flock prolificacy. Proc. N. Z. Soc. Anim. Prod. 44, 37-40. [ Links ]
Meyer, H.H., Haribaskar, S., Abdulkhaliq, A.M. & Thompson, J.M., 1998. Callipyge gene effects on lamb growth, carcass traits, muscle weights and meat characteristics. 6th World Congress on Genet. Appl. Livest. Prod. Sci. 25, 161-172. [ Links ]
Olivier, J.J., 1999. The South African Merino performance testing scheme. In: Premium Quality Wool Symposium. Proc. Assoc. Advmnt Anim. Breed. Genet. 13, 119-124. [ Links ]
Olivier, W.J., Snyman, M.A., Van Wyk, J.B. & Erasmus, G.J., 1998. Genetic parameter estimates for fitness traits in South African Merino sheep. Livest. Prod. Sci. 56, 71-77. [ Links ]
Oman, J.S., Waldron, D.F., Griffin, D.B. & Savell, J.W., 1999. Effects of breed-type and feeding regimen on goat carcass traits. J. Anim. Sci. 77, 3215-3218. [ Links ]
Ponzoni, R.W., Fleet, M.R., Walkley, J.R.W. & Walker, S.K., 1985. A note on the effect of the F gene on wool production and live weight of Booroola X South Australia Merino lambs. Anim. Prod. 40, 367-369. [ Links ]
Sañudo, C., Sierra, I., Olleta, J.L., Martin, L., Campo, M.M., Santolaria, P., Wood, J.D. & Nute, G.R., 1998. Influence of weaning on carcass quality, fatty acid composition and meat quality in intensive lamb production systems. Anim. Sci. 66, 175-187. [ Links ]
Stanford, K., Woloschuk, C.M., McClelland, L.A., Jones, S.D.M. & Price, M.A., 1997. Comparison of objective external carcass scores for prediction of lamb carcass quality. Can. J. Anim. Sci. 77, 217-223. [ Links ]
Teixeira, A., Delfa, R. & Treacher, T., 1996. Carcass composition and body fat depots of Galego Bragnançano and crossbred lambs by Suffolk and Merino Precoce sire breeds. Anim. Sci. 63, 389-394. [ Links ]
Visscher, A.H., Dijkstra, M., Lord, E.A., Süss, R., Rösler, H-J., Heylen, K. & Veerkamp, R.F., 2000. Maternal and carrier effects of the Booroola gene on food intake, growth and carcass quality of male lambs. Anim. Sci. 71, 209-217. [ Links ]
Williams, A.J. & Thornberry, K.J., 1992. The skin of medium wool Merino sheep and its relationship to wool production. Proc. Aust. Soc. Anim. Prod. 19, 138-141. [ Links ]
# Corresponding author. E-mail: jasperc@elsenburg.com

