










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.33 no.1 Pretoria 2003
REVIEW ARTICLE
Genetic markers and their application in livestock breeding in South Africa: A review
E. van Marle-KösterI, #; L.H. NelII
IDepartment of Animal & Wildlife Sciences, , University of Pretoria, Pretoria 0002, South Africa
IIDepartment of Microbiology & Plant Pathology, University of Pretoria, Pretoria 0002, South Africa
ABSTRACT
The development of molecular biological techniques has created new possibilities for the selection and genetic improvement of livestock. The discovery of the PCR had a major impact on the research of eukaryotic genomes and contributed to the development and application of various DNA markers. DNA markers have already found wide application in parentage verification, individual identification and control of genetic disorders. The ultimate use of DNA markers would be to identify quantitative trait loci (QTL) in order to practice genotypic selection. This paper reviews DNA markers (RAPD, DFP, RFLP AFLP, minisatellites, microsatellites, SNP) and provides a brief overview of the current application of these markers in animal breeding.
Keywords: DNA markers, molecular tools, genome mapping, QTL
Introduction
Traditionally, the genetic improvement of livestock breeds has been based on phenotypic selection. The past century was characterized by the development of quantitative theory and methodology towards the accurate selection and prediction of genetic response (Walsh, 2000). This resulted in the selection of a number of economically important genetic traits in cattle, sheep, pigs and poultry. Discoveries made by Landsteiner in the early 1900's on human blood group variability and by Erlich & Morgenroth and Todd & White on blood groups in farm animals, set the scene for research into immuno-genetics and genetic variability among animals (Hines, 1999). As early as 1940, Irwin and co-workers at the University of Wisconsin used blood group antigens for parentage verifications in the Holstein Friesians (Hines, 1999). During the 1950's Stormont (Hines, 1999) studied the blood group systems in cattle and the applications of blood groups proved to be a powerful tool in the detection of incorrect parentage, with obvious and significant implications for stud breeders. It is, however the development of molecular biology during the past three decades that created exciting new means for studying livestock genetics and animal breeding. Selection according to genotype has indeed become an important tool in the breeding of farm animals. In very recent years the molecular biological approach has also been revolutionized by the application of new technologies. In this regard one of the major role players has been the conceptually simple technique of polymerase chain reaction (PCR), which can be used to dramatically increase any target DNA in a logarithmic and controlled fashion (Nicholas, 1996).
Polymerase chain reaction was conceived in 1983 although the first related publications only started to appear in 1985. In 1989, Science magazine selected PCR as the "major scientific development", Taq polymerase (the enzyme essential to the success of PCR) as "molecule of the year" and in 1993, Kary Mullis received the Nobel Prize (Chemistry) for his role in developing PCR. Although its impact has been extraordinary, the concept of DNA amplification by PCR is simple. The chemistry involved in PCR depends on the complementarity (matching) of the nucleotide bases in the double-stranded DNA helix. When a molecule of DNA is sufficiently heated, the hydrogen bonds holding together the double helix are disrupted and the molecule separates or denatures into single strands. If the DNA solution is allowed to cool, the complementary base pairs are able to restore the original double helix. In order to use classical PCR, the exact sequence of nucleotides that flank (are situated on either side of) the area of interest (the target area to be amplified), must be known. This is the absolute minimum data necessary before a classical PCR reaction can be used. The researcher must determine or use previous data (known sequence data) to synthesize complementary 5'-3' oligonucleotides (primers) of about 20 nucleotides. However, in using PCR for genotyping purposes, several random priming strategies are commonly employed, as will be discussed during the course of this review. After priming, the key to the amplifiction of the DNA is a heat stable polymerase. This enzyme reads the original template in the 3'-5' direction and synthesizes a new complementary template in the 5'-3' direction, using free deoxynucleotide triphosphates (dNTP's) as building blocks. A PCR thus consists of a number of cycles of denaturation, annealing and extension. The product synthesized in one cycle serves as a template in the next, so that the original DNA is doubled in every cycle and the target DNA molecules are typically increased by several millionfold. The amplicon or PCR-product can then be visualized on an agarose or polyacrylamide gel (Erlich, 1991; Nicholas, 1996; Turner et al., 1998). PCR has the advantage of being a relatively fast, sensitive and reliable method. Not only can it amplify very small amounts of DNA, but can also amplify degraded or poor sources of DNA (Erlich, 1991; Strachen & Read, 1999). PCR has become an essential tool in molecular biology and plays a leading role in virtually all techniques that are presently applicable to the analysis and characterization of genomes. PCR is applied to amplify known sequences of a sample of DNA or for arbitrary priming of variable regions of the genome; the technique is suitable for use with various DNA markers such as Variable Number Tandem Repeats (VNTR's), Randomly Amplified Polymorphic DNA (RAPD), Amplified Fragment Length Polymorphism (AFLP) and Restricted Fragment Length Polymorphism (RFLP) (Erlich, 1991).
Specific genetic marker techniques
Restricted Fragment Length Polymorphism (PCR-RFLP)
This technique relies on the amplification of variable regions of the target genome, with the amplicon then being digested with one or more sequence-specific restriction enzymes. The DNA fragments of different lengths are then subjected to electrophoresis and fragments migrate according to their weights, the smaller fragments faster and the larger fragments more slowly (Nicholas, 1996). Thus, RFLP generally refers to the differences in banding patterns obtained from DNA fragments, after sequence-specific cleavage with restriction enzymes. Before the advent of PCR, RFLP analyses typically entailed restriction enzyme (RE) digestion of chromosomal DNA, followed by electrophoretic separation, membrane blotting and hybridization with a labeled probe, usually radioactive. Although reliable, this process is time-consuming, cumbersome and expensive. However, the ability to amplify a selected target a millionfold through PCR has abolished the need for the problematic blotting and hybridization analyses. Although many enzymes still need to be tested in the initial phase to be able to identify the polymorphism, it is today an easy, reliable and relatively cheap marker to use (Dodgson et al., 1997) and PCR-RFLP is, therefore, probably the simplest PCR procedure for comparisons of sequence polymorphism. Restriction enzymes (BamHI, EcoRI SacI and Taql) have often been shown to be useful in obtaining RFLP patterns for haplotype identification in individuals (Smith et al., 1996; Spike et al., 1996). Potential disadvantages of the RFLP technique include the dimorphic nature, since a RFLP only indicates the presence or absence of a cleavage site and, therefore, does not provide a great deal of genotypic information.
Random Amplified Polymorphic DNA (RAPD)
Random amplified polymorphic DNA is also known as arbitrarily primed PCR (AP-PCR), or as a DNA amplification fingerprinting technique (DAF). This technique is based on the use of short, arbitrary primers in PCR reaction and can be used to produce relatively detailed and complex DNA profiles for detecting amplified fragment length polymorphisms between organisms. In the simplest format, only one short oligonucleotide, usually eight to 10 nucleotides in length, is used. However, multiple primers are usually applied; and a range of five to 21 nucleotides has proven successful if detection is coupled with polyacrylamide gel electrophoresis. Relaxed PCR conditions allow for multiple unspecific priming sites on opposite DNA strands, even if the match is imperfect. A successfully amplified template sequence will, however, only span from a priming site sequence to a nearby complementary sequence. Depending on the primer/template combination and ratios, amplified products range from less than 10 to over a 100. In this way, a spectrum of products characteristic for each template and primer combination is typically obtained and these can be adequately resolved and visualized using polyacrylamide gel electrophoresis and silver staining. Agarose gel electrophoresis and ethidium bromide staining can also be used but detect only the major fragments.
Random amplified polymorphic DNA markers have the advantage that they can be obtained at a reasonable cost and will generally amplify a range of fragments of most DNA and show polymorphism. Certain primers will produce unrelated patterns between unrelated animals and identical ones for very closely related animals. Presumably primer sites are randomly distributed along the target genome, and flank both conserved and highly variable regions. Wide variation in band intensity can be shown to be reproducible between experiments, which could be the result of multiple copies of the amplified regions in the template or the efficiency with which particular regions are amplified. The polymorphic bands obtained from a RAPD can also be cloned for further analysis. A major disadvantage is that the RAPD primers are very sensitive to PCR conditions and this may lead to poor reproducibility (Dodgson et al., 1997). In comparison to other genetic profiling techniques discussed here, the reliability of RAPDs is at best moderate.
Amplified Fragment Length Polymorphism (AFLP)
Amplified fragment length polymorphism is a DNA fingerprinting technique that is based on the detection of DNA fragments, subjected to restriction enzymes, followed by selective PCR amplification. The DNA is cut with two restriction enzymes and double stranded adapters are then ligated to the ends of the DNA fragments to generate the template for PCR. The specific adapter, ligated to the DNA fragment, determines the sequence of AFPL primers. These primers are thus in reality semi-arbitrary and the assay determines the distribution of DNA restriction sites throughout the genomes in question by DNA amplification. AFLP procedures can be manipulated to suit specific applications through the selection of the restriction enzymes and the design of the PCR primers. Typically a rare-cutter and frequent-cutter restriction enzyme is combined to ensure the generation of small fragments (frequent-cutter) but to limit the number of fragments (rare-cutter) at the same time. PCR primers can be designed to have no selective bases on the 3' ends if the targeted templates are simple elements such as plasmids or bacterial artificial chromosomes. As in other techniques for fingerprinting, fragments are separated and analysed using gel electrophoresis. The AFLP technique can be performed at a reasonable cost, has the advantage of a higher reproducibility than RAPDs, and has been used extensively in plant genome mapping (Vos et al., 1995). More recently this technique has also found application in genetic studies of livestock with successes reported, for example, in cattle and chickens (Herbergs et al., 1999; Nijman et al., 1999).
Compared to RAPDs, higher marker densities are achieved with AFLPs and the stringency of the PCR step is also much higher in the AFLP strategy, although manipulation of band sizes and numbers is still possible. As a result AFLP is more highly reproducible and reliable. Development costs are low but running costs are higher than for RAPD analyses (1.5-2 times the cost of RAPD/sample). Being slightly more technical, the levels of skills required are also higher than those required for RAPD analysis and in addition, DNA preparations must be of particularly high quality and purity to ensure restriction enzyme digestion.
Minisatellites
Eukaryotic genomic DNA consists of non-coding and coding DNA. Thus, some parts of the genome encode for the synthesis of proteins through the translation of RNA, which is transcribed from the DNA template (coding DNA). However, non-coding DNA, which makes up a large proportion of the genomes of the eukaryotes, typically contains regulatory elements such as promoters and enhancers but can, in many cases, also contain repetitive elements, e.g. multiple repeats (Turner et al., 1998). These repetitions include satellite DNA, which comprises thousands of tandem repeats in one site, as well as mini- and microsatellite DNA, depending on the number of repeats. Minisatellites were first described as hypervariable tandem repeats, when found in the human genome. This led to the use of the term Variable Number of Tandem Repeats (VNTR) in reference to repetitive units, which include mini- and microsatellites. Minisatellites usually range in size from 10 to 100 bases and microsatellites from one to six bases (Nicholas, 1996; Turner et al., 1998). Although both minisatellites and microsatellites occur throughout the eukaryotic genome, the minisatellites tend to be concentrated in the telomere regions and in sites associated with a high frequency of recombination (Bruford & Wayne, 1993; Nicholas, 1996). The number of repeats in the satellite DNA is highly hypervariable and differs among individuals of a species. For this reason these repeats may be very effectively applied as DNA markers in the study of genetic differences. Minisatellite markers, however, have certain limitations, for example, they do not uniformly mark the genome, the marker fragment is difficult to clone and they are dominant markers if the repeat is used as a probe, which reduces the potential information for genotyping (Dodgson et al., 1997).
Microsatellites
Microsatellites consist of tandem repeats between one and six bp, repeated up to 60 times and may be referred to as "simple sequence repeats" (SSR) or "sequence-tagged microsatellite sites" (STMS). Hamada and colleagues first demonstrated these domains during the early eighties (Tautz & Renz, 1984; Smeets et al., 1989; Tautz, 1989). Repeat units may consist of (A)n, (TG)n, (CA)n or (AAT)n repeats. Microsatellites are highly polymorphic due to the variation in the number of repeats. It is not uncommon to find up to 10 alleles per locus and heterozygosity values of 60% in a relatively small number of samples (Goldstein & Polack, 1997). Microsatellites tend to mutate with mutation rates of up to 10-2 per generation (Bruford & Wayne, 1993).
A large number of microsatellite markers have been mapped for various species, including humans, mice, fruit flies and farm animals (cattle, sheep, pigs and chickens) (Goldstein & Pollack, 1997, Taylor et al., 1998; Groenen et al., 2000). These elements are most valuable markers in studies on genetic variability, parentage verifications and genome mapping projects. Various commercial kits are available for animal genotyping and animal parentage verifications for example StockMarks for Cattle® and StockMarks for Horse® parentage typing (http://www.appliedbiosystems.com). Primers developed for amplification of microsatellite loci can often be used between related species, which further promotes the application of microsatellites.
Single Nucleotide Polymorphisms (SNP)
Single nucleotide polymorphisms (also referred to as "snip") is the most recent contribution to studying DNA sequence variation. A SNP is found where different nucleotides occur at the same position in the DNA sequence. These markers are abundant and found in both coding and non-coding regions of the genome (Stoneking, 2001). It is estimated that when comparing two human DNA sequences, there will be between 1.6 and 3.2 million SNP. They are bi-allelic markers, indicating a specific polymorphism in two alleles only of a population. SNP in coding regions can be directly associated with the protein function and as the inheritance pattern is more stable, they are more suitable markers for selection over time (Beuzen et al., 2000). Most RFLP and AFLP markers are also the result of a SNP in a restriction enzyme recognition site, which confirms the importance of SNP markers. SNP have become the preferred markers in genetic disease studies in humans and they are also being searched for in various livestock species, as researches direct their attention towards functional genomics (Heaton, 2000; White et al., 2001). Reports from the "3rd International Meeting on Single Nucleotide Polymorphism and Complex Genome Analysis" indicate that there are essential technological aspects requiring more attention for a broader and cost-effective application. This includes more complete SNP databases containing descriptions of allele frequencies in various populations, haplotype and linkage disequilibrium arrangements, as well as the establishment of genotyping platforms such as Micro-arrays and Dynamic Allele Specific Hybridization (DASH) (White et al., 2001).
Application of genetic markers
DNA markers have a potential application over a relatively broad field in animal breeding and genetics. The technique has direct practical application for livestock breeders, for example for parentage verification, individual identification and identification of certain genetic disorders. A very exciting and fast developing application of genetic markers is in the mapping of the various animal genomes. Conservationists also use various genetic markers in evolutionary and genetic biodiversity studies.
Genome mapping
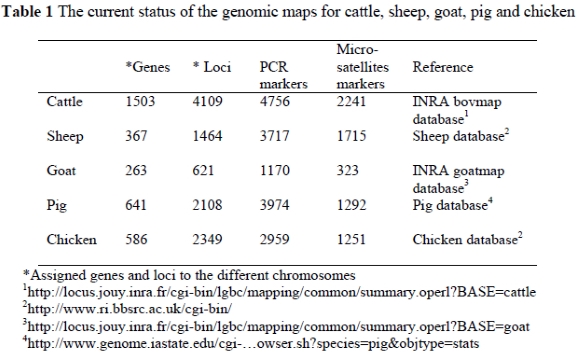
Genome mapping is almost synonymous with the Human Genome Project, which was established during 1990 as a 15 year project to map the estimated 30 000 genes in the human genome (www.ornl.gov/hgmis). The project involved 20 laboratories around the world unravelling the DNA sequences of the human genome. This project had far reaching implications for genetic research including the quest to map genomes of other species (Baltimore, 2001). The first attempt at the construction of a genetic map for the chicken genome dates back to 1936 (Fillion, 1998). Since then reference chicken families in the USA (East Lansing), UK (Compton) and the Netherlands (Wageningen) were established and several maps were constructed, which resulted in a consensus map for the chicken genome (Bumstead & Palyga, 1992; Crittenden et al., 1993; Cheng et al., 1998; Groenen et al., 2000). For most of the domesticated species such as cattle, pigs, sheep and equine, genetic maps are under construction. Ten laboratories are for example involved in the mapping of the bovine genome (http://locus.jouy.inra.fr/cgi-bin/bovmap), while six laboratories are contributing to the mapping of the sheep genome (www.projects.roslin.ac.uk/sheepmap). The current status of the genomic maps for cattle, sheep, pig and chicken are shown in Table 1. Livestock genome mapping is a complex and time-consuming effort but, once completed, holds promise for finding functional genes, Quantitative Trait Loci (QTL) and genes associated with disease resistance.
Efforts to map the genomes of different species can be expected to progress rapidly as the maps for different species are comparatively analysed (Rubin, 2001). For example, comparative mapping of the human genome and those of various farm animal species is in progress (Dodgson & Cheng, 1999; Hayes et al., 2002). A total of 154 autosomal shared segments have been identified between the chicken genome and the human genome, which may be very useful in human genetic and disease studies (Schmid et al., 2000). The research group under the leadership of J E Womack is involved in comparative mapping of the bovine genome, relative to the human and mouse genomes (www.tau.edu/gen/womack). Approximately 105 conserved segments between the human and bovine genomes have been reported by Band et al. (2000). Comparative mapping between pig chromosome 4 and human chromosome 1 is underway, searching for a QTL associated with fatness and growth, which may result in narrowing the QTL interval (Berg et al., 2002). Comparative mapping has several potential advantages. Such advantages include the identification of conserved regions between species, important contributions to the search for QTL and the provision of valuable information for gene expression studies.
Quantitative Trait Loci
In farm animals, most of the genetic traits of economic importance are the result of quantitative variation. Therefore, locating these loci, the so-called QTL, has become of great value. In order to identify a QTL for a specific trait, many animals have to be genotyped for a large number of markers on different chromosomes. Phenotypic data for the trait is also required. By means of statistical analysis, genetic and phenotypic data are combined and it is possible to find the most likely location on the chromosome responsible for the specific trait (Bovenhuis et al., 1997; Sonstegard et al., 2001). In many cases a candidate locus approach, instead of a whole genome scan, is followed in the search for a QTL (Walsh, 2000). The identification and confirmation of QTL is complex, time-consuming and also rather costly, but promises lucrative commercial returns.
In order for breeders to apply Marker Assisted Selection (MAS), QTL must be confirmed and tested. MAS could also find application where major genes are involved and with traits with low heritablity (Davis & Denise 1998). MAS programs usually require the identification of the QTL for the trait, evaluation of the QTL (where the markers are tested in a target populations) and implementation, where the animals can be genotyped and genetic and phenotypic information be combined to predict genetic merit (Davis & Denise, 1998).
Quantitative trait loci for several traits in beef cattle have been researched (Table 2). Some of these traits have already been assigned to specific chromosomes, while others are still in the process of being confirmed. In beef cattle, meat quality has to date received much of the attention in the search for QTL. The calpastatin gene associated with beef tenderness has initially been identified as a candidate gene for MAS (Marshall, 1999). Further work by Casas et al. (2000) indicated QTL affecting both growth and carcass composition in cattle associated with myostatin. A diagnostic test for a major gene for marbling in beef cattle (geneSTAR®), has now been developed and is commercially available in the USA, Australia and southern Africa (D C Nicol & Genetic Solutions. 2001; www.geneticsolutions.com). The test indicates if the animal has the favourable genotype with either one desirable allele (1-STAR) or two desirable alleles (2-STAR).
In dairy cattle the loci responsible for milk production and protein and fat content have received considerable attention. Initially, milk yield and protein quantity were linked to chromosomes 14 and 21 by Coppieters et al. (1998) and Vaiman (1999) respectively, which led to further work and the discovery of five possible regions associated with milk and protein yield, fat and protein percentage and milk yield, fat and protein yield, fat yield and protein percentage on chromosomes 1, 6, 9, 10 and 20, respectively (Vaiman, 1999). Potential QTL for fat percentage and yield and protein percentage were recently reported for chromosomes 3 and 6 respectively (Ashwell et al., 2002; Nielsen et al., 2002). Conformational type traits, associated with dairy form and milk yield have been found on chromosome 27 (Ashwell et al., 1998; Sonstegard et al., 2001).
As growth and disease are of economic importance for poultry breeders, these traits have been emphasized in the search for QTL. Van Kaam et al. (1998) tested 368 markers, scanning the whole chicken genome in their search for QTL affecting body weight in the chicken and identified chromosome 1 as the most likely position. Further work on mapping QTL for growth and fatness on chickens is underway (Jacobsson et al., 2002). The HMGI-C gene has also been identified as a candidate gene for the adw locus or autosomal dwarf gene in the chicken, which is of importance in the broiler breeding stock programmes (Ruyter-Spira et al., 1998).
There are a number of mapping projects underway searching for QTL in swine. A summary of the most recent projects compiled by Stricker (1999), includes eight research groups working on various economic important traits in swine (www.genome.iastate.edu/maps/recent/QTL-projj.html). Traits affecting reproduction such as age at puberty, ovulation rate, testicular size, and growth and carcass traits are included in the various projects (Rohrer & Keele, 1998a; b; Wang et al., 1998).
Biodiversity studies
It is inevitable that selection, inbreeding and various crossbreeding systems may lead to the loss of genetic variation within breeds and that the breed itself may become extinct. For this reason, the scientific community identified the need for the conservation of livestock resources. During 1992 the Food and Agricultural Organization (FAO) launched a programme for the Global Management of Farm Animal Genetic Resources, with the main objective being to identify conservation activities and create an awareness of possible losses of genetic resources on an international basis (Scherf, 1995; Gandini & Oldenbroek, 1999). A global programme was launched for all the different farm animal species for genetic characterization using DNA markers. Genetic markers such as DFP's, RAPD and microsatellites have been used in studying genetic variability in cattle, sheep, goats, chickens, swine and horses (Buchanan et al., 1994; Van Zeveren et al., 1995; MacHugh et al., 1997; Vanhala et al., 1998; Krüger et al., 2002). Genetic variability within and among populations is often of importance and may contribute to the selection and preservation of genetic resources. A variety of indigenous livestock breeds in South Africa have been included for genetic characterization (L. Harris, Livestock Improvement Institute, Agricultural Research Centre, Irene, RSA. Pers. Comm.).
Conclusion and South African perspective
The developments over the past ten years in molecular genetics have created the potential for the application of functional genomics over a broad field of animal improvement and breeding. In breeding, QTL will for example be applied in the genotypic selection of superior animals. In South Africa livestock breeders already have access to parentage verifications based on tests with genetic markers. Individual genotyping is done for genetic defects such as BLAD and DUMPS in dairy cattle at the ARC Livestock Improvement Institute at Irene. The ARC Animal Improvement Institute has also developed a system, based on DNA identification, to identify individual animals and prove ownership in cases of stock theft (Lidcat or Livestock Identification Catalogue). The practical application of all these technologies will to a large extent depend on the feasibility of not only the research, but also the commercialisation of the diagnostic tests. DNA technology is now well developed and provides many opportunities for genetic improvement in livestock in combination with the established quantitative methodologies. It can be expected that these technologies will, in line with worldwide trends and developments, rapidly find a broader application in South African agriculture.
References
Arranz, J.J., Coppieters, W., Berzi, P., Cambisano, N., Grisart, B., Karim, L., Marcq, F., Moreau, L., Mezer, C., Riquet, J., Simon, P., Vanmanshoven, P., Wagenaar, D. & Georges, M., 1998. A QTL affecting milk yield and composition maps to bovine chromosome 20: A confirmation. Anim. Gen. 29, 107-115. [ Links ]
Ashwell, M.S., Da, Y., Vanraden, P.M., Rexroad, C.E. & Miller, R.H., 1998. Detection of putative loci affecting conformational type traits in an elite population of United States Holsteins using microsatellite markers. J. Dairy Sci. 81, 1120-1125. [ Links ]
Ashwell, M.S., Heyen, D.W., Da, Y., Sonstegard, T.S., Tassel, C.P. & Lewin, H.A., 2002. Detection of QTL affecting milk production in 6 Diary Bull DANN Repository grandsire families. Proc. XXVIII Int. Conf. Anim. Gen., August 11-15, 2002. Göttingen, Germany. [ Links ]
Baltimore, D., 2001. Our genome unveiled. Nature 409, 814-816. [ Links ]
Band, M.R.J.H., Larson, M., Reibeiz, C.A., Green, D.W., Heynen, J., Donovan, R., Windish, C., Steining, C., Mahyuddin, P., Womack, J.E. & Lewin, H.A..2000. An ordered comparative map of the cattle and human genomes. Genome Res. 10, 1359-1368. [ Links ]
Berg, F., Archibald, A., Anderson, S. & Moller, M., 2002. Comparative genome analyses between pig chromosome 4 and human chromosome 1 and 8. Proc. XXVIII Int. Conf. Anim. Gen. August 11-15, 2002. Göttingen, Germany. [ Links ]
Beuzen, N.D., Stear, M.J. & Chang, K.C., 2000. Molecular markers and their use in animal breeding. Vet. J. 160, 42-52. [ Links ]
Bovenhuis, H., Van Arendonk, J.A.M., Davis, G., Elsen, J-M., Haley, C.S., Hill, W.G., Baret, P.V., Hetzel, D.J.S. & Nicholas, F.W., 1997. Detection and mapping of quantitative trait loci in farm animals. Livest. Prod. Sci. 52, 135-144. [ Links ]
Bruford, M.W. & Wayne, R.K., 1993. Microsatellites and their application to population genetic studies. Currt. Op. in Gen. and Dev. 3, 939-943. [ Links ]
Buchanan, F.C., Adams, L.J., Littlejohn, R.P., Maddox, J.F. & Crawford, A.M., 1994. Determination of evolutionary relationships among sheep breeds using microsatellites. Genomics 22, 397-403. [ Links ]
Bumstead, N. & Palyga, J., 1992. A preliminary linkage map of the chicken genome. Genomics 13, 690-697. [ Links ]
Casas, E., Shackelford, S.D., Keele, J.W., Stone, R.T., Kappes, S.M., Koohmaraie, M., 2000. Quantitative trait loci affecting growth and carcass composition of cattle segregating alternate forms of myostatin. J. Anim. Sci. 78, 560-569. [ Links ]
Cheng, H.H, Burt, D.W. & Dodgson, J.B., 1998. Recent advances in poultry genome mapping. Proc.: Symp. Series 8th World Conf. Anim. Prod. June 28-July 4, 1998. Seoul National University, Seoul, Korea. [ Links ]
Coppieters, W., Riquet, J., Arranz, J.J., Berzi, P., Cambisano, N., Grisart, B., Karim, L., Marcq, F., Moreau, L., Nezer, C., Simon, P., Vanmanshoven, P., Wagenaar, D. & Georges, M., 1998. A QTL with major effect on milk yield and composition maps to bovine chromosome 14. Mam. Genome 9, 540-544. [ Links ]
Crittenden, L.B., Provencher, L., Santangelo, L., Levin, H.A., Briles, R W., Briles, E. & Dodgson, J.B., 1993. Characterization of a Red Jungle Fowl by White Leghorn backcross reference population for molecular mapping of the chicken genome. Poult. Sci. 72, 334-348. [ Links ]
Davis, G.P. & Denise, S.K., 1998. The impact of genetic markers on selection. J. Anim. Sci. 76, 2331-2339. [ Links ] Dodgson, J.B. & Cheng, H.H., 1999. Poultry genomics: An alien perspective. Ag. Boitech. Net. Jan. 1999. Vol 1. ABN 005. [ Links ]
Dodgson, J.B., Cheng, H.H. & Okimoto, R., 1997. DNA marker technology: A revolution in animal genetics. Poult. Sci. 76, 1108-1114. [ Links ]
Elo, K.T., Vilkki, J., De Koning, D.J., Velmala, R.J. & Maki-Tanila, A.V., 1999. A quantitative trait locus for live weight maps to bovine chromosome 23. Mam. Genome 10, 831-835. [ Links ]
Erlich, H.A., 1991. PCR Technology: principles and applications for DNA amplification. W.H. Freeman and Company, New York. [ Links ]
Fillion, V., 1998. The chicken as a model to study microsomes in birds: review. Gen. Sel. Evol. 209-219. [ Links ] Gandini, G.C. & Oldenbroek, J.K., 1999. Choosing the conservation strategy. In: Genebanks and the conservation of farm animal genetic resources. Ed. Oldenbroek, J.K., DLO Institute for Animal Science and Health, The Netherlands. [ Links ]
Goldstein, D.B. & Pollock, D.D., 1997. Launching microsatellites: A review of mutation process and methods of phylogenetic inference. J. Hered. 10, 88, 335-342. [ Links ]
Groenen, M.A.M., Cheng, H., Bumstead, N., Benkel, B.F., Briles, W.E., Burke, T., Burt, D.W., Crittenden, L.B., Dodgson, J., Hillel, J., Lamont, S., Ponce de Leon, A., Soller, M., Takahashi, H. & Vignal, A., 2000. A consensus map of the chicken genome. Genome Res. 137-147. [ Links ]
Hayes, H., Elduque, M., Gautier, L., Schibler, E., Cribiu, E. & Eggen, A., 2002. Gene mapping progress in cattle and updated comparative map with man, mouse, rat and pig. Proc. XXVIII Int. Conf. Anim. Gen. August 11-15, 2002. Göttingen, Germany. [ Links ]
Heaton, M.P., 2000. SNP frequency and haplotype diversity in beef cattle cytokine genes. Plant & Animal Genome VIII Conf., January 9-12, 2000. San Diego, CA. [ Links ]
Herbergs, J., Siweck, M., Crooijmans, R.P.M.A., Van der Poel, J.J. & Groenen, M.A.M., 1999. Multicolour fluorenscent detection and mapping of AFLP markers in chicken (Gallus domesticus). Anim. Gen. 30, 274-285. [ Links ]
Hines, H.C., 1999. Blood groups and biochemical polymorphisms. In: Genetics of cattle. Eds. Fries, R. & Ruvinsky, A., CABI Publishing. [ Links ]
Jacobsson, L.U., Hee-Bok, P., Siegel, P.B. & Anderson, L., 2002. Mapping of Quantitative Loci for growth and fatness in chickens. Proc. XXVIII Int. Conf. Anim. Gen. August 11-15, 2002. Göttingen, Germany. [ Links ]
Kirkpatrick, B.W., Byla, B.M. & Gregory, K.E., 2000. Mapping quantitative trait loci for bovine ovulation rate. Mam. Genome 11, 136-139. [ Links ]
Krüger, K., Stranzinger, G. & Rieders, S., 2002. A full genome scan panel of horse (Equus caballus) microsatellite markers applied to different equid species. Proc. XXVIII Int. Conf. on Anim. Gen. August 11-15, 2002. Göttingen, Germany. [ Links ]
MacHugh, D.E., Shriver, M.D., Loftus, R.T., Cunningham, P. & Bradley, D.G., 1997. Microsatellite DNA variation and the evolution, domestication and phylogeography of Taurine and Zebu cattle (Bos taurus and Bos indicus). Genetics 146, 1071-1086. [ Links ]
Marshall, D.M.1999. Genetics of meat quality. In "The genetics of cattle". Eds. Fries, R. & Ruvinsky, A. CABI Publishing CAB International, UK. [ Links ]
Nicholas, F.W., 1996. Introduction to veterinary genetics. Oxford University Press. [ Links ]
Nielsen, V., Guldbrandtsen, M.S., Jensen, J., Sorensen, D.A. & Bendixen, C., 2002. Proc. XXVIII Int. Conf. on Anim. Gen. August 11-15, 2002. Göttingen, Germany. [ Links ]
Nijman, I.J., Bradley, D.G., Hanotte, O., Otsen, M. & Lenstra, J.A., 1999. Satellite DNA plymorphisms and AFLP correlate with Bos-indicus-taurus hybridization. Anim. Gen. 30, 265-273. [ Links ]
Rohrer, G.A. & Keele, J.W., 1998a. Identification of quantitative trait loci affecting carcass composition in Swine: I. Fat deposition traits. J. Anim. Sci. 76, 2247-2254. [ Links ]
Rohrer, G.A. & Keele, J.W., 1998b. Identification of quantitative trait loci affecting carcass composition in Swine: II. Muscling and wholesale product yield traits. J. Anim. Sci. 76, 2255-2262. [ Links ]
Rubin, G.M., 2001.The Draft sequences: Comparing species. Nature 409, 820-821 [ Links ]
Ruyter-Spira, Carolien, P., Gu, Z.L., Van der Poel, J.J. & Groenen, M.A.M., 1998. Bulked segregant analysis using microsatellites: Mapping of the dominant white locus in the chicken. Poult. Sci. 76, 386-391. [ Links ]
Scherf, B.D., 1995. Developing the global inventory for poultry genetic resources. Proc. Third Global Conf. on Conservation of Domestic Animal Genetic Resources. August 1-5, 1994. Queens University, Canada. Eds. Crawford, R.D., Lister, E.E. & Buckley, J.T., Rare Breeds International, Warwickshire, UK. [ Links ]
Schmid, M., Nanda, I., Guttenbach, M., Steinlein, C., Hoehn, M., Schartl, M., Haaf, T., Weigend, S., Fries, R., Buerstedde, J-M., Wimmers, K., Burt, D.W., Smith, J., A'Hara, S., Law, A., Griffin, D.K., Bumstead, N., Kaufman, J., Thomson, P.A., Burke, T., Groenen, M.A.M., Crooijmans, R.P.MA., Vignal, A., Fillon, V., Morisson, M., Pitel, F., Tixier-Boichard, M., Ladjali-Mohammedi, K., Hillel, J., M. Ki-tanila, A., Cheng, H.H., Delany, M.E., Burnside, J. & Mizuno, S., 2000. First report on chicken genes and chromosomes. Cyto. Cell Gen. 90, 169-218. [ Links ]
Smeets, E., Jones, C.P., Bartlett, J. & Nestor, K.E., 1989. Use of randomly amplified polymorphic DNA markers for the genetic analysis of relatedness and diversity in chickens and turkeys. Poultry Sci. 75, 579-584. [ Links ]
Smith, E.J, Jones, C.P., Bartlett, J. & Nestor, K.E., 1996. Use of randomly amplified polymorphic DNA markers for the genetic analysis of relatedness and diversity in chickens and turkeys. Poult. Sci. 75, 579-584. [ Links ]
Sonstegard, T.S., Van Tassel, C.P. & Ashwell, M.S., 2001. Dairy cattle genomics: Tools to accelerate genetic improvement? J. Anim. Sci.79 (E. Suppl.), E307-E315. [ Links ]
Spike, C.A., Bumstead, N., Crittenden, L.B. & Lamont, S.J., 1996. RFPL mapping of expressed sequence tags in the chicken. J. Hered. 87, 6-9. [ Links ]
Stoneking, M., 2001. Single nucleotide polymorphisms: From the evolutionary past. Nature 409, 821-822. [ Links ]
Strachan, T. & Read, A.P., 1999. Human molecular genetics (2nd ed.). BIOS Scientific Publishers. [ Links ]
Tautz, D., 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucl. Acids Res. 17, 16-18. [ Links ]
Tautz, D. & Renz, M., 1984. Simple sequences are ubiquitous repetitive components of eukaryotic genomes. Nucl. Acids Res. 12, 4127 -4138. [ Links ]
Taylor, J.F., Eggen, A., Aleysasin, A., Armitage, S.M., Barendse, W., Beever, J.E., Bishop, M.D., Brennenman, R.A., Burns, B.M., Davis, S.K., Elo, K.T., Harizius, B., Kappes, S.M., Keele, J.W., Kemp, S.J., Kirkpatrick B.W., Lewin, H.A., Ma, R.Z., McGrwa, R.A., Pomp, D., Stone, R.T., Sugimoto, Y., Teale, A.J., Vaiman, D., Vilkki, J., Williams, J.L, Yeh, C-C. & Zanotti, M.C., 1998. Report of the first workshop on the chromosome on the genetic map of bovine chromosome 1. Anim. Gen. 29: 228-235. [ Links ]
Turner, P.C., McLennan, A.G., Bates, A.D. & White, M.R.H., 1998. Instant notes in molecular biology. BIOS Scientific Publishers Limited, Oxford, UK. [ Links ]
Vaiman, D., 1999. The molecular genetics of cattle. In: The Genetics of Cattle. Eds. Fries, R. & Ruvinsky, A., CABI Publishing, CAB International, UK. [ Links ]
Vanhala, T., Tuiskula-Haavisto, M., Elo, K., Vilkki, J. & Maki-Tanilla, A., 1998. Evaluation of genetic variability and genetic distances between eight chicken lines using microsatellite markers. Poult. Sci. 77, 783-790. [ Links ]
Van Kaam, J.B.C.H.M., Van Arendonk, J.A.M., Groenen, M.A.M., Bovenhuis, H., Vereijken, A.L.J., Crooijmans, R.P.M.A., Van der Poel, J.J. & Veenendaal, A., 1998. Whole genome scan for quantitative trait loci affecting body weight in chickens using a three generation design. Livest. Prod. Sci. 54, 133-150. [ Links ]
Van Zeveren, A., Peelman, L., Van de Weghe, A. & Bouquet, Y., 1995. A genetic study of four Belgian pig populations by means of seven microsatellite loci. J. Anim. Breed. Gen. 112, 191-204. [ Links ]
Velmala, R.J., Vilkki, H.J., Elo, K.T., De Koning, D.J. & Maki-Tanila, A.V., 1999. A search for quantitative trait loci for milk production traits on chromosome 6 in Finnish Ayrshire cattle. Anim. Gen. 30, 136-143. [ Links ]
Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van de Lee, T., Hornes, M., Frijters, A., Pot, J., Peleman, J., Kuiper, M. & Zabeau, M., 1995. AFLP: A new technique for DNA fingerprinting. Nucl. Acids Res. 23, 4407-4414. [ Links ]
Walsh, B., 2000. Minireview: Quantitative genetics in the age of genomics. Theor. Pop. Biol. 59, 175-184. [ Links ]
Wang, L., Yu, T-P., Tuggle, C.K., Liu, H-C. & Rothschild, M.F., 1998. A directed search for quantitative trait loci on chromosome 4 and 7 in swine. J. Anim. Sci. 76, 2560-2567. [ Links ]
White, P.S., Kwok, P., Oefner, P. & Brookes, A.J., 2001. 3rd Int. Meeting on Single Nucleotide Polymorphism and Complex Genome Analysis: SNP's: 'Some Notable Progress'. Eur. J. Hum. Gen. 9, 316-318. [ Links ]
# Corresponding author. E-mail: ekoster@postino.up.ac.za


{kind=link}