










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSAMJ: South African Medical Journal
On-line version ISSN 2078-5135
Print version ISSN 0256-9574
SAMJ, S. Afr. med. j. vol.103 n.1 Pretoria Jan. 2013
FORUM
ISSUES IN MEDICINE
A new class of stem cells in South Africa: iPS cells
R BalloI; L J GreenbergII; S H KidsonI
IDepartment of Human Biology of the University of Cape Town (UCT)
IIDivision of Human Genetics, Department of Clinical Laboratory Sciences, Institute of Infectious Disease and Molecular Medicine, UCT
ABSTRACT
Created from adult rather than embryonic cells, induced pluripotent stem (iPS) cells represent a breakthrough in stem cell science, and their pioneers have been recognised with the 2012 Nobel Prize in Medicine. These cells offer new hope in the treatment of pathogenetic diseases, but there is still a way to go on the road to effective therapeutic applications.
The use of bone marrow, umbilical cord and other adult stem cells for the treatment of various conditions (mostly haematological) is familiar to SAMJ readers, as are the scientific and ethical debates surrounding the potential use of human embryonic stem (hES) cells for tissue repair.1,2 In the specialist medical and scientific literature, however, it is a new class of stem cells, induced pluripotent stem (iPS) cells, that are causing the greatest excitement of late, and with reason.
Human iPS cells are somatic (diploid) cells, obtained by relatively non-invasive procedures (e.g. skin biopsy), that have been genetically 'induced' to revert back to an embryonic stem-cell-like state. The term 'pluripotent' refers to a cell's ability to differentiate into any cell type in the body. Both iPS and hES cells are pluripotent. To turn a somatic cell into an iPS cell, simply stated, a very small skin biopsy is taken from a patient, a skin fibroblast cell culture is established in vitro, and the fibroblast is reprogrammed into an iPS cell. These immortal iPS cells can be grown en masse in vitro and differentiated into the particular cell type required for therapeutic or non-therapeutic purposes (Fig. 1).
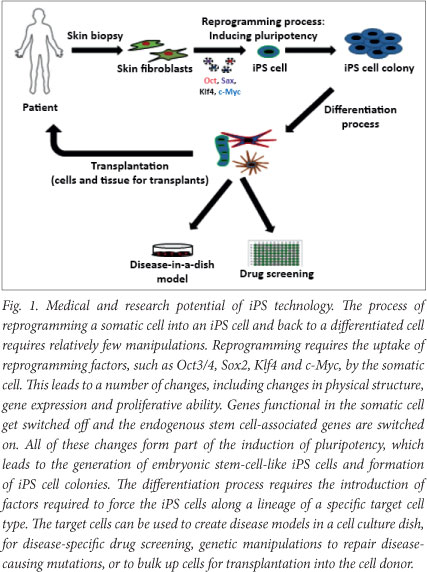
The key advantage of iPS cells over other stem cells is that they are patient-specific (and therefore immuno-compatible) and can be grown in infinite amounts. Moreover, they are not dogged by the ethical and religious controversies associated with hES cells, yet still have the same properties as hES cells. They also offer the possibility of conducting 'clinical-trials-in-the-dish', providing a platform for drug screening, disease modelling and gene/cell therapy in pre-clinical studies.3,4
How are iPS cells made?
When cell differentiation occurs, the cell follows a process of changes in gene activity whereby embryonic-specific genes are inactivated and differentiation-specific genes are activated. The end result of this differentiation 'programme' is a specialised cell of one type or another (e.g. cardiac muscle cells or neurons). To 'reprogramme' a fully differentiated adult cell into an iPS cell is surprisingly straightforward - all that is needed is reactivation of the embryonic regulatory genes and inactivation of the differentiation-specific genes.
This was achieved in ground-breaking studies in 2006/7 by Takahashi and Yamanaka, who showed that the ectopic expression of four key embryonic regulatory genes (Oct3/4, Sox2, Klf4 and c-Myc) reactivates endogenous pluripotency genes in mouse and human fibroblasts.5,6 These achievements won Yamanaka the 2012 Nobel Prize in Medicine.
iPS cells for therapeutic use
Many reports and reviews on the production of iPS cells and their possible therapeutic potential have rapidly followed. iPS cells have been derived from a wide range of cell types and differentiated into an equally broad range of cell types involved in neurological, haematological, metabolic, cardiac and vascular, pancreatic and hepatic conditions.7-10 Conditions in which treatment using iPS cells is being considered and actively pursued include genetic and acquired degenerative diseases, amelioration of the effects of ageing, and novel treatments for some of South Africa (SA)'s most serious infectious diseases. The true potential of iPS cells has yet to be established. Many concerns about their derivation must be addressed before they are ready for therapeutic use.
iPS cells and disease-in-the-dish models
Disease-in-the-dish studies have progressed much further. A broad spectrum of both monogenic diseases such as Duchenne's muscular dystrophy and Huntington's disease, as well as complex adult-onset diseases (e.g. Parkinson's, Alzheimer's and schizophrenia, to name a few) have been modelled in vitro.9,10 How effective iPS cell-derived disease models are at reproducing the disease cellular phenotype, and at demonstrating the effects of drugs, is more easily recognised in early-onset diseases that have a strong genetic component and an identifiable disease-associated cellular phenotype. This is exemplified by a patient-derived iPS cell model for familial dysautonomia, which was used to show that the disease phenotype could be partially normalised by a plant hormone.11
An example of the medical application of iPS cell technology was demonstrated by Itzhaki et al.,12 who used an iPS-model for Long-QT syndrome to identify complex effects of cardiotoxic drugs and protective pharmacological agents for disease sufferers.
Our studies of spinocerebellar ataxia 7 (SCA7), a late-onset neurodegenerative genetic disease found almost exclusively in families of black origin in SA, exemplify our use of iPS cell technology in disease modelling.13 The development of effective therapies for a group of genetic conditions known as the polyglutamine (polyQ) diseases is particularly relevant to SA populations where, for example, the frequencies of spinocerebellar ataxia type 1 (SCA1) and type 7 (SCA7) are among the highest in the world.14
To date, our research has been primarily focused on the study of the SCA7 protein in transfected neuronal cell lines and transgenic mouse models.15,16A unique clinical feature of this condition is loss of vision associated with retinal degeneration (in particular the macula), the aetiology of which is unknown. To explore the cause of retinal cell death in SCA7 patients, we have derived iPS cells from skin biopsies following standard protocols (UCT HREC ref. 460/2010 and 434/2011, both renewed in 2012 until 2013) and carried out in accordance with national stem cell regulations (promulgation in progress), cultured them en masse, and have successfully differentiated them into retinal pigment epithelial cells and neuronal cells.16 With these unique cells in hand, we are now able to investigate the cellular vulnerabilities conferred on the cells of the eyes of our South African SCA7 patients, and have a powerful tool to explore models of cell interactions and cell death. In addition, we are able to examine and observe patient-specific drug treatments that might eventually delay the onset of blindness arising as a result of cell death in South African SCA7 patients.
Risks, limitations and challenges
Despite the evident success of iPS cell technology to date, the field has been complicated by a number of challenges, not least that Takahashi and Yamanaka and many others used viral vectors (retrovirus, lentivirus) to transport the reprogramming genes into the somatic cells.5,6 The risk is that the viral integration itself could result in rogue cells with tumorigenic propensities. This is of particular concern as the technology rapidly moves towards clinical application.
Other methods of delivering reprogramming genes into cells use non-integrating agents of induction, such as the deleted persistent variant of the Sendai virus (SeVdp), viral-free non-integration approaches, proteins, mRNA and miRNA.17-22 Most non-integrating methods, however, result in reduced reprogramming efficiencies.23 In our experience, induction with the Sendai virus has been most successful following our previous utilisation of retroviruses. Following these challenges, researchers have included rigorous quality-control steps to test the purity and integrity of iPS cells before differentiation.
Conclusions
Creating cells takes time and a great deal of care, and is not suitable for acute conditions. However, the rise of large-scale bio-repositories may yet deal with this limitation. Technical research challenges and ethical controversies have delayed the progress of iPS cell technology and narrowed the scope of iPS cell banks to research projects, rather than treatment. This has resulted in recruiting iPS cells from patients and their family members rather than from larger population samples. However, in an unprecedented move, Yamanaka plans to create a bank of stem cells from fetal cord blood for therapeutic use in Japan, shifting the potential of iPS cell technology to new heights.24
It is vital that the SA medical and scientific fraternity become familiar with iPS cell technology - a reality that has demonstrated evidence of proof-of-principle.10 Researchers and clinical practitioners should be encouraged to communicate with accredited stem cell or tissue culture laboratories, and should inform patients about potential iPS cell prospects. Bearing in mind the variation of disease phenotypes and clinical manifestations, it is important to build up an SA bank of patient-derived iPS cells from different patients diagnosed with the same disease for a more accurate representation of underlying pathology. There is no doubt that iPS cell technology and patient-derived disease-in-a-dish models have the potential: (i) to exponentially increase our understanding of underlying disease mechanisms and cellular phenotype responses to drug treatments; and (ii) to introduce novel treatments for some of SA's most serious diseases.
Acknowledgements. The authors thank UCT, NRF and MRC for support of this research, and Dennis Lin for producing the figure.
References
1. Pepper MS. Cell-based therapy - navigating troubled waters. S Afr Med J 2010;100(5):286-288. [ Links ]
2. Sommer P. Stem cells - 13 years since their isolation, is there any proven benefit to the patient? S Afr Med J 2011;101(12):868-869. [ Links ]
3. Stadtfeld M, Hochedlinger K. Induced pluripotency: History, mechanisms, and applications. Genes Dev 2010;24:2239-2263. [http://dx.doi.org/10.1101/gad.1963910] [ Links ]
4. Plath K, Lowry WE. Progress in understanding reprogramming to the induced pluripotent state. Nat Rev Genet 2011;12:253-265. [http://dx.doi.org/10.1038/nrg2955 ] [ Links ]
5. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006;126(4):663-676 [http://dx.doi.org/10.1016/j.cell2006.07.024] [ Links ]
6. Takahashi,K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007;131:1-12. [http://dx.doi.org/10.1016/j.cell2007.11.019] [ Links ]
7. Park IH, Arora N, Huo H, et al. Disease-specific induced pluripotent stem cells. Cell 2008;134:877-886. [http://dx.doi.org/10.1016/j.ceE.2008.07.041] [ Links ]
8. Wu SM, Hochedlinger K. Harnessing the potential of induced pluripotent stem cells for regenerative medicine. Nat Cell Biol 2011;13(5):497-505. [http://dx.doi.org/10.1038/ncb0511-497] [ Links ]
9. Grskovic M, Javaherian A, Strulovici B, Daley GQ. Induced pluripotent stem cells-opportunities for disease modelling and drug discovery. Nat Rev Drug Discov 2011;10(12):915-929. [http://dx.doi.org/10.1038/nrd3577] [ Links ]
10. Robinton DA, Daley GQ. The promise of induced pluripotent stem cells in research and therapy. Nature 2012;481:295-305. [http://dx.doi.org/10.1038/nature10761] [ Links ]
11. Lee G, Papapetrou EP, Kim H, et al. Modelling pathogenesis and treatment of familial dysautonomia using patient-specific iPSCs. Nature 2009;461:402-408. [http://dx.doi.org/10.1038/nature08320] [ Links ]
12. Itzhaki I, Maizels L, Huber I, et al. Modelling the long QT syndrome with induced pluripotent stem cells. Nature 2011;471:225-230. [http://dx.doi.org/10.1038/nature09747] [ Links ]
13. Greenberg J, Solomon G, Vorster A, et al. Origin of the SCA7 gene mutation in South Africa: Implications for molecular diagnostics. Clin Genet 2006;70:415-417. [http://dx.doi.org/10.1111/j.1399-0004.2006.00680.x] [ Links ]
14. Smith DC, Bryer A, Watson L, Greenberg LJ. Inherited polyglutaminespinocerebellar ataxias in South Africa. S Afr Med J 2012;102(8):683-686. [http://dx.doi.org/10.7196/samj.5521] [ Links ]
15. Scholefield J, Greenberg LJ, Weinberg MS, et al. Design of RNAi hairpins for mutation-specific silencing of ataxin-7 and correction of a SCA7 phenotype. PLoS One 2009;4:e7232. [http://dx.doi.org/10.1371/ journal.pone.0007232] [ Links ]
16. Watson LM, Scholefield J, Greenberg LJ, Wood MJ. Polyglutamine disease: from pathogenesis to therapy. S Afr Med J 2012;102(6):481-486. [ Links ]
17. Li Y, Zhang Q, Yin X, et al. Generation of iPSCs from mouse fibroblasts with a single gene, Oct4, and small molecules. Cell Res 2011;21:196-204. [http://dx.doi.org/10.1038/cr.2010.142] [ Links ]
18. Nishimura K, Sano M, Ohtaka M, et al. Development of defective and persistent Sendai virus vector. J Biol Chem 2011;286(6):4760-4771. [http://dx.doi.org/10.1074/jbc.M110.183780] [ Links ]
19. Jia F, Wilson KD, Sun N, et al. A nonviral minicircle vector for deriving human iPScells. Nat Methods 2010;7(3):197-201.[http://dx.doi.org/10.1038/nmeth.1426] [ Links ]
20. Kim D, Kim CH, Moon J. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 2009;4:472-476. [http://dx.doi.org/10.1016/j.stem.2009.05.005] [ Links ]
21. Warren L, Manos P, Ahfeldt T, et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 2010;7:1-13. [http://dx.doi.org/10.1016/j.stem.2010.08.012] [ Links ]
22. Li Z, Yang CS, Nakashima K, Rana TM. Small RNA-mediated regulation of iPS cell generation. EMBO J. 2011;30(5):823-834.[ http://dx.doi.org/10.1038/emboj.2011.2] [ Links ]
23. Sommer CA, Mostoslavsky G. Experimental approaches for the generation of induced pluripotent stem cells. Stem Cell Res &Ther 2010,1:26. [http://dx.doi.org/10.1186/scrt26] [ Links ]
24. Cyranoski D. Stem-cell pioneer banks on future therapies. Nature News 7 Aug 2012. http://www.nature.com/news/stem-cell-pioneer-banks-on-fiiture-therapies-1.11129 (accessed 7 November 2012). [ Links ]
Accepted 3 December 2012.
 Corresponding author:
Corresponding author:
R Ballo (robea.ballo@uct.ac.za)

