










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSAMJ: South African Medical Journal
On-line version ISSN 2078-5135
Print version ISSN 0256-9574
SAMJ, S. Afr. med. j. vol.101 n.6 Pretoria Jun. 2011
ORIGINAL ARTICLES
Negative association of MC3R variants with weight and blood pressure in Cape Town pupils aged 11 - 16 years
Yandiswa Y YakoI; Biotumelo L FanampeIII; Shafick M HassanIV; Rajiv T ErasmusII; Lize van der MerweVI; Tandi E MatshaV
IBSc Hons (Biochem), MSc (Med Biochem), PhD (Chem Path). Department of Chemical Pathology, Faculty of Health Sciences, Stellenbosch University, Tygerberg
IIMB BS, FMC Path (Nig), FACB, DABCC (Am Board Cert), DHSM (Natal), FCPath. Department of Chemical Pathology, Faculty of Health Sciences, Stellenbosch University, Tygerberg
IIIMTech. Department of Biomedical Technology, Faculty of Health and Wellness Sciences, Cape Peninsula University of Technology, Cape Town
IVMSc. Department of Biomedical Technology, Faculty of Health and Wellness Sciences, Cape Peninsula University of Technology, Cape Town
VPhD. Department of Biomedical Technology, Faculty of Health and Wellness Sciences, Cape Peninsula University of Technology, Cape Town
VIPhD. Biostatistics Unit, MRC, Cape Town, and Department of Statistics, University of the Western Cape, Bellville
ABSTRACT
BACKGROUND: Human and animal studies support the role of MC4R and MC3R in human obesity, but limited data are available on the genetic contribution to obesity in South African populations.
OBJECTIVE: To screen obese-overweight South African pupils for MC3R and MC4R polymorphisms that may play a role in the development of obesity.
DESIGN: A cross-sectional study screened 227 obese-overweight (115 black and 112 coloured) and 204 normal weight (94 black, 110 coloured) school pupils for the presence of MC4R and MC3R polymorphisms using a single strand conformation polymorphism, subsequent sequencing, and allele specific restriction enzyme analysis.
RESULTS: Two polymorphisms were detected in the MC3R (T6K and V81I) but none in MC4R . After adjusting for age, gender and case-control status, the frequency distributions of T6K and V81I genotype and allele varied significantly between the ethnic groups. The frequency of the V81I A allele was significantly lower in coloured overweight-obesity than normal pupils. In coloured pupils, both polymorphisms were associated with obesity indices and total cholesterol. The T6K A allele was also associated with lower blood pressure. Likewise, different T6K-V81I haplotypes demonstrated negative associations with obesity indices and blood pressure.
CONCLUSIONS: We demonstrated that the MC3R polymorphisms have a protective effect on metabolic traits; however, further analysis is required to confirm whether this translates to a lower incidence of metabolic syndrome in coloured populations.
Obesity is a multifactorial body weight abnormality resulting from genetic and environmental interactions. Of the body mass index (BMI) variation among obese individuals of different populations and within the same family, 40 - 70% are estimated to be inherited, and 30% are attributed to cultural and socio-economic factors.1,2 More than 100 genes have been implicated in the heterogeneity of body weight, among which are genes that encode proteins or peptides of the central nervous system that affect conscious and unconscious aspects of energy intake and expenditure. Melanocortin receptors, particularly the melanocortin 4 receptor gene (MC4R ), play a major role in the development of both monogenic and polygenic obesity.3,4 Mutations in MC4R are the most common cause of early onset obesity, accounting for up to 6%, particularly in white populations.3 Furthermore, more than 50 additional MC4R single nucleotide polymorphisms have been reported in obese and control individuals, with different effects depending on the population and family studied.4 Recently, obesity-related mutations (A293T, I335S, X361S, I183, A70T and M134I) have been identified in the melanocortin-3 receptor gene (MC3R ) - another energy-regulating melanocortin receptor.5,6 Similar to MC4R polymorphisms, common variants in MC3R show significant frequency variation among different populations, accompanied by controversial effects on obesity phenotypes.5,7
Epidemiological studies8 show that overweight and obesity prevalence rates in South African pupils are fast approaching those observed in developed countries, but the contribution of genetic factors is under-investigated. We analysed MC4R and MC3R for sequence variants that may contribute to the development of obesity in pupils from South Africa.
Methods
We aimed at determining the prevalence of metabolic syndrome and obesity in South African school pupils residing in Cape Town.9 A total of 431 (227 obese-overweight, and 204 normal) individuals of 11 -16 years of age were selected, comprising black and coloured South Africans. The following measurements were performed on all pupils: anthropometric measurements (body weight and height, waist and hip circumference, waist-hip ratio, skinfold thickness etc.). Skinfold thickness was measured at 3 different body sites (subscapular, supra-ileac and upper arm); body mass index (BMI) was calculated as weight per square meter (kg/m2); and 3 readings were taken for blood pressure and waist and hip circumferences. Subjects were classified according to their weight status as obese, overweight and normal using the International Obesity Task Force criteria.10 Blood glucose, lipid levels and blood pressure have previously been described.9
Mutation analysis and genotyping
Genomic DNA was extracted from either whole blood collected in vacutainer ethylenediamine tetra-acetic acid (EDTA) tubes or capillary blood collected onto Whatman FTA Cards (Merck Laboratories, UK). Subjects were screened for MCR3 and MCR4 coding sequence variants by amplifying the coding regions and splice junctions11,12 and subsequent single strand conformation polymorphism (SSCP) analysis. Amplicons with aberrant band patterns were further analysed by automated sequencing using ABI 3730. Subjects were genotyped for the presence of identified sequence variants using the allele-specific restriction enzyme analysis. The C allele of the K6T variant abolished the Mae III restriction enzyme recognition site, while the G allele of the V81I polymorphism created a second restriction recognition site for the BseD 1 enzyme.
Statistical analysis
Our sample size calculation was guided by the following factor: If the 'risk' allele frequency in the general population (controls) is 40%, then 227 cases and 204 controls have 80% power to detect, at a 5% significance level, a 'risk' allele frequency of 52% in cases.
Logistic regression assessed case-control association with genotype and race in joint models. Allelic distributions differed highly significantly between ethnic groups. The differences were such that adjusting trait-gene association analyses for ethnicity would not be valid. All analyses were therefore stratified by ethnicity. We tested association between quantitative traits and genotypes, alleles (additive) and haplotypes for the two polymorphisms. Triglycerides and diastolic and systolic blood pressure were log-transformed prior to analysis, because of skewed distributions. Effect estimates were obtained from linear models. Genotype-association (2 degrees of freedom) was tested by coding genotype as a categorical factor and allelic association (1 degree of freedom) with a numerical variable, counting the number of derived (A) alleles. We inferred haplotypes from the genotypes of the two variants and compared frequencies between obese and control learners. Models were adjusted for age and gender, and replicated measurements were controlled for as random effects.
Data were analysed using the programming language R (www.rproject.org) and specifically packages DGC-genetics (LD, Hardy-Weinberg, genotype and allelic association) and haplo.stats (inferred haplotype association).
Ethical approval was obtained from the Research Ethics Committee of the University of Stellenbosch (Project number: N07/ 07/ 160) and from the Faculty of Health and Wellness Science Ethics Committee, Cape Peninsula University of Technology (Project Reference number: CPUT/ HAS-REC 0016). Written consent was obtained from the parents or guardians of pupils, and oral assent from pupils was obtained on the sampling day.
Results
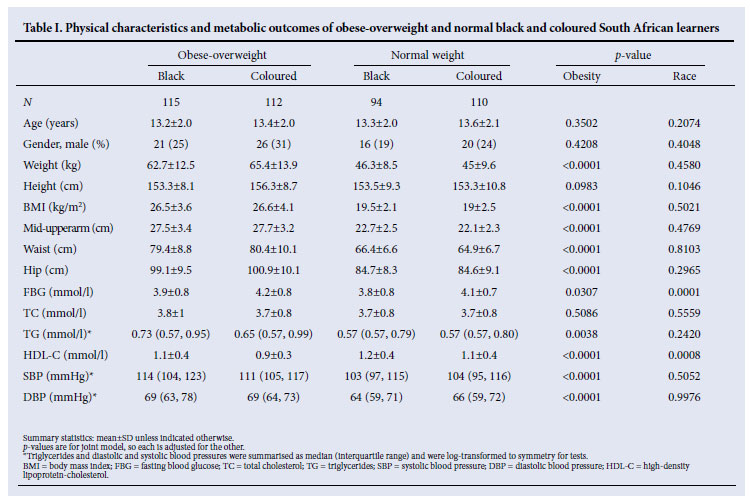
Table I summarises physical characteristics and metabolic outcomes according to ethnicity. By selection, obesity indices were significantly higher in obese-overweight pupils. Analyses were adjusted for age and gender. The fasting blood glucose (FBG) was independently significantly higher in coloured than in black pupils (p=0.0001) and obese-overweight than in normal-weight pupils (p=0.0307). The HDL-C was higher in normal weight than obese-overweight pupils (p=0.0008) and in black compared with coloured pupils (p<0.0001). Triglyceride levels were similar in both racial groups, but significantly higher in obese-overweight pupils (p=0.0038).
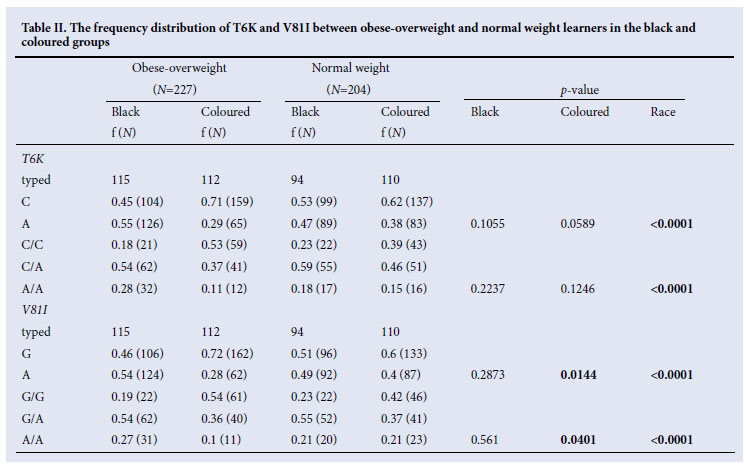
No sequence variants were identified on MC4R , while two common sequence variants (T6K and V81I) were identified in the MC3R coding region. The two coding variants were in strong linkage disequilibrium in all groups, ranging from D'= 0.7437 in coloured normal-weight pupils to D'=0.9264 in the black obese pupils. The frequency distributions (both genotype and allele) differed significantly between the ethnic groups, after adjusting for casecontrol status. The differences were such that potential effects would be cancelled out if the genotype association test were adjusted for ethnicity. Therefore all subsequent analyses were stratified according to ethnicity. The frequency of the V81I A allele was significantly different between coloured overweight-obesity and normal weight pupils (respectively 28% and 40%, p=0.0144) while that of the T6K A allele was only significantly different between the two ethnic groups. The A allele frequency was significantly higher in black pupils (p <0.0001) (Table II). After adjusting for gender and age, both T6K and V81I showed associations with obesity indices (body weight, BMI, hip and mid-upper arm) and total cholesterol, while only the T6K was associated with blood pressure in coloured pupils. In the latter pupils, the estimated effect of the T6K A allele was a decrease of 0.814 cm in mid-upper arm circumference (p=0.0268), 2.873 cm in hip circumference (p=0.0099), 3.225 kg in weight (p=0.0216) and a corresponding 0.959 kg/m2 in BMI (p=0.0476). In addition, the T6K A allele reduced the systolic and diastolic blood pressures, respectively by 2.498 (p=0.0467) and 2.087 mmHg (p=0.0213). In coloured pupils, the V81I A allele decreased total cholesterol by 0.162 mmo/l (p=0.0307), waist circumference by 2.141 cm (p=0.0362), hip circumference by 2.852 cm (p=0.0071), mid-upper arm circumference 0.878 cm (p=0.0119), weight 3.575 kg (p=0.0074) and BMI 1.168 kg/ m2 (p=0.0110).
There was no significant variation in the distribution of the T6K-V81I haplotype in the black group (global p=0.1089) between obeseoverweight and normal weight pupils. Furthermore, no significant association between the T6K-V81I haplotype and physical or metabolic traits were observed in black pupils. However, the C-A was higher in normal weight (5% v. 2%), and A-A haplotype was higher in obese-overweight (52% versus 44%), respectively (p=0.0688) and 0.0749, while the C-G haplotype was most common in the normal weight (48%) rather than in obese-overweight (43%). In coloured pupils, the haplotype distributions differed significantly between obese-overweight and normal pupils (global p=0.0185); C-G was more prevalent in obese-overweight coloured pupils (69% v. 55%, p=0.0063) whereas C-A was more frequent in normal weight pupils (8% v. 2%, p=0.0142). In coloured pupils, after adjusting for age and gender, the C-A haplotype had 2.4 kg/m2 less BMI than the C-G, while the A-A haplotype increased weight by 3.3 kg less. Systolic and diastolic blood pressures were decreased by 10 and 7.5 mmHg respectively in A-G haplotype compared with the C-G haplotype in coloured pupils.
Discussion
Melanocortin 4 and 3 receptors are expressed in neurons of specific areas of the brain that regulate appetite and basal metabolic activity. Melanocortin receptors are part of the leptin-melanocortin pathway that plays a role in glucose homeostasis, insulin secretion, regulation of food intake, and energy balance. Neuropeptides in this pathway regulate food intake and energy balance by binding to melanocortin receptors, particularly MC4R , and to a lesser extent MC3R . Human and animal studies support the role of MC4R in human obesity,3,13 while MC3R variants have recently been implicated as pathogenic mutations that may cause dominantly inherited obesity.5,6 We analysed MC4R and MC3R for sequence variants that may contribute to polygenic obesity in two South African ethnic groups, namely coloured and black populations. Sequence analysis of MC3R in our study groups detected 2 common polymorphisms that showed variability in the distribution pattern between these groups.
The minor A allele frequency of both polymorphisms were higher in black pupils than in coloured, and almost equally distributed in the black population independent of obesity status. In the coloured group, the minor A allele of both polymorphisms in this study had a similar frequency ranging from 28% to 40% independent of obesity status. Compared with American whites,14,15 the homozygosity of these variants in coloured children was not as low. A prevalence of 15.8% of the T6K and V81I MCR3 homozygote minor alleles in African-Americans compared with 1.7% in whites has been reported.14 The variants were associated with several metabolic traits in the coloured group. This association is further reflected in the inferred haplotype analysis. Both variants were associated with obesity indices and total cholesterol, but only T6K was associated with blood pressure in coloured pupils. In the black pupils, T6K was associated with triglyceride only.
Likewise, T6K-V81I haplotype analysis demonstrated a negative association of A-A and C-A haplotypes, compared with the C-G haplotype with indices of obesity, while the A-G was negatively correlated with blood pressure compared with the C-G haplotype in coloured pupils, suggesting a possible protection from obesity and hypertension. The effect posed by the C-A haplotype should, however, be analysed with caution and investigated further as it was observed in only 2% of obese and 5% of normal pupils. Although the minor alleles of both T6K and V81I were more prevalent in black pupils than in coloured, neither allele nor haplotype association tests demonstrated correlation with obesity or other metabolic parameters in the former ethnic group. The A-A haplotype was most common (52%) in obese and overweight pupils whereas the C-G haplotype was the most common (48%) in normal weight black pupils. Similarly, an association between these polymorphisms and body weight in African-American women was not demonstrated.16 Our results contrast with others that demonstrated an association between double homozygosity of these variants and high body weight, with minor allele carriers having greater body fat mass than those carrying the major alleles.5,17 Furthermore, MCR3 knock-out mice had increased fat mass and reduced lean mass, but hyperphagia, while heterozygotes had normal body composition.17
The association between the T6K-V81I and blood pressure (A-G having highly significantly lowered blood pressure than C-G, systolic 10 mmHg, diastolic 7.5 mmHg) in coloured pupils further supports the dual role of MC3R in weight and blood pressure regulation.18 Human and animal studies support the role of MC3R in the development of obesity, but only animal models demonstrated its contribution in blood pressure regulation. Mice lacking MC3R had elevated levels of gamma melanocortin-stimulating hormone (γ-MSH) on both low-and high-salt diet, but developed hypertension on the high-salt diet,19 which suggests that the role of MC3R in blood pressure may only be apparent in the presence of a high salt concentration. MC3R may also play a role in regulation of blood pressure in humans, as epidemiological and functional studies have demonstrated the causative role of obesity in hypertension.20, 21
The MC4R (another melanocortin receptor) has also been implicated in blood pressure regulation. Animal and human studies showed that the defective MC4R confers a relative protective role on the expected increase of blood pressure in obese subjects.22,23 Intracerebroventricular injection of γ-MSH elevated blood pressure and heart rate in wild-type mice but not in obese mice lacking MC4R.22 Obesity increases blood pressure, but a study23 showed that humans with MC4R-deficiency had lower blood pressure than the control group. Blood pressure of control subjects was similar to those of overweight and obese subjects without MC4R defects, suggesting that MC4R -deficient subjects were relatively protected from the expected relationship between overweight and raised blood pressure. In our study, no sequence variants were identified in the MC4R , which is not surprising owing to rarity of polymorphisms implicated in mild obesity in this gene.4 Similarly to our findings, others also failed to detect sequence variants in MC4R .24 According to Jacobson et al., MC4R mutations are likely to occur in populations enriched with severe or early-onset obesity, and therefore are an uncommon cause of obesity in the general population. 24
Blood pressure, central obesity and elevated lipid profile are used to assess metabolic syndrome, which is a multiplex risk factor for cardiovascular diseases and type 2 diabetes. Individuals with lower waist circumference and blood pressure have a lower risk of developing cardiovascular diseases than those with higher values. Although the MC3R variants show a negative effect in blood pressure and weight gain among coloured pupils, the prevalence of metabolic syndrome among black and coloured pupils was not significantly different.9 These findings may therefore be a reflection of gene-gene interaction and/or environmental factors rather than a measured risk of metabolic syndrome and/or cardiovascular diseases in these racial groups.
A limitation of this study was the use of the SSCP method to detect possible sequence variants, which might have contributed to the detection rate. This technique has been reported to detect up to 98% (addition of 15% urea in both 8% and 10% polyacrylamide gels increases the detection limit from 90 to 98%) of nucleotide changes.25 SSCP is also subject to human error regarding interpretation of mobility shifts; and other sequence variants that could have been detected by ABI sequencing might have been missed.
Conclusion
Our results support the role of MC3R variants in weight regulation, with larger effects in the coloured population, in whom T6K and V81I polymorphisms also exact different metabolic effects, supporting the dual role of MC3R in regulating body weight and blood pressure. Although our results demonstrate that the MC3R polymorphisms have a protective effect on these metabolic traits, further study is required to confirm whether this translates into a lower incidence of metabolic syndrome in coloured populations.
This research was supported by a grant from the University Research Fund of the Cape Peninsula University of Technology, Harry Crossley and the National Health Laboratory Services. These funders had no unfluence in this article besides funding the project. We also thank the Department of Education of the Western Cape Provincial Government, participating pupils, and the support staff of the obesity, diabetes and metabolic syndrome research group pupils.
References
1. Comuzzie AG, Allison DB. The search for human obesity genes. Science 1998;280:1374-1377. [ Links ]
2. Hill JO, Peters JC. Environmental contributions to the obesity epidemic. Science 1998;280(5368):1371-1374. [ Links ]
3. Farooqi IS, Keogh JM, Yeo GS, Lank EJ, Cheetham T, O'Rahilly S. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N Engl J Med 2003;348(12):1085-1095. [ Links ]
4. Carroll L, Voisey J, van Daal A. Gene polymorphisms and their effects in the melanocortin system. Peptides 2005;26:1871-1885. [ Links ]
5. Mencarelli M, Walker GE, Maestrini S, et al. Sporadic mutations in melanocortin receptor 3 in morbid obese individuals. Eur J Hum Genet 2008;16(5):581-586. [ Links ]
6. Gotoda T, Scott J, Aitman TJ. Molecular screening of the human melanocortin-4 receptor gene: identification of a missense variant showing no association with obesity, plasma glucose, or insulin. Diabetologia 1997;40:976-979. [ Links ]
7. Lee YS, Poh LK, Kek BL, Loke KY. The role of melanocortin 3 receptor gene in childhood obesity. Diabetes 2007;56(10):2622-2630. [ Links ]
8. Reddy S, Resnicow K, James S, Kambaran N, Omardien R, Mbewu A. Underweight, overweight and obesity among South African adolescents: results of the 2002 National Youth Risk Behaviour Survey. Public Health Nutr 2009;12(2):203-207. [ Links ]
9. Matsha T, Hassan S, Bhata A, et al. Metabolic syndrome in 10-16-year-old learners from the Western Cape, South Africa: Comparison of the NCEP ATP III and IDF criteria. Atherosclerosis 2009;205(2):363-365. [ Links ]
10. Cole TJ, Bellizzi MC, Flegal KM, Dietz WH. Establishing a standard definition for child overweight and obesity worldwide: International survey. Br Med J 2000;320:1240-1251. [ Links ]
11. Vaisse C, Clement K, Durand E. Melanocortin-4 receptor mutations are a frequent and heterogeneous cause of morbid obesity. J Clin Invest 2000;106:253-262. [ Links ]
12. Schalin-Jäntti C, Valli-Jaakola K, Oksanen L, et al. Melanocortin-3-receptor gene variants in morbid obesity. Int J Obes Relat Metab Disord 2003;27(1):70-74. [ Links ]
13. Weide K, Christ N, Moar KM, et al. Hyperphagia, not hypometabolism, causes early onset obesity in melanocortin-4 receptor knockout mice. Physiol Genomics 2003;13(1):47-56. [ Links ]
14. Feng N, Young SF, Aguilera G, et al. Co-occurrence of two partially inactivating polymorphisms of MC3R is associated with pediatric-onset obesity. Diabetes 2005;54:2663-2667. [ Links ]
15. Santoro N, Perrone L, Cirillo G, et al Effect of the melanocortin-3 receptor C17A and G241A variants on weight loss in childhood obesity. Am J Clin Nutr 2007;85:950-953. [ Links ]
16. Li WD, Joo EJ, Furlong EB, et al. Melanocortin 3 receptor (MC3R) gene variants in extremely obese women. Int J Obes Relat Metab Disord 2000;24:206-210. [ Links ]
17. Savastano DM, Tanofsky-Kraff M, Han JC, et al. Energy intake and energy expenditure among children with polymorphisms of the melanocortin-3 receptor. Am J Clin Nutr 2009;90(4):912-920. [ Links ]
18. Butler AA, Cone RD. The melanocortin receptors: lessons from knock out models. Neuropeptides 2002;36:1-8. [ Links ]
19. Ni XP, Pearce D, Butler AA, et al. Genetic disruption of gamma-melanocyte stimulating hormone signaling leads to salt-sensitive hypertension in the mouse. J Clin Invest 2003;111:1251-1258. [ Links ]
20. Oosterom J, Nijenhuis WA, Schaaper WM, et al. Conformation of the core sequence in melanocortin peptides directs selectivity for the melanocortin MC3 and MC4 receptors. J Biol Chem 1999;274:16853-16860. [ Links ]
21. Garrison RJ, Kannel WB, Stokes J III, Castelli WP. Incidence and precursors of hypertension in young adults: the Framingham Offspring Study. Prev Med 1987;16:235-251. [ Links ]
22. Ni XP, Butler AA, Cone RD, Humphreys MH. Central receptors mediating the cardiovascular actions of melanocyte stimulating hormones. J Hypertens 2006;24:2239-2246. [ Links ]
23. Greenfield JR, Miller JW, Keogh JM, et al. Modulation of blood pressure by central melanocortinergic pathways. N Engl J Med 2009;360(1):44-52. [ Links ]
24. Tao YX, Segaloff DL. Functional characterization of melanocortin-3 receptor variants identify a lossof-function mutation involving an amino acid critical for G protein-coupled receptor activation. J Clin Endocrinol Metab 2004;89(8):3936-3942. [ Links ]
25. Nollau P, Wagener C. On behalf of the IFCC Scientific Division, Committee on Molecular Biology Techniques. Methods for detection of point mutations: performance and quality assessment. Clin Chem 1997;43:1114-1128. [ Links ]
Accepted 3 November 2010.
Corresponding author: T Matsha (matshat@cput.ac.za)


{kind=link}
{kind=link}