










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSAMJ: South African Medical Journal
versão On-line ISSN 2078-5135
versão impressa ISSN 0256-9574
SAMJ, S. Afr. med. j. vol.100 no.11 Pretoria Nov. 2010
ORIGINAL ARTICLES
Genetic variations in androgen metabolism genes and associations with prostate cancer in South African men
P FernandezI; P M de BeerII; L van der MerweIV; C F HeynsIII
IPhD. Department of Urology, Stellenbosch University, Tygerberg, W Cape
IIMSc. Department of Urology, Stellenbosch University and Tygerberg Hospital, Tygerberg
IIIMB ChB, MMed (Urol), PhD, FCSSA (Urol). Department of Urology, Stellenbosch University and Tygerberg Hospital, Tygerberg
IVPhD. Biostatistics Unit, Medical Research Council of South Africa, Tygerberg, and Department of Statistics, University of the Western Cape, Bellville, W Cape
ABSTRACT
BACKGROUND: In South Africa white men have the highest incidence of prostate cancer (PCa), coloured (mixed ancestry) men have an intermediate incidence, and low incidences are reported for black and Asian men. It has been suggested that ethnic differences in incidence and mortality of PCa are related to genetic variations in genes that regulate androgen metabolism. We investigated the role of genetic variants in the androgen metabolism genes and the probability of developing PCa in South African coloured and white men.
METHODS: Genotype and allele counts and frequencies of single nucleotide polymorphisms (SNPs) in CYP3A5, CYP3A4 and CYP3A43 were assessed in coloured men (160 case individuals, 146 control individuals) and white men (121 case individuals, 141 control individuals).
RESULTS: A genetic association indicating an increased probability of developing PCa was observed with the G allele of the SNP rs2740574 in CYP3A4 in coloured men, the A allele of rs776746 (CYP3A5) and the G allele of rs2740574 (CYP3A4) in white men, and the G allele of rs2740574 and the C allele of rs501275 (CYP3A43) in the combined ethnic groups analysis. In addition, we identified allele combinations (termed haplotypes) with significantly higher frequencies in the PCa case individuals than in the control individuals.
CONCLUSIONS: The findings support the role of variants in genes that regulate androgen metabolism and the probability of developing PCa. The study paves the way to identify other genetic associations in South African men, and to establish genetic profiles that could be used to determine disease progression and prognosis.
Prostate cancer (PCa) is the second most common histologically diagnosed malignancy in South African men, with an estimated annual incidence rate of 17.8/100 000.1 The age-standardised incidence rate (ASIR) of PCa in South African men in the 55 -59-year age range is 56/100 000; this figure rises to 455/100 000 in men older than 75 years.1 In North America, incidences of PCa are the highest in black men, intermediate in white men and the lowest in Asian men.2 In South Africa, the highest incidence is reported in white men (Northern European ancestry) and the lowest in black and Asian men, with an intermediate incidence reported for coloured (mixed ancestry) men.1 However, substantial under-diagnosis and under-reporting may be the reason for the low incidence in South African black men.3
It has been suggested that ethnic differences in incidence and mortality of PCa are related to genetic variations in genes that regulate androgen metabolism. Studies addressing this hypothesis have demonstrated that genes that regulate androgen metabolism are associated with PCa susceptibility. Findings include genes that encode proteins belonging to the cytochrome P450 (CYP) family (a group of enzymes that are involved in the metabolism of xenobiotics, steroids, vitamins and sex hormones).4 The CYP3A subfamily is a group of enzymes that are key deactivators of testosterone. The CYP3A locus consists of four genes, CYP3A5, CYP3A7, CYP3A4 and CYP3A43, each gene containing 13 exons and located on chromosome 7q21-22.1. We did not screen CYP3A7 because it is predominantly expressed in fetal stages of development4 and we therefore considered it unlikely to play a role in the pathogenesis of PCa.
The CYP3A5 enzyme catalyses the 6 β-hydroxylation of testosterone, producing a less biologically active form of testosterone that is more readily eliminated.4 The single nucleotide polymorphism (SNP) rs776746 (6986A>G; in some publications the A allele = CYP3A5*1 and G allele = CYP3A5*3) in intron 3 creates a cryptic splice site resulting in a messenger ribonucleic acid (mRNA) that is more unstable and more rapidly degraded than the mRNA formed by the wild-type A allele.4 In Finnish men, the rs776746 A allele was significantly associated with PCa bone metastases.5
The CYP3A4 enzyme is involved in the oxidation of testosterone to 2 β-, 6 β- or 15 β-hydroxytestosterone. For the SNP rs2740574 (-392A>G; the A allele = CYP3A4*1A and G allele = CYP3A4*1B) in the 5' regulatory region of the gene, the G allele was initially shown to be associated with PCa in white and black American men,6 which was replicated in subsequent investigations. In black American men, no association was found between the rs2740574 A or G alleles, or other SNP alleles in CYP3A4, and the probability of developing PCa.7
CYP3A43 is expressed predominantly in the prostate and less significantly in the testis, kidney and pancreas.8 The CYP3A43 enzyme is inactive, but splicing of CYP3A43 exon 1 to CYP3A4 and CYP3A5 exons produces hybrid mRNA products, of which the longest CYP3A43/CYP3A4 chimeric isoform can hydroxylate testosterone.8 An association was demonstrated between the G allele of rs680055 (60084G>C [Pro340Ala]) and the probability of developing PCa in white American men with a positive family history of the disease,6 while an association was observed between the G allele and probability of PCa in black American men, after adjusting for age and pack-years of cigarette smoking.9 A significant association was demonstrated between the rs680055 G allele and probability of developing PCa in white American men with a history of benign prostatic hyperplasia (BPH).10 We did not screen rs680055 and chose to screen the SNP rs501275 (75726T>C) in CYP3A43 because we considered it to be more polymorphic in our screening population of coloured and white men.
Little PCa genetic research has been undertaken in South Africa. We have reported associations between SNP alleles in a gene involved in mediating inflammatory responses and the probability of developing PCa in South African men.11 In the present study, we screened genetic variants in the androgen metabolism genes CYP3A5, CYP3A4 and CYP3A43 to determine whether these play a role in the development of PCa and report findings of our case-control genetic association study of self-reported South African coloured and white men.
Methods
Study population
Unrelated men from the South African coloured and white ethnic groups were enrolled in the case-control study to determine genetic factors associated with the probability of developing PCa. The study comprised 160 coloured individuals (mean age 69 (range 47 -88) years) and 121 white individuals (mean age 71 (range 48 -90) years) with histologically confirmed PCa. All were from the Western Cape province of South Africa and had undergone radical prostatectomy, transurethral resection of the prostate or prostatic biopsy in the Department of Urology at Tygerberg Hospital. Control individuals were selected from subjects admitted to the same hospital during the same period and comprised 146 coloured men (mean age 65 (range 52 -91) years) and 141 white men (mean age 67 (range 52 -88) years) from the same geographical region.
Blood samples were collected from each subject. Clinical characteristics, including prostate-specific antigen (PSA), Gleason grade, tumour node metastasis (TNM) stage, age at diagnosis and family history, were obtained from medical records. Controls had PSA levels <2.5 ng/ml and normal findings on digital rectal examination (DRE). Subjects were informed and gave written consent to participate and allow their biological samples to be genetically analysed, according to the Helsinki Declaration. The study was approved by the Faculty of Health Sciences Human Research Ethics Committee, Stellenbosch University.
SNP genotyping
Genomic deoxyribonucleic acid (DNA) was extracted from whole blood using a QIAamp DNA kit (Qiagen, GmbH). The polymerase chain reaction restriction fragment length polymorphism (PCR-RFLP) method used to determine the CYP3A4 rs274057412 and CYP3A5 rs77674613 genotypes has been reported previously. For PCR amplification of the CYP3A43 rs501275 polymorphism we used the primers F 5'-GTCAATGGCAATTTTCTGTT-3' and R 5'-CTGTCTTCACAAACCAGATG-3' in a 50 µl reaction mix containing 50 ng of genomic DNA, 120 ng of each of the forward (F) and reverse (R) primers, 200 µM of dATP, dCTP, dGTP and dTTP, 1.5 mM MgCl2, 1 µl 10 × Taq DNA polymerase buffer, and 0.5 U Taq DNA polymerase. PCR conditions were as follows: 7 minutes at 94ºC; 30 cycles of 30 seconds at 94ºC, 30 seconds at 52ºC, and 45 seconds at 72ºC; and finally 5 minutes at 72ºC. The 210 base pair (bp) PCR amplification product was digested with the restriction enzyme RsaI (Promega, USA) in a volume of 10 µl for 2 hours at 37ºC. The restriction fragments were analysed in a 3% agarose gel stained with ethidium bromide. A fragment at 210 bp corresponds to a homozygous wild-type genotype (TT) and two fragments at 119 bp and 91 bp correspond to the homozygous variant genotype (CC). Three fragments at 210 bp, 119 bp and 91 bp represent the heterozygous genotype (TC).
Statistical analysis
Genotype and allele counts and frequencies were calculated for each SNP. Haplotype frequencies were inferred for a haplotype consisting of allele combinations, one allele from each gene. Each polymorphism was tested in the control individuals to confirm that they were in Hardy-Weinberg equilibrium (HWE). Linkage disequilibrium (LD) (the concept of alleles inherited together at a frequency higher than by chance) between adjacent polymorphisms was assessed. We tested each SNP for an ethnic-genetic interaction for probability of developing PCa. For each SNP in each ethnic group, we tested additive allelic association with PCa, adjusted for age. This provided p-values and estimated allelic odds ratios (ORs), which are the number with which the odds are multiplied, for each additional copy of a specific allele. We adjusted for ethnicity and age in the combined group, and because the South African coloured population is mixed, we corrected for their population stratification by multiplying the obtained p-values by an inflation factor (λ) of 3.06, which we had determined previously in a study on the same group.11 We used the same inflation factor to correct the p-values for the combined ethnic groups, which is very conservative because it is only required for the coloured individuals in the combined group. PCa ORs (and 95% confidence intervals (CIs)) for specific haplotypes compared with the reference (highest frequency) haplotype were estimated, as previously described.11 We used logistic regression models to test the additive allelic association with PCa and estimate the ORs (with corresponding 95% CIs), while adjusting for age by including it as a covariate in the models. Logistic regression was used to test genotype-ethnicity interaction on the probability of developing PCa, and to test haplotype association with PCa. Analyses were done in R, a language and environment for statistical computing available from http://www.R-project.org. The R packages genetics, LDheatmap and haplo.stats were used.
Results
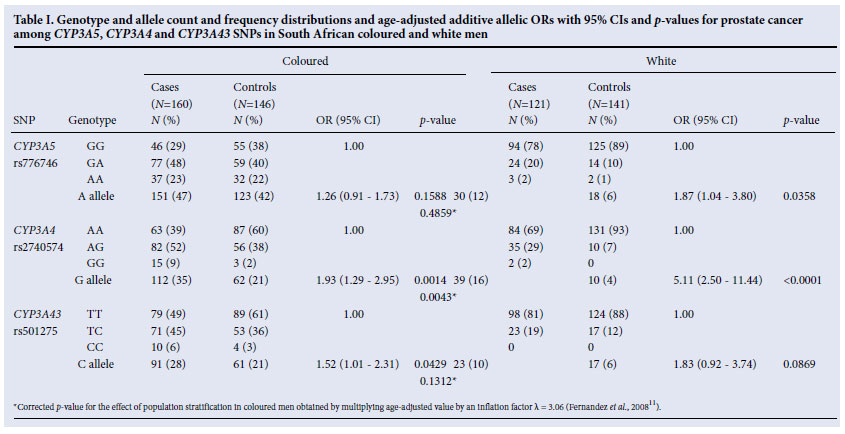
Genotypes were compared within each of the two ethnic groups for rs776746, rs2740574 and rs501275. All three SNPs were in HWE among the controls in both populations (results not shown). For the purpose of this study, the rs776746 A allele was considered the variant allele because it occurred at a lower frequency in both populations, although biologically it produces the wild-type protein product.
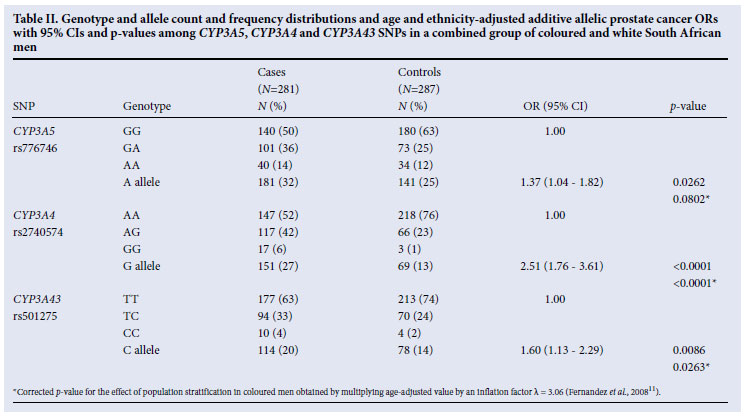
For the coloured group, after adjusting for age, a significant association with PCa was observed for the rs2740574 G allele (OR 1.93; 95% CI 1.29 -2.95; p=0.0014) (each G allele almost doubles the odds of PCa) and the rs501275 C allele (OR 1.52; 95% CI 1.01 -2.31; p=0.0429) (Table I). However, after correcting for population stratification, only the effect of the rs2740574 G allele remained significant (p=0.0043) (Table I). For the white group, after adjusting for age, a significant association with PCa was observed for the rs776746 A allele (OR 1.87; 95% CI 1.04 -3.80; p=0.0358) and rs2740574 G allele (OR 5.11; 95% CI 2.50 -11.44; p<0.0001). There was no significant interaction between ethnicity and genotype on case-control status. We could therefore combine the two ethnic groups, which showed significant associations with PCa with the variant alleles of all three SNPs, although only the association of the rs2740574 G allele and rs501275 C allele remained significant after correcting for the effect of population stratification in the coloured group (Table II). Across the case and control groups in both ethnic groups we observed statistically significant LD between the SNPs (results not shown).
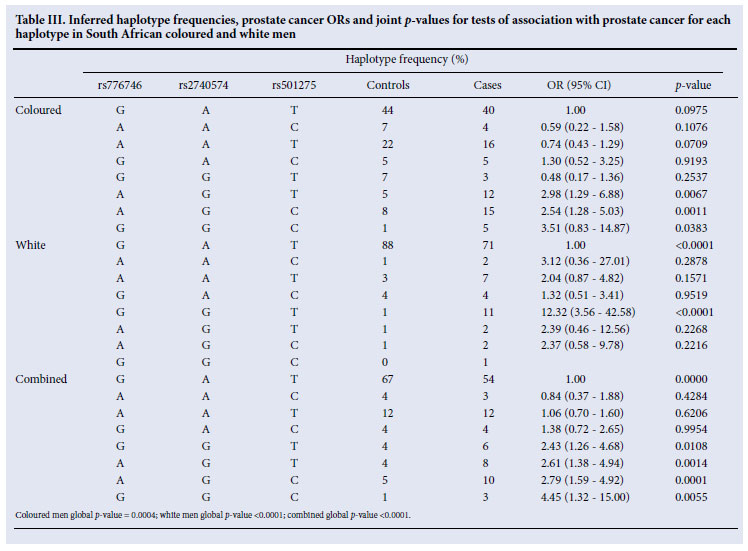
In the white group, a highly significant difference in frequency of the most common haplotype, GAT, was observed between case and control individuals (p<0.0001) (Table III). The inferred frequency was 88% in the control group versus 71% in the case group. In coloured men, we observed the same trend, 44% in the control group and 40% in the case group, but it was not statistically significant (Table III). In the combined ethnic groups, haplotype GAT was also significantly less prevalent in the PCa case individuals (54%) than in the controls (67%) (Table III). In coloured men, haplotypes AGT, AGC and GCC were significantly more prevalent in the case group than in the control group (Table III). In white men, only haplotype GGT showed a highly significant difference, 11% in the case group and 1% in the control group (Table III). For the combined ethnic groups analysis, haplotypes GGT, AGT, AGC and GGC were significantly more prevalent in the case group than in the control group (Table III).
Discussion
Our findings support the involvement of CYP3A5, CYP3A4 and CYP3A43 alleles in the probability of developing PCa in South African men. We report a unique finding indicating increased PCa probability associated with the rs776746 A allele (CYP3A5*1). Additionally, we confirmed an association between the rs2740574 G allele (CYP3A4*1B) and PCa occurrence.6
We demonstrated that the rs776746 A allele was associated with developing PCa in South African white men, whereas in Finnish men the allele was not associated with development of PCa but was associated with bone metastases (disease progression).5 The sample size of our case individuals limited the analytical power to test for associations between the SNP alleles and disease extent. Individuals harbouring at least one copy of the A allele express large amounts of CYP3A5, whereas having a copy of the variant G allele causes alternative splicing and protein truncation resulting in an absence of CYP3A5.4 Men with the A allele would therefore have increased testosterone oxidation, which decreases the bio-availability of testosterone for conversion to 5α-dihydrotestosterone (5α-DHT), the principal androgenic hormone involved in regulating prostate cell growth (more testosterone oxidation = less 5α-DHT = less prostate cell growth). Our association with the A allele contradicts this hypothesis, although it might lend support to this variant being in LD with the causal variant in CYP3A5 or a nearby gene.
The CYP3A4 rs2740574 G allele has been associated with PCa aggressiveness.6 We did not stratify the case individuals by disease aggressiveness because of our small sample size. No substantial effect of the G allele variant on CYP3A4 expression has been identified.14 However, it has been shown in white individuals that the CYP3A4 rs2740574 polymorphism is in tight LD with the CYP3A5 rs776746 polymorphism.4,6,15 Furthermore, it has been suggested that reported associations between the variant G allele and PCa could be due to the polymorphic CYP3A5 expression.15 We observed significant LD between rs776746 and rs2740574, lending support to possible synergistic interaction between their alleles in PCa development in South African men.
Associations have been reported with a specific allele of the CYP3A43 coding SNP rs680055 and PCa, only after stratifying for a positive family history of PCa,6 a history of cigarette smoking9 or a history of BPH.10 We observed a direct association with the CYP3A43 rs501275 C allele and PCa probability only in our combined analysis. It is unclear why the C allele increased probability of the disease, given that CYP3A43 has been suggested to be a pseudogene,4 having a specific role of contributing exons to CYP3A5 and CYP3A4 transcripts to produce hybrid mRNAs.8 Moreover, it is unclear how the rs501275 polymorphism in intron 4 of the gene could play a role in PCa causation as it is probably non-functional. Because the CYP3A locus exhibits significant LD, the association with CYP3A43 might alternatively reflect the effect of another variant in the gene, or in CYP3A5 or CYP3A4.
We report significant differences in haplotype frequency distributions by PCa case-control status, with and without stratifying by ethnicity. The higher frequency of haplotype GAT in the control individuals (Table III) might suggest that this haplotype is protective against developing PCa in white South African men. The CYP3A5-CYP3A4-CYP3A43 haplotypes may contain additional information about disease probability prediction beyond single SNP analysis, suggesting that other variants in these genes or variants in other genes in the region may be involved in the development of PCa.
Limitations of our study include the small sample size, although it was sufficient to detect small to moderate associations. Additionally, the sample size limited our ability to test for associations with disease aggressiveness or disease extent. Cytochromes metabolise xenobiotics and an association with PCa and pack-years of cigarette smoking has been demonstrated.9 Information related to cigarette smoking and alcohol use was occasionally omitted in medical records, and our control individuals were not obliged to indicate cigarette and alcohol use; we were therefore unable to incorporate these data into our analyses. However, future studies with larger sample sizes and collection of all relevant covariates should address these limitations.
Conclusions
We detected variants in genes at the CYP3A locus that are associated with PCa in South African men. Our previous findings support the role of genes involved in inflammation and the development of PCa in South African coloured men.11 This study supports the role of genetic variants in genes that regulate androgen metabolism in the development of PCa in South African men. It has been suggested that prostate tumours initiated under conditions of chronic inflammation are fed by exposure to androgens, resulting in clinically more severe tumours.10 Our study does not conclusively identify a PCa causative variant, although it strongly suggests that the causative variant may lie within or closely adjacent to the CYP3A locus. Further studies in larger sample sizes and in other South African population groups should confirm the associations in CYP3A5, CYP3A4 and CYP3A43, or identify associations in other genes in the region, which will allow stratified analyses by disease aggressiveness. This could lead to uncovering the genetic relationship between genotype and haplotype profiles associated with disease progression and prognosis.
This work was funded by the National Research Foundation (NRF) Thuthuka Programme, the Medical Research Council (MRC) of South Africa and the J H Hayes Fund, but the views and opinions expressed are those of the authors.
References
1. Mqoqi N, Kellett P, Sitas F, Jula M. Incidence of histologically diagnosed cancer in South Africa, 1998-1999. In: National Cancer Registry. Johannesburg: National Cancer Registry of South Africa, 2004. [ Links ]
2. Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ. Cancer statistics, 2009. CA Cancer J Clin 2009;59: 225-249. [ Links ]
3. Heyns CF, Naude AM, Visser AJ, et al. Early diagnosis of prostate cancer in the Western Cape. S Afr Med J 2001; 91: 679-684. [ Links ]
4. Kuehl P, Zhang J, Lin Y, et al. Sequence diversity in CYP3A promoters and characterization of the genetic basis of polymorphic CYP3A5 expression. Nature Genet 2001;27:383-391. [ Links ]
5. Vaarala MH, Mattila H, Ohtonen P, Tammela TLJ, Paavonen TK, Schleutker J. The interaction of CYP3A5 polymorphisms along the androgen metabolism pathway in prostate cancer. Int J Cancer 2008; 122:2511-2516. [ Links ]
6. Zeigler-Johnson C, Friebel T, Walker AH, et al. CYP3A4, CYP3A5, and CYP3A43 genotypes and haplotypes in the etiology and severity of prostate cancer. Cancer Res 2004;64:8461-8467. [ Links ]
7. Sarma AV, Dunn RL, Lange LA, et al. Genetic polymorphisms in CYP17, CYP3A4, CYP19A1, SRD5A2, IGF-1, and IGFBP-3 and prostate cancer risk in African-American men: The Flint Men's Health Study. Prostate 2008;68:296-305. [ Links ]
8. Finta C, Zaphiropoulos PG. Intergenic mRNA molecules resulting from trans-splicing. J Biol Chem 2002;277:5882-5890. [ Links ]
9. Stone A, Ratnasinghe LD, Emerson GL, et al. CYP3A43 Pro(340)Ala polymorphism and prostate cancer risk in African Americans and Caucasians. Cancer Epidemiol Biomarkers Prev 2005;14:1257-1261. [ Links ]
10. Rebbeck TR, Rennert H, Walker AH, et al. Joint effects of inflammation and androgen metabolism on prostate cancer severity. Int J Cancer 2008;123:1385-1389. [ Links ]
11. Fernandez P, de Beer PM, van der Merwe L, Heyns CF. COX-2 promoter polymorphisms and the association with prostate cancer risk in South African men. Carcinogenesis 2008;29:2347-2350. [ Links ]
12. van Schaik RH, de Wildt SN, van Iperen NM, Uitterlinden AG, van den Anker JN, Lindemans J. CYP3A4-V polymorphism detection by PCR-restriction fragment length polymorphism analysis and its allelic frequency among 199 Dutch Caucasians. Clin Chem 2000;46:1834-1836. [ Links ]
13. van Schaik RH, van der Heiden IP, van den Anker JN, Lindemans J. CYP3A5 variant allele frequencies in Dutch Caucasians. Clin Chem 2002;48:1668-1671. [ Links ]
14. Spurdle AB, Goodwin B, Hodgson E, et al. The CYP3A4*1B polymorphism has no functional significance and is not associated with risk of breast or ovarian cancer. Pharmacogenetics 2002;12: 355-366. [ Links ]
15. Wojnowski L, Hustert E, Klein K, et al. Re: modification of clinical presentation of prostate tumors by a novel genetic variant in CYP3A4. J Natl Cancer Inst 2002;94:630-631. [ Links ]
Accepted 4 May 2010.
Corresponding author: P Fernandez (pf3@sun.ac.za)


{kind=link}
{kind=link}
{kind=link}