










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.64 n.1 Pretoria 2022
http://dx.doi.org/10.4102/koedoe.v64i1.1674
REVIEW ARTICLE
Reflecting on research produced after more than 60 years of exclosures in the Kruger National Park
Corli Wigley-CoetseeI, II; Tercia StrydomI, III; Danny GovenderI, IV; Dave I. ThompsonV, VI; Navashni GovenderII, VII; Judith BothaI; Chenay SimmsI; Adolf ManganyiI; Laurence KrugerVIII; Jacques VenterI; Cathy GreaverI; Izak P. SmitI, IX
ISavanna and Grassland Node, Scientific Services, Kruger National Park, Skukuza, South Africa
IISchool of Natural Resource Management, George Campus, Nelson Mandela University, George, South Africa
IIIDepartment of Soil, Crop and Climate Sciences, University of the Free State, Bloemfontein, South Africa
IVDepartment of Paraclinical Sciences, University of Pretoria, Pretoria, South Africa
VSAEON Ndlovu Node, Scientific Services, Kruger National Park, Phalaborwa, South Africa
VISchool of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, Johannesburg, South Africa
VIIConservation Management, Kruger National Park, Skukuza, South Africa
VIIIOrganization for Tropical Studies, Faculty of Science, University of Cape Town, Cape Town, South Africa
IXCentre for African Ecology, School of Animal, Plant and Environmental Sciences, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Herbivores are a main driver of ecosystem patterns and processes in semi-arid savannas, with their effects clearly observed when they are excluded from landscapes. Starting in the 1960s, various herbivore exclosures have been erected in the Kruger National Park (KNP), for research and management purposes. These exclosures vary from very small (1 m2) to relatively large (almost 900 ha), from short-term (single growing season) to long-term (e.g. some of the exclosures were erected more than 60 years ago), and are located on different geologies and across a rainfall gradient. We provide a summary of the history and specifications of various exclosures. This is followed by a systematic overview of mostly peer-reviewed literature resulting from using KNP exclosures as research sites. These 75 articles cover research on soils, vegetation dynamics, herbivore exclusion on other faunal groups and disease. We provide general patterns and mechanisms in a synthesis section, and end with recommendations to increase research outputs and productivity for future exclosure experiments.
CONSERVATION IMPLICATIONS: Herbivore exclosures in the KNP have become global research platforms, that have helped in the training of ecologists, veterinarians and field biologists, and have provided valuable insights into savanna dynamics that would otherwise have been hard to gain. In an age of dwindling conservation funding, we make the case for the value added by exclosures and make recommendations for their continued use as learning tools in complex African savannas
Keywords: disease; fire; herbaceous layer; herbivory; plant productivity; savanna; species diversity.
Introduction
The earliest research exclosures were erected in the Kruger National Park (KNP) in the late 1960s and early 1970s (e.g. Makhohlola exclosure) with the intention 'to provide areas where the grazing effects of animals could be excluded' (Joubert 1986). This was done partly to complement the Experimental Burn Plots (EBPs), a fire-manipulation experiment initiated in the 1950s which focused on studying the effects of different fire seasons and frequencies on vegetation, but where herbivores had free access (see Biggs et al. 2003). However, some of these fenced-off areas (e.g. Hlangwini and N'waswitshumbe exclosures) were also used to support a range of operational management needs, including the breeding of locally rare antelope species (e.g. roan, Hippotragus equinus equinus and tsessebe, Damaliscus lunatus lunatus) and as holding areas for reintroduced white (Ceratotherium simum simum) and black (Diceros bicornis) rhinoceros (e.g. Hlangwini and Black rhino exclosures).
After the detection of widespread tuberculosis in buffalo (Syncerus caffer) in southern KNP, a buffalo breeding enclosure was set up at Satara in 2000 (i.e. Satara [Buffalo] exclosure). Then, in 2002, herbivore exclosures were erected along the Letaba and Sabie rivers after extensive floods removed most of the riparian vegetation (i.e. Letaba and Nkhuhlu exclosures) (O'Keefe & Alard 2002). These exclosures include both full exclusion (i.e. where all but the smallest mammals are excluded) and partial exclusion (excluding only elephant [Loxodonta africana] and, because of their height, giraffe [Giraffa camelopardalis giraffa]), and each divided into burned and unburned sections. The primary purpose here was to understand the effects of herbivore size and the interaction with fire, on vegetation dynamics. A sub-objective was to document the recovery of riparian vegetation after the floods in 2000 (O'Keefe & Alard 2002).
During the early 2000s, several external research programmes which included exclosures in their experiments, were established in KNP. The University of Cape Town (UCT) erected two electrified exclosures, one on the October triennial fire treatment of the Shabeni EBP string at Pretoriuskop and another in the north-west corner of the Satara buffalo exclosure (hereafter UCT Satara exclosure). These exclosures formed part of the broader Tree Grass Programme, with the main objective to investigate hypotheses predicting tree-grass coexistence in savannas (February & Higgins 2010). In addition, several international universities (California Santa Barbara, Colorado State, Florida International, New Mexico, North Carolina Greensboro, Wyoming, and Yale) and local institutions (University of KwaZulu-Natal and South African Environmental Observation Network [SAEON]; hereafter the USA-SA consortium) have, since 2005, used multiple small exclosures near Satara (including on the EBPs and in the Satara (Buffalo) exclosure) to study the interactions between landscape position, fire, rainfall and herbivore size on herbaceous dynamics (Buis et al. 2009; Koerner et al. 2014). Full exclosures exclude all herbivores larger than a hare (Buis et al. 2009) and partial exclosures only allowed access to herbivores ≤ 0.85 m at shoulder height (e.g. impala [Aepyceros melampus] and steenbok [Raphicerus campestris]) (Burkepile et al. 2016).
In 2018, researchers from several institutions (Universities of Florida and Eswatini being prominent), erected an exclosure experiment (called BROWSE, including two full and three partial exclosures) south of Lower Sabie tourist camp, close to the location of the Makhohlola exclosure. This experiment is not reviewed here as the exclosures are relatively new. Apart from these, a host of other external research groups have used small exclosures (i.e. 1 m2), short-term (one growing season) and long-term, to answer various research questions.
Although exclosures can be valuable research sites, they are expensive research infrastructure to install and maintain, especially in areas with high densities of elephants (Grant et al. 2008; Kioko et al. 2008). Although many of the exclosures were initially erected using external funds (e.g. Andrew. W. Mellon Foundation and the National Science Foundation [USA]), South African National Parks (SANParks) assumed responsibility for their maintenance and repair, with the assistance of SAEON in some cases. SANParks, like many other conservation agencies with largely self-generated funding through tourism and with a decreasing government grant, has limited funds to acquire and maintain research infrastructure. Additionally, SANParks is registered as a non-profit parastatal agency, which means operational funds are not guaranteed and cannot be carried over from year to year, thus making long-term budgeting difficult. Many of the exclosures (e.g. UCT Shabeni, Makhohlola, Black rhino exclosure, Capricorn; see Online Appendix 1 or Figure 1 for current status) have seen a steady deterioration of fences or have been decommissioned because of the rising costs of maintaining what is now an ageing infrastructure, in an environment with an increasing population of elephants (Smit et al. 2020). The more recent financial impacts of the global COVID-19 (coronavirus disease 2019) pandemic on tourism specifically, has further exacerbated the situation (Smith et al. 2021), with wide-scale cost curtailment across SANParks.
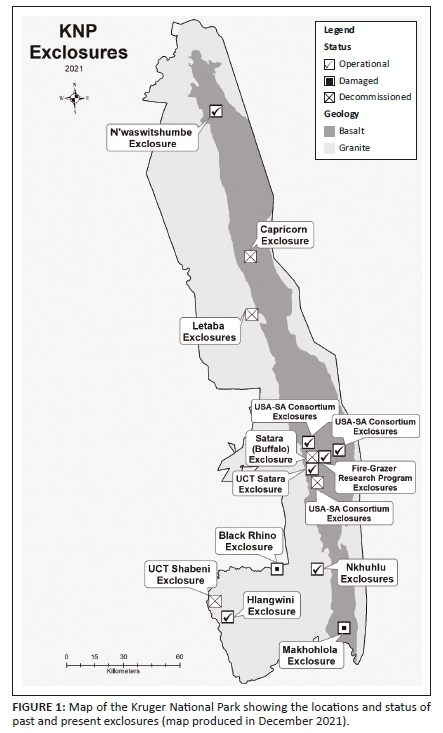
Here, we provide a review of exclosure-based research in KNP. Additionally, we were interested to evaluate the realised impact of these exclosures from the perspective of research output. Research published in peer-reviewed literature was reviewed under five different groupings, based on the history and objectives of the respective exclosures; that is, (1) the exclosures with the longest history (Hlangwini, N'waswitshumbe and Makhohlola), (2) the Nkhuhlu and Letaba exclosures, (3) disease work in the Satara (Buffalo) exclosure, (4) the Tree Grass Programme (UCT Shabeni and UCT Satara), and (5) a grouping capturing all the smaller exclosure work around Satara. Online Appendix 1 provides information about the exclosures, including age, size, experimental design and current status. Furthermore, Online Appendix 2 provides details of the sources used during this review, including the exclosures used and the main variables measured. Considering all these fenced areas as 'herbivore exclosures', even when some of them have enclosed selected herbivores for certain periods, stems from the fact that (1) elephant, an important landscape engineer in the park, was fenced out of structures, (2) herbivore densities in the exclosures were generally lower than areas outside the exclosures, (3) a few of them have been used both as enclosures and exclosures, and (4) fire has been excluded or carefully managed in many of the enclosures and exclosures.
Methods
This review examines the utility of small to large (1 m2 - 870 ha) and temporary (one growing season) to long-term (≥ 15 years) herbivore exclosures in KNP as research infrastructure. To gather all relevant literature, we conducted a Google Scholar database search using keywords including various combinations of 'exclosure OR enclosure #name', 'enclosure KNP' or 'exclosure KNP', as well as the names of the sections of KNP in which the exclosures are located. There are various spellings of the same exclosure name in the literature and all the spelling variations were included in the search. A single standardised spelling for each is used here. We focussed on published work; however, where work was not published, we referred to the corresponding thesis, if available. Work done within exclosures was included regardless of whether the objectives were related directly to herbivory or not. Research on the breeding of rare antelope was not included, as this information is poorly documented in the formal literature. Much of the history and lessons learned are captured in unofficial reports, meeting minutes, correspondence and ranger diaries, but not formally curated (i.e. not listed in the SANParks literature bibliography). Although this history and the associated lessons learned are important and worthy of publication, it requires a dedicated study beyond the scope of the current review.
Review findings
A total of 75 peer-reviewed published works were retrieved during the search which ended on 31 August 2021.
Hlangwini, N'waswitshumbe and Makhohlola exclosures
Eighteen published studies include the Hlangwini (Figure 2), Makhohlola (Figure 3) and N'waswitshumbe (Figures 4 and 5) exclosures. These exclosures were reviewed together as they are often used as replicates in the same studies, and share more or less the same long-term history of low to no herbivore presence. Although some soil research has taken place (e.g. Craine, Morrow & Fierer 2007; Craine, Morrow & Stock 2008; Holdo & Mack 2014; Wigley et al. 2014), the exclosures have mainly been used to compare the effects of herbivore presence or absence on woody vegetation. Complementary work from elsewhere in KNP that used small temporary exclosures to investigate bottlenecks in tree recruitment is included here (Midgley et al. 2020; Namah, Midgley & Kruger 2019). In addition, some studies have investigated the consequences of excluding herbivores for other faunal groups (MacFadyen et al. 2012; McCleery et al. 2018) and they, too, are included here.



Herbivore effects on woody cover
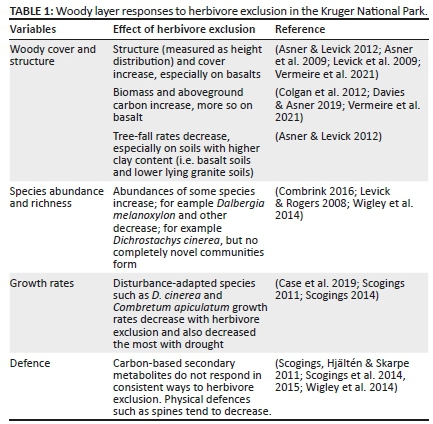
Airborne LiDAR (Light Detection and Ranging) technology has shown that herbivores increase tree-fall rates, decreases woody cover, and alter 3-D structure of woody vegetation and canopy height (Asner et al. 2009; Asner & Levick 2012; Colgan et al. 2012; Levick et al. 2009, 2010) (Figures 3-5). However, the magnitude of the effects is dependent on landscape position and soils (Asner et al. 2009; Colgan et al. 2012). This is apparent in Figure 2, where the tree cover is quite similar inside and outside of the Hlangwini exclosure, situated on nutrient-poor sandy soils, compared to the very pronounced differences visible at Makhohlola and N'waswitshumbe (Figures 3-5), where the soils are much more fertile.
Herbivores impact the abundance of certain species. For instance, Dalbergia melanoxylon, Combretum imberbe, C. hereroense and Lannea schweinfurthii were rarely found outside the exclosures. Alternatively, Dichrostachys cinerea and Albizia harveyii are either more common outside of exclosures or increases faster outside of exclosures over time (Levick & Rogers 2008; Wigley et al. 2014).
Work from elsewhere in KNP using small 1 m2 exclosures showed that the recruitment bottleneck for some tree species such as Kigelia africana and Senegalia (Acacia) nigrescens occurs via predation on seedlings (Namah et al. 2019; Voysey et al. 2021). Even when trees succeed through the recruitment bottleneck, persistent and chronic pruning of plants by elephants and other herbivores can result in mass sterilisation and a change in the adult population demography. From work that included the N'waswitshumbe exclosure, it was shown that most African palms (Hyphaene petersiana) in KNP are prevented from reaching sexual maturity and thus the ability to form seeds, because of chronic elephant herbivory (Midgley et al. 2020).
Herbivore effects on other faunal groups
These exclosures have hardly been used to study the impacts of herbivores on other faunal groups; only four publications were found that addressed it. One study at N'waswitshumbe exclosure found no differences in small mammal species composition in and out of the exclosure (MacFadyen et al. 2012). Small mammal densities were more affected by time since the last veldfire where a drastic reduction in densities were observed post-fire. Alternatively, at the Makhohlola exclosure, which until 2016 supported a far greater biomass of large trees and shrubs than the surrounds, the diversity of small mammals and birds were far greater than the surrounding control plots (McCleery et al. 2018). This work, which included many additional sites, led to the conclusion that the greater density of woody plants when protected from herbivores, results in higher habitat heterogeneity, which supports greater species and functional guild diversity. The mass sterilisation of H. petersiana, mentioned before, may have cascading effects on biodiversity because many species feed on the fruits and flowers (e.g. vervet monkeys, porcupines, squirrels and wasps) and birds would nest within tall individuals (e.g. woodpeckers) (Midgley et al. 2020). We suggest that these cascading effects on biodiversity may be true for any loss of plant species or reduction in vegetation structural diversity with herbivory, although the removal of herbivores may also result in the loss of dispersal opportunities for some species (Midgley, Gallaher & Kruger 2012).
Nkhuhlu and Letaba exclosures
The Nkhuhlu exclosures (full exclosure shown in Figures 6, 7; partial in Figure 8) underpin 26 publications; while the Letaba exclosures were used in 9 articles. The Nkhuhlu exclosures are situated about 25 km away from Skukuza, which is KNP's administrative headquarters and includes a large research camp as well as research support facilities, which likely contributes to its popularity as a research site. The Letaba exclosures on the other hand, are quite remote with an access road that is often inaccessible after rains. The main body of work that has taken place in the Nkhuhlu exclosures addresses the effects of herbivores, and drought on the woody and herbaceous layers.



Soil and plant nutrients
Thorough soil classification was done when the exclosures were erected (Paterson & Steenekamp 2003), but limited follow-up soil work has taken place. Jacobs et al. (2007) investigated nitrogen (N) fixation in Philenoptera violacea in the Nkhuhlu exclosure. They found that P. violacea fixes N and this ability varies with the soil conditions and life history stage; that is, leaves of saplings and juveniles fix higher levels of atmospheric N than mature plants. Although these studies utilised the exclosures as study sites, they did not compare N and phosphorus (P) limitation with and without herbivores or N fixation with and without herbivores. As part of their study on arthropod diversity in the two exclosures, Thoresen et al. (2021) measured a range of soil fertility indicators (including the macro-elements, pH and soil moisture), and found that herbivore exclusion increased both soil nutrients and moisture. However, herbivores had little effect on soil bacterial and fungal communities (Vermeire et al. 2021).
Herbivore effects on the woody layer
The earliest vegetation work from the two exclosures described and mapped their plant communities (Siebert & Eckhardt 2008; Siebert, Siebert & Eckhardt 2010). The woody cover generally increases with herbivore exclusion (Figures 6-8) (Asner & Levick 2012; Asner et al. 2009). These increases are accompanied by increases in woody litter and dead or senescing material, which Asner et al. (2009) suggested could result in more intense fires, where areas are protected from herbivores. Using LiDAR-derived metrics for aboveground carbon (C) pools, Davies and Asner (2019) showed that aboveground C increased over a 6-year period, regardless of herbivore presence, but gains were highest where herbivores were completely excluded.
The woody species composition was slow to react to the exclusion of herbivores at Nkhuhlu, with no change detected after 5 years (Scogings et al. 2012). However, some changes in species abundance were seen after 13 years (Combrink 2016). Similar to what was observed at the N'waswitshumbe and Makhohlola exclosures, Combrink (2016) and Scogings (2011) also found that D. cinerea's abundance increased and growth rates were faster when not protected from herbivores.
Although herbivores may negatively impact woody vegetation survival, severe droughts can be more acute in the short-term. During the 2015-2016 drought, Case et al. (2019) investigated the role of herbivores on tree mortality at Nkhuhlu, and concluded that drought impacts outweighed herbivore effects on tree mortality during below-average rainfall periods. Mortality, however, was most pronounced in the partial exclosure where only elephants and giraffes were excluded, than in the full exclosure. This was in contrast to the average rainfall years where mortality probability was highest in the area open to herbivores, more so than in the two fenced areas (Scogings 2014). It was suggested that this may reflect a combination of meso-herbivore displacement (e.g. impala preferring the partial exclosure to avoid elephants) and herbivore diet-switching; for example work elsewhere showed that mixed feeding impalas utilised a higher proportion of trees during drought than during average rainfall years (Abraham, Hempson & Staver 2019).
Work from KNP exclosures, including Nkhuhlu, found little consistent responses of C-based secondary metabolites (CBSMs) to herbivore exclusion (Scogings, Hjältén & Skarpe 2011, 2013; Scogings et al. 2014, 2015; Wigley et al. 2014). Some species' CBSMs were the highest (e.g. C. apiculatum) with herbivore absence, but responses were species-dependent (Scogings et al. 2013, 2014). For trees in general, the time since first rainfall and season seem to be more reliable predictors of nutrient and tannin concentrations, than the presence of mammal herbivores (Scogings, Hjältén, et al. 2013; Scogings et al. 2013, 2014). In contrast to CBSMs, the presence of herbivores has more consistent effects on structural defences. Intense browsing results in increased shoot growth and branching, and increased spine density and length (Engdahl 2009; Scogings et al. 2013; Wigley et al. 2014).
Herbivore effects on the herbaceous layer
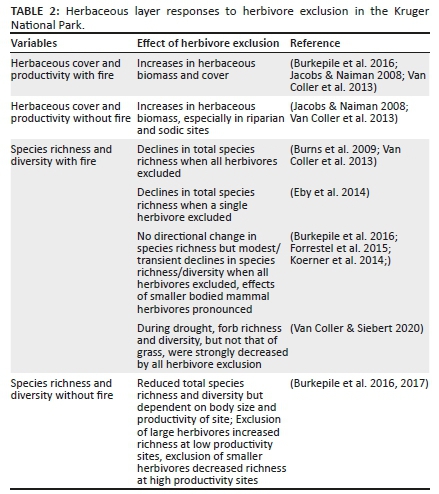
The utilisation of sodic patches is very high when accessible to herbivores at the Nkhuhlu exclosure (Figure 6), resulting in higher N and sodium (Na) and higher N cycling (Alard 2009). Compared to sodic patches, crest patches are relatively water and N limited environments that result in less nutritious grass forage, which is generally less utilised. Nutrient-rich, sodic sites seem to be resilient in the face of high utilisation, with exclusion of herbivores decreasing herbaceous species richness as well as functional diversity (Jacobs & Naiman 2008; Van Coller & Siebert 2015, 2020; Van Coller, Siebert & Siebert 2013). Work from Nkhuhlu exclosure has highlighted the important contribution that forbs make to herbaceous species richness and diversity (e.g. ~80% of herbaceous species richness) (Siebert & Scogings 2015; Van Coller & Siebert 2020), as well as functional richness with many palatable forbs being heavily utilised by herbivores (Siebert & Scogings 2015).
Herbivore effects on other faunal groups
Generally, mammalian herbivores had negative effects on the total invertebrate abundance at the two exclosures (Jonsson et al. 2010; Thoresen et al. 2021), but their presence also resulted in novel species being present (Haba, Herbert & Yang 2019). Depending on landscape position, some groups' abundance (e.g. spiders in riparian areas) increased the most when meso-herbivores were present (i.e. partial exclosure). While arthropod abundance decreased with herbivores presence, its diversity increased (Thoresen et al. 2021). These patterns were related to the indirect effects of herbivores through altering interactions between vegetation, fire, microclimate, and soil nutrients. At Nkhuhlu exclosure, Haba et al. (2019) found an interaction between herbivore presence, S. nigrescens, ants and scale insects. Ants were present with and without herbivores, but Tachardina africana, a scale insect, was only found on S. nigrescens trees where herbivore was present.
Disease work in the Satara buffalo exclosure
Specific-pathogen-free buffalo breeding project
The Satara (Buffalo) exclosure was erected to breed specific-pathogen-free buffalo that could be translocated out of the Foot and Mouth Disease (FMD) endemic area in KNP, and into the FMD-free zone of South Africa. This involved setting up founder herds of Tuberculosis (BTb)-negative and Brucellosis-negative animals caught in northern KNP and transferring them to the Satara (Buffalo) exclosure, to produce calves that would go through a series of quarantine procedures after weaning. The project produced over 400 BTb-, Brucellosis-, FMD- and Corridor disease-free calves that were used to restock Addo Elephant and Mokala National Parks in South Africa and Gorongosa National Park in Mozambique (J. Malan [SANParks], pers. comm., 17 September 2021).
Oregon State University Disease Ecology Programme
From 2015 to 2017, the remaining animals in the Satara exclosure were used in an Oregon State University (OSU) Disease Ecology Research Programme that produced 8 publications and 10 postgraduate qualifications. The unique, longitudinal nature of the OSU study allowed for several diseases to be tracked over time, in individual animals and through the population, revealing temporal patterns not discoverable through typical cross-sectional studies. The Satara exclosure enabled this research, since studying a high number of buffalo, subject to environmental and seasonal disease dynamics, at bi-monthly intervals over 3 years, would otherwise not have been possible. The study was unique in that it used a large social animal in a natural setting; most other epidemiology studies of this kind are laboratory-based. The work helped establish that buffalo could serve as an ideal wild model system, linking individual immunology, environmental change (linked to metabiome health), and parasite transmission dynamics (Combrink et al. 2020; Couch et al. 2017, 2021; Ezenwa et al. 2019; Glidden et al. 2017, 2020; Jolles & Ezenwa 2015; Sisson et al. 2017).
Tree grass programme
The UCT Tree Grass Programme was established in KNP in 2005 and operated for 10 years. The two exclosures (UCT Shabeni and UCT Satara) were part of a broader programme which delivered more than 30 publications and had > 20 postgraduate students involved (E. February [UCT], pers. comm., 03 September 2021). The main objective of the programme was to investigate tree-grass coexistence in terms of two dominant hypotheses, namely, resource-based and disturbance-based. Much of the research that took place in the exclosures addressed the issues around allocation of resources between trees and grasses (February & Higgins 2010; February et al. 2013; Higgins, Keretetse & February 2015; February, Pausch & Higgins 2020).
The results show a high degree of overlap of tree and grass roots, which gives rise to high potential competition (February et al. 2013). Trees had increased growth rates when grasses were removed, but not when they received extra water. However, grass biomass was not influenced by the presence of trees, but was significantly and positively influenced by total precipitation increases, thus suggesting that increasing rainfall facilitates grass competition with trees. Later work by other researchers found that although grasses, saplings and trees used water from the shallower depths when water is not limiting, saplings and trees shifted water uptake to deeper depths when resources are scarce (Kulmatiski & Beard 2013a). Also, small experimental increases in rainfall intensity, without changing the total amount, benefits trees as water is pushed deeper into the soil (Kulmatiski & Beard 2013). If precipitation intensity increases in the future, deep soil water will increase, which may benefit shrubs more than grasses, with associated increases in woody encroachment as a result (Berry & Kulmatiski 2017; Kulmatiski & Beard 2013b).
Smaller-scale exclosures around Satara
During 2005-2006, a USA-SA consortium of researchers initiated a programme with the main objective to broadly test whether general rules can be formulated in terms of drivers, responses and functioning, across grassy ecosystems in North America and South Africa (Knapp et al. 2004, 2006). The group established 38.5 m2 exclosures on (1) the annual, triennial and unburned blocks of the Satara, N'wanetsi and Marheya EBP strings (Figure 9), (2) annual and no-burn blocks inside the Satara (Buffalo) exclosure, and (3) at two sites off the EBPs at mid- and foot-slope catenal positions. In addition, from 2006 to 2008, 1 m2 exclosures were erected on fire treatments to quantify aboveground net primary productivity (ANPP) (Figure 9). The focus of the work evolved from comparing systems across continents to focusing on the single and combined effects of fire, soil fertility (landscape position) and herbivory in African savannas. This research infrastructure has produced 25 publications and supported 17 graduate students and 8 postdoctoral researchers.

Additionally, in 2013, a research group from the Universities of the Witwatersrand (WITS) and Liverpool (hereafter the Fire-Grazer Research Programme) established an experiment north of the N'wanetsi EBP string to test the influence of fire season and fire size on grazers, and the effect of post-burn herbivory on the grass layer (Donaldson et al. 2018). The programme has produced nine publications, and supported seven postgraduate students and two postdoctoral researchers (S. Archibald [WITS], pers. comm., 21 September 2021). For the experiment, 1 m2 exclosures were erected on (1) four replicates each of three fire treatments (unburned, early-burns and late-burns), two fire sizes (0.25 and 5 ha), and three 25 ha late-burn plots, and (2) lawn areas where the grass was generally below 5 cm tall, tussock (bunch) grass areas where the grass was taller than 30 cm and fires were excluded for 3 years, and communities burned at the beginning of the experiment; that is, the end of dry season in 2015. The research of both the USA-SA consortium (now in its 16th year of herbivore exclusion) and the Fire-Grazer Research Programme (8th year of fire manipulation in 2021), is ongoing.
Grazer and fire effects on productivity
The initial research investigated the drivers of annual herbaceous ANPP, using the exclosures on the EBPs and in the Satara (Buffalo) exclosure. Grass and forb ANPP were unaffected by fire regime or whether recent grazing was by a single species (i.e. buffalo) or a mixed suite of species (Buis et al. 2009). Furthermore, Knapp et al. (2012) tested whether overcompensation (i.e. higher ANPP when grazed) took place and showed that regardless of whether areas were burned or not, and regardless of whether herbivory took place by a single species or a diverse suite of species, the total ANPP was most often fully compensated for but not overcompensated. Undercompensated grass ANPP during drier years may be off-set by increased forb ANPP.
The research which took place during drought (either 2015-2016 or experimentally induced) supported emerging patterns elsewhere that show that the herbaceous layer of grasslands and savannas have low resistance but high resilience to drought (Koerner & Collins 2014; Donaldson et al. 2019; Wilcox et al. 2020).Working in full exclosures on the three different fire regimes of the EBPs immediately after the 2015-2016 drought, Wilcox et al. (2020) found that the total ANPP was very resilient to drought and normalised 1 year post-drought when herbivores were excluded. However, responses are affected by the interaction between herbivory and fire. In the absence of herbivores, tussock grasses are generally able to withstand severe drought and maintain cover, and when burned, there is a significant increase in productivity when rainfall returns (Donaldson et al. 2019). Unburned, non-grazed tussock grasses respond poorly to rain. With intense herbivory, however, burned tussock grasses had significant mortality during the drought.
A low severity 3-year drought was experimentally imposed by Koerner and Collins (2014) at plots in an annually burned block of the Satara (Buffalo) exclosure, where herbivory was also excluded for the duration of the experiment, as well as in the UCT Satara exclosure, where herbivory and fire had been excluded since 2002. Twice a year herbivory was simulated by clipping grasses, but not forbs,. With infrequent burning and a history of grazing exclusion (UCT Satara), grass cover and total ANPP were decreased by clipping (i.e. grazing) and drought, as well as by a clipping and drought interaction. With a history of disturbance (i.e. annual burning and low intensity grazing by a single herbivore), there was no effect of clipping or drought on grass cover or total ANPP. In summation, the productivity of the herbaceous layer is resilient in the face of drought, however, the grazing and burning history as well as current precipitation of the area govern responses.
Grazer and fire effects on species richness
Similar to the Nkhuhlu exclosures, the exclusion of herbivores in general decreases herbaceous plant diversity (Burkepile et al. 2017; Burns, Collins & Smith 2009). Eby et al. (2014), working inside the Satara (Buffalo) exclosure, compared plots that excluded buffalo for 7 years with buffalo present, in areas that burned annually and where fire was excluded for 10 years. The removal of buffalo resulted in decreased herbaceous richness and diversity when areas were burned annually, but not when areas were not burned. The increased dominance of one of Themeda triandra, Panicum coloratum or Digitaria eriantha, in annually burned areas with herbivore exclusion, resulted in decreases in richness and diversity (Figure 10).

Although species richness and diversity decreased with herbivore removal during some years on the infrequently burned and unburned areas, there was no definitive change in species or functional community structure after 7 of herbivore removal at any fire frequency (Forrestel, Donoghue & Smith 2015; Koerner et al. 2014). A follow-up study compared species richness and dominance when different sized herbivores were excluded with either full or partial exclosures (Figure 11) (Burkepile et al. 2016). On more frequent burns, exclusion of only larger herbivores (e.g. elephants, zebra [Equus quagga burchellii], wildebeest [Connochaetus taurinus]) increased plant abundance and total cover but not plant diversity, while also excluding smaller species (e.g. impala, warthog [Phacochoerus africanus], steenbok) resulted in modest and transient declines in plant diversity. On unburned areas, herbivore exclusion led to increased dominance and shading, which resulted in declines in plant richness and diversity.
Furthermore, the effect of herbivore exclusion on species richness was shown to depend on site productivity (catenal position) and herbivore size (Burkepile et al. 2017). The exclusion of all herbivores generally increased plant species richness (i.e. forbs and grasses combined) at low productivity, but decreased richness at high productivity (Figure 12). The exclusion effects of only smaller herbivores showed declines in species richness across the productivity gradient. Grass species richness declined regardless of the type of herbivore exclusion or level of productivity, while forb richness depended on productivity and herbivore size, with the largest decreases as a result of the exclusion of smaller herbivores at high productivity.

Herbivore habitat preferences
Both the USA-SA consortium and the Fire-Grazer Research Programme showed that grazers respond positively to areas burned recently, and also prefer more frequently burned areas in general (Burkepile et al. 2013; Donaldson et al. 2018). As a result, fires can be used to create intensely used patches (Donaldson et al. 2018). Fires less than 25 ha attracted sufficient grazers to shorten grass height, but only for one growing season. The repetition of late burns (i.e. September/October) for an additional 2 years resulted in the active selection of these areas by wildebeest and impala, and subsequently, grazing pressure was high enough to initiate positive feedbacks and maintain lawns that excluded repeat fires. When grazing was excluded, grasses quickly recovered to pre-fire levels by the end of the season.
Herbivore effects on grass nutrition
The results from research that included the Satara (Buffalo) exclosure as a single grazer site with less intense utilisation, found that grasses are higher in N and P in areas with a combination of wildlife species present, as opposed to areas with only a single species present (Baumgartner et al. 2015; Treydte et al. 2013). These results reflect in part a switch in the dominant grass species when a diverse suite of wildlife species is present. Urochloa mosambicensis, for instance, which was common in the mixed wildlife sites, is higher in N and P when compared to D. eriantha, which was common in the single species sites. However, the same grass species were also more nutrient rich in the mixed wildlife sites, as opposed to the single herbivore species site, which may reflect a positive feedback between being frequently grazed and increased nutrient content.
Synthesis
Within the context of a large protected area such as KNP, the herbivore exclusion sites enable intensive study of one of the main drivers in disturbance-driven savanna systems, that is herbivory by mammalian herbivores. They allow the partitioning of effects of those disturbances that managers can partly control (e.g. fire regimes, herbivore densities and disease management) from those that cannot be manipulated easily (e.g. droughts, floods and atmospheric CO2 concentrations). The understanding and knowledge that result from this research inform management decisions and policies, allowing SANParks to learn and adapt in an ever-changing environment.
Realising the value of exclosures as research infrastructure in KNP has been varied. The largest exclosures, which also have the longest histories, have been relatively underutilised as research sites (e.g. Hlangwini and N'waswitshumbe). This may be partly because of the lack of replicated treatments and the problem of pseudoreplication (Holdo & Mack 2014), the relatively remote location of these exclosures, and the lack of dedicated research funding within SANParks. The more successful exclosures, from a research output perspective, are those that have been erected through fully-funded research programmes that include dedicated personnel and bursaries for students, with specific objectives in mind, and where logistics such as distance to laboratories and research accommodation have been taken into account.
The two areas of savanna ecology in which a significant body of work has been produced, across multiple exclosures in KNP, are the woody layer and herbaceous layer dynamics. Most of the woody work was done in the Nkhuhlu exclosure, with additional work from the Hlangwini, N'waswitshumbe and Makhohlola exclosures. Multiple studies from the Nkhuhlu exclosure and the Satara-based programmes focused on the interactions between herbivores and fire on the herbaceous layer, with some additional work addressing potential competition between trees and grasses.
Woody layer
It is clear that herbivory decreases woody cover and simplifies vegetation structure (Table 1, Figures 3-8). The effects on woody species richness is less clear; unique species occur with or without herbivory, but the most pronounced effects of herbivores are changing abundances, rather than creating unique woody communities. For instance, certain species thrive when faced with disturbance by herbivores (and fire) and decrease when protected from herbivory. The KNP subscribes to the assumption that increased heterogeneity (e.g. a range of woody cover states ranging from open to dense areas) will increase resilience and biodiversity (see Rogers 2003; Smit & Prins 2015). More open savanna is often more productive in terms of herbaceous productivity and the herbivore assemblage may change along a woody cover gradient (Smit & Prins 2015). Woodlands of intermediate densities harbour high faunal biodiversity, but this may be reversed at high levels of woody densities (McCleery et al. 2018; Stanton et al. 2018). The implication is that a range of herbivore densities should ensure a range of woody densities, which in turn will maximise biodiversity.
Herbaceous layer
Herbaceous productivity is mostly maintained with high herbivory and frequent fire, and is only impacted during extreme droughts, but recovers rapidly post-drought (Buis et al. 2009; Knapp et al. 2012). Species diversity also depends on the presence of herbivory and often decreases when herbivores are excluded (Burkepile et al. 2016; Eby et al. 2014; Van Coller et al. 2013, 2018). There seems to be an intermediate level of herbaceous biomass that ensures highest herbaceous species richness and diversity, and which is maintained by herbivore presence (Van Coller & Siebert 2015). However, grazing and fire have very important interactive effects on herbaceous dynamics; herbaceous richness is the highest in the absence of fire and declined more with herbivore exclusion than when burned (Smith et al. 2016a).
The initial dominance of B. radicans compared to more palatable grasses often governs responses to herbivore removal (Eby et al. 2014; Forrestel et al. 2015). Low levels of herbivory (such as in Satara (Buffalo) exclosure) or herbivory exclusion (such as UCT Satara) give rise to herbaceous layers dominated by tall, palatable grasses (Figure 10). High levels of herbivory and fire on the other hand give rise to the dominance of Bothriochloa radicans. When palatable grasses are dominant, the removal of herbivory results in palatable grasses being stimulated when burned, and the increase in the dominance of these in turn result in lower species richness (Eby et al. 2014). On the EBPs however, B. radicans is dominant in the more frequently burned plots. As B. radicans is mostly avoided by herbivores, herbivore exclusion results in little overall change in species richness because B. radicans remains the dominant species. Even though it is difficult to decrease the dominance of B. radicans through shorter-term fire or herbivore exclusion, B. radicans was greatly reduced during the recent drought, with drought-recovery seeing an increase in palatable grasses (Swemmer et al. 2018; Wigley-Coetsee & Staver 2020).
Apart from an interaction between fire regime, grazer exclusion and herbaceous dynamics, effects of herbivore exclusion depend on the types and sizes of animals and their interactions with different fire regimes (Burkepile et al. 2016), herbivore densities (Burkepile et al. 2017; Treydte et al. 2013), and site productivity (Burkepile et al. 2017). Another important consideration is the balance of grazer, mixed feeder and browsing herbivores (Burkepile et al. 2017; Burns et al. 2009; Treydte et al. 2013). In systems where the removal of herbivores results in pronounced directional loss of herbaceous species richness over time, the herbivores often belong to one type, for example only grazers (Eby et al. 2014; Smith et al. 2016b). The removal of smaller herbivores, which include the mixed-feeding impala, has the most conspicuous impacts; for example lowered species richness and diversity, increased plant dominance and increased plant litter (Burkepile et al. 2016, 2017). This may be a result of both their numerical dominance in KNP as well as their considerable utilisation of the forb layer. A review of herbivore exclusion effects on the herbaceous as well as woody layers across African savannas (which included data from the KNP exclosures) also showed that herbivore impacts are dominated by small-bodied mixed feeders, which was mostly because of their high population densities (Staver et al. 2021).
Lastly, little work has compared mammalian herbivory with invertebrate herbivory impacts. The only study on this topic compared termite herbivory when mammals were excluded, and found that although both types of herbivory were more intense on termite mounds than off, termite herbivory removed more biomass than mammal herbivores during the wet season (Davies et al. 2016).
Given the experimental design and obvious effects of herbivory on savanna systems, exclosures have provided excellent learning opportunities for students across the field of education. We have highlighted the number of graduate students who were involved in exclosure research in KNP. Yet, these exclosures have also played a major role in undergraduate training. For example, the Organization of Tropical Studies (OTS) has been working in KNP since 2004 and has conducted student projects in multiple exclosures across the park. To date about 35 undergraduate student research projects from OTS have used SANParks exclosures (Unpublished data; OTS) to understand the effect of mega-herbivores on woody plant demography, changes in community composition of woody plants and grasses, the consequences of mutualism breakdown for plant demography, the effect of invertebrate versus mammalian herbivory on termite mounds, phenotypic variation in physical defences of woody plants and the influence of vegetation structure on invertebrate (butterflies and dung beetles) and small vertebrate communities. Many of these smaller research projects served as pilot projects for graduate studies and some of these have been published (e.g. Haba et al. 2019; Midgley et al. 2020; Namah et al. 2019). Multiple local and foreign universities, including UCT, WITS, North West University, Bayreuth (Germany), University of Florida (USA), and Western Sydney University (Australia), currently utilise exclosures as part of their field-based experiential training. Importantly, the exclosures create opportunities that would not otherwise be available. For instance, studying disease ecology and eco-immunology in large ungulates in a natural system with a diversity of large dangerous animals is both expensive and logistically challenging. The Satara (Buffalo) exclosure created a tractable and unique, longitudinal study system which has allowed training in a fairly safe environment.
Implications and recommendations for establishing exclosures
The lessons learned from setting up and managing exclosures, and increasing the research uptake thereof, include the following:
-
Give consideration to logistics such as ease of access, including distance from research camps and presence of all-weather access roads in selecting exclosure sites.
-
Ensure a budget and maintenance plan that stretches beyond the establishment of the exclosure, and which is linked to the time frame for which the exclosure is needed (e.g. 20 years). Build collaborations and joint ownership of exclosures with land managers during the planning phase.
-
Purposefully design the exclosure experiment with research questions in mind, for example variable fire treatments, across catenal positions, allowing selective exclusion of different-size herbivores.
-
Adapt the size of the exclosures to fit the research questions asked; for example the USA-SA consortium exclosures have been very productive in terms of research outputs. Although these exclosures cover a relatively small area, they are numerous and large enough to capture dynamic herbaceous layer changes.
-
Employing 'clever' design may save much money in the long-term; for instance, electrified 2- or 3-strand wire exclosures with electrified wires positioned horizontally up to 1 m, such as those used at Mpala in Kenya and Grumeti in Tanzania (A. Uys [Maritaba], pers. comm. 20 September 2021), are cost effective and extremely efficient as a barrier against megafauna movement. Full exclosures inside partial exclosures allow for some level of protection of the full exclosure against elephant damage and can decrease overall costs of maintenance.
-
Canvas the broader research community to solicit baseline information which will add value to the exclosure and stimulate further research (e.g. floristic composition, pre-treatment conditions, aerial photographic and LiDAR surveys, etc.). Make baseline information available as a 'starter pack' for researchers and students - this can include information pertaining to the year of establishment, size, spatially-explicit treatment history (e.g. fire treatments), long-term average and yearly rainfall, estimates of herbivore density outside exclosures, and so on. Collaboration beyond the initial group and objectives have made exclosures such as the USA-SA consortium exclosures very productive and maintains high energy and interest regarding the exclosures.
-
Prepare a closure or hand-over plan for the exclosure, which may include a review of research (i.e. knowledge generated, publications, students graduated, etc.), and a long-term maintenance plan including costing. Do not assume that the land manager or owner will be in the position to maintain ageing infrastructure beyond the lifespan of the programme.
Conclusion
Fence-line contrasts between exclosures and the surrounding landscape are very effective in demonstrating the significant effects and interactions of herbivores in savanna ecosystems to both managers and the public alike, and for stimulating new research ideas for scientists. In addition, exclosures are excellent outdoor classrooms and laboratories for training the next generation of ecologists. With a renewed interest in the implications of the mass extinction of mammal herbivores (e.g. Rowan & Faith 2019) and 'rewilding' as an ecological restorative mechanism in response, exclosures are one of the critical tools to explore these concepts (see e.g. two recent Special Issues on mega-herbivore extinction and rewilding by Malhi et al. 2016; Smith et al. 2016a).
The loss of exclosure infrastructure in KNP represents a missed opportunity, in that it may form a global research platform similar to the EBPs. This fire experiment has been running uninterrupted for seven decades and have become one of the primary sites for studying fire effects globally (e.g. Pellegrini et al. 2021). These types of long-term research infrastructure and associated data that outlive the typical 3-5 year funding and project cycles are some of the main reasons researchers choose to conduct research in KNP. As a result, KNP is the premier savanna research destination in Africa (Smit et al. 2017). Lastly, some of the long-term exclosure work from KNP reminds us not only how resilient savannas are in the face of disturbance, but also how they are shaped by disturbances, realising outcomes which are difficult to achieve by management alone.
Acknowledgements
The authors would like to thank all the South African National Parks (SANParks) staff including section rangers, research assistants and technicians who have worked so hard for many years to service, maintain, and survey exclosures in the KNP. The authors also wish to thank Dr. Anthony Swemmer and South African Environment Observation Network (SAEON) for all the support, both logistically and financially, in maintaining and surveying some of the exclosures, especially the Letaba exclosure.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
C.W-C., T.S., D.G., D.I.T., L.K. and I.P.S. contributed to the text; C.W-C., T.S., C.G., N.G., D.G. and I.P.S. conceptualised the work; J.B., A.M., L.K., and J.V. provided stats on exclosure maintenance; C.W-C. and T.S. analysed literature and data and C.S. provided the map.
Ethical considerations
This article followed all ethical standards for research without direct contact with human or animal subjects.
Funding information
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.
Data availability
All data, in this case works of literature reviewed have been summarised in Online Appendix 2.
Disclaimer
The views and opinions expressed in this article are those of the authors and do not necessarily reflect the official policy or position of any affiliated agency of the authors.
References
Abraham, J.O., Hempson, G.P. & Staver, A.C., 2019, 'Drought-response strategies of savanna herbivores', Ecology and Evolution 9(12), 7074-7056. https://doi.org/10.1002/ece3.5270 [ Links ]
Alard, G.F., 2009, 'A study of selected grass ecophysiological characteristics as well as a comparison of the quality and quantity of grass in sodic and crest patches within a semi-arid granitic savanna catena in the southern KNP, South Africa', PhD thesis, University of the Witwatersrand, Johannesburg. [ Links ]
Asner, G.P. & Levick, S.R., 2012, 'Landscape-scale effects of herbivores on treefall in African savannas', Ecology Letters 15(11), 1211-1217. https://doi.org/10.1111/j.1461-0248.2012.01842.x [ Links ]
Asner, G.P., Levick, S.R., Kennedy-Bowdoin, T., Knapp, D.E., Emerson, R., Jacobson, J. et al., 2009, 'Large-scale impacts of herbivores on the structural diversity of African savannas', Proceedings of the National Academy of Sciences 106(12), 4947-4952. https://doi.org/10.1073/pnas.0810637106 [ Links ]
Baumgartner, S.A., Treydte, A.C., Grant, C.C. & Van Rooyen, J., 2015, 'Can diverse herbivore communities increase landscape heterogeneity? Comparing wild and domestic herbivore assemblages in a South African savanna', Perspectives in Plant Ecology, Evolution and Systematics 17(1), 34-43. https://doi.org/10.1016/j.ppees.2014.11.002 [ Links ]
Berry, R.S. & Kulmatiski, A., 2017, 'A savanna response to precipitation intensity', PloS one 12(4), e0175402. https://doi.org/10.1371/journal.pone.0175402 [ Links ]
Biggs, R., Biggs, H.C., Dunne, T.T., Govender, N. & Potgieter, A.L.F., 2003, 'Experimental burn plot trial in the Kruger National Park: History, experimental design and suggestions for data analysis', Koedoe 46(1), 1-15. https://doi.org/10.4102/koedoe.v46i1.35 [ Links ]
Buis, G.M., Blair, J.M., Burkepile, D.E., Burns, C.E., Chamberlain, A.J., Chapman, P.L. et al., 2009, 'Controls of aboveground net primary production in mesic savanna grasslands: An inter-hemispheric comparison', Ecosystems 12(6), 982-995. https://doi.org/10.1007/s10021-009-9273-1 [ Links ]
Burkepile, D.E., Burns, C.E., Tambling, C.J., Amendola, E., Buis, G.M., Govender, N. et al., 2013, 'Habitat selection by large herbivores in a southern African savanna: The relative roles of bottom-up and top-down forces', Ecosphere 4(11), 1-19. https://doi.org/10.1890/ES13-00078.1 [ Links ]
Burkepile, D.E., Fynn, R.W., Thompson, D.I., Lemoine, N.P., Koerner, S.E., Eby, S. et al., 2017, 'Herbivore size matters for productivity-richness relationships in African savannas', Journal of Ecology 105(3), 674-686. https://doi.org/10.1111/1365-2745.12714 [ Links ]
Burkepile, D.E., Thompson, D.I., Fynn, R.W., Koerner, S.E., Eby, S., Govender, N. et al., 2016, 'Fire frequency drives habitat selection by a diverse herbivore guild impacting top-down control of plant communities in an African savanna', Oikos 125(11), 1636-1646. https://doi.org/10.1111/oik.02987 [ Links ]
Burns, C.E., Collins, S.L. & Smith, M.D., 2009, 'Plant community response to loss of large herbivores: Comparing consequences in a South African and a North American grassland', Biodiversity and Conservation 18(9), 2327-2342. https://doi.org/10.1007/s10531-009-9590-x [ Links ]
Case, M.F., Wigley-Coetsee, C., Nzima, N., Scogings, P.F. & Staver, A.C., 2019, 'Severe drought limits trees in a semi-arid savanna', Ecology 100(11), e02842. https://doi.org/10.1002/ecy.2842 [ Links ]
Colgan, M.S., Asner, G.P., Levick, S.R., Martin, R.E. & Chadwick, O.A., 2012, 'Topo-edaphic controls over woody plant biomass in South African savannas', Biogeosciences 9(5), 1809-1821. https://doi.org/10.5194/bg-9-1809-2012 [ Links ]
Combrink, L., 2016, 'Savanna woody regeneration in response to different treatments of herbivory and fire', PhD thesis, North-West University (South Africa), Potchefstroom Campus. [ Links ]
Combrink, L., Glidden, C.K., Beechler, B.R., Charleston, B., Koehler, A.V., Sisson, D. et al., 2020, 'Age of first infection across a range of parasite taxa in a wild mammalian population', Biology Letters 16(2), 20190811. https://doi.org/10.1098/rsbl.2019.0811 [ Links ]
Couch, C.E., Movius, M.A., Jolles, A.E., Gorman, M.E., Rigas, J.D. & Beechler, B.R., 2017, 'Serum biochemistry panels in African buffalo: Defining reference intervals and assessing variability across season, age and sex', PLoS One 12(5), e0176830. https://doi.org/10.1371/journal.pone.0176830 [ Links ]
Couch, C.E., Stagaman, K., Spaan, R.S., Combrink, H.J., Sharpton, T.J., Beechler, B.R. et al., 2021, 'Diet and gut microbiome enterotype are associated at the population level in African buffalo', Nature Communications 12(1), 1-11. https://doi.org/10.1038/s41467-021-22510-8 [ Links ]
Craine, J.M., Morrow, C. & Fierer, N., 2007, 'Microbial nitrogen limitation increases decomposition', Ecology 88(8), 2105-2113. https://doi.org/10.1890/06-1847.1 [ Links ]
Craine, J.M., Morrow, C. & Stock, W.D., 2008, 'Nutrient concentration ratios and co-limitation in South African grasslands', New Phytologist 179(3), 829-836. https://doi.org/10.1111/j.1469-8137.2008.02513.x [ Links ]
Davies, A.B. & Asner, G.P., 2019, 'Elephants limit aboveground carbon gains in African savannas', Global Change Biology 25(4), 1368-1382. https://doi.org/10.1111/gcb.14585 [ Links ]
Davies, A.B., Van Rensburg, B.J., Robertson, M.P., Levick, S.R., Asner, G.P. & Parr, C.L., 2016, 'Seasonal variation in the relative dominance of herbivore guilds in an African savanna', Ecology 97(6), 1618-1624. https://doi.org/10.1111/gcb.14585 [ Links ]
Donaldson, J.E., Archibald, S., Govender, N., Pollard, D., Luhdo, Z. & Parr, C.L., 2018, 'Ecological engineering through fire-herbivory feedbacks drives the formation of savanna grazing lawns', Journal of Applied Ecology 55(1), 225-235. https://doi.org/10.1111/1365-2664.12956 [ Links ]
Donaldson, J.E., Parr, C.L., Mangena, E.H. & Archibald, S., 2019, 'Droughts decouple African savanna grazers from their preferred forage with consequences for grassland productivity', Ecosystems, 23, 689-701. https://doi.org/10.1007/s10021-019-00438-x [ Links ]
Eby, S., Burkepile, D.E., Fynn, R.W., Burns, C.E., Govender, N., Hagenah, N. et al., 2014, 'Loss of a large grazer impacts savanna grassland plant communities similarly in North America and South Africa', Oecologia 175(1), 293-303. https://doi.org/10.1007/s00442-014-2895-9 [ Links ]
Engdahl, F., 2009, 'Herbivory on woody plants and induced responses in two similar species of Acacia in the Kruger National Park, South Africa', PhD thesis, Wildlife, Fish, and Environmental Studies, Swedish University of Agricultural Sciences. [ Links ]
Ezenwa, V.O., Jolles, A.E., Beechler, B.R., Budischak, S.A. & Gorsich, E.E., 2019, 'The causes and consequences of parasite interactions: African buffalo as a case study', in K. Wilson, A. Fenton & D. Tompkins (eds.), Wildlife Disease Ecology: Linking Theory to Data and Application, pp. 129-160, Cambridge University Press, Cambridge. [ Links ]
February, E., Pausch, J. & Higgins, S.I., 2020, 'Major contribution of grass roots to soil carbon pools and CO 2 fluxes in a mesic savanna', Plant and Soil 454, 207-215. https://doi.org/10.1007/s11104-020-04649-3 [ Links ]
February, E.C. & Higgins, S.I., 2010, 'The distribution of tree and grass roots in savannas in relation to soil nitrogen and water', South African Journal of Botany 76(3), 517-523. https://doi.org/10.1016/j.sajb.2010.04.001 [ Links ]
February, E.C., Higgins, S.I., Bond, W.J. & Swemmer, L., 2013, 'Influence of competition and rainfall manipulation on the growth responses of savanna trees and grasses', Ecology 94(5), 1155-1164. https://doi.org/10.1890/12-0540.1 [ Links ]
Forrestel, E.J., Donoghue, M.J. & Smith, M.D., 2015, 'Functional differences between dominant grasses drive divergent responses to large herbivore loss in mesic savanna grasslands of North America and South Africa', Journal of Ecology 103(3), 714-724. https://doi.org/10.1111/1365-2745.12376 [ Links ]
Glidden, C., Beechler, B., Buss, P.E., Charleston, B., De Klerk-Lorist, L.-M., Maree, F.F. et al., 2017, 'Detection of pathogen exposure in African buffalo using non-specific markers of inflammation', Frontiers in Immunology 8, 1944. https://doi.org/10.3389/fimmu.2017.01944 [ Links ]
Glidden, C.K., Koehler, A.V., Hall, R.S., Saeed, M.A., Coppo, M., Beechler, B.R. et al., 2020, 'Elucidating cryptic dynamics of Theileria communities in African buffalo using a high-throughput sequencing informatics approach', Ecology and Evolution 10(1), 70-80. https://doi.org/10.1002/ece3.5758 [ Links ]
Grant, C.C., Bengis, R., Balfour, D., Peel, M.,Davies-Mostert, W., Killian, H.,Little, R., Smit, I., Garai, M. & Henley, M., 2008, 'Controlling the distribution of elephants', in R.J Scholes & K.G Mennell (eds.), Elephant Management: A Scientific Assessment for South Africa, pp. 329-369, Wits University Press, Johannesburg. [ Links ]
Haba, Y., Herbert, R. & Yang, L., 2019, 'An undescribed relationship between the knobthorn tree (Senegalia nigrescens), ants, and a lac scale insect in South Africa', African Journal of Ecology 57(4), 595-598. https://doi.org/10.1111/aje.12627 [ Links ]
Higgins, S.I., Keretetse, M. & February, E.C., 2015, 'Feedback of trees on nitrogen mineralization to restrict the advance of trees in C4 savannahs', Biology Letters 11(8), 20150572. https://doi.org/10.1098/rsbl.2015.0572 [ Links ]
Holdo, R.M. & Mack, M.C., 2014, 'Functional attributes of savanna soils: Contrasting effects of tree canopies and herbivores on bulk density, nutrients and moisture dynamics', Journal of Ecology 102(5), 1171-1182. https://doi.org/10.1111/1365-2745.12290 [ Links ]
Jacobs, S.M. & Naiman, R.J., 2008, 'Large African herbivores decrease herbaceous plant biomass while increasing plant species richness in a semi-arid savanna toposequence', Journal of Arid Environments 72(6), 891-903. https://doi.org/10.1016/j.jaridenv.2007.11.015 [ Links ]
Jacobs, S.M., Pettit, N.E. & Naiman, R.J., 2007, 'Nitrogen fixation by the savanna tree Philenoptera violacea (Klotzsch) Schrire (Apple leaf) of different ages in a semi-arid riparian landscape', South African Journal of Botany 73(1), 163-167. https://doi.org/10.1016/j.sajb.2006.09.001 [ Links ]
Jolles, A.E. & Ezenwa, V.O., 2015, 'Ungulates as model systems for the study of disease processes in natural populations', Journal of Mammalogy 96(1), 4-15. https://doi.org/10.1093/jmammal/gyu007 [ Links ]
Jonsson, M., Bell, D., Hjältén, J., Rooke, T. & Scogings, P.F., 2010, 'Do mammalian herbivores influence invertebrate communities via changes in the vegetation? Results from a preliminary survey in Kruger National Park, South Africa', African Journal of Range & Forage Science 27(1), 39-44. https://doi.org/10.2989/10220111003703468 [ Links ]
Joubert, S.C.J., 1986, 'Masterplan for the management of Kruger National Park', Vol 1-6. Unpublished document, Skukuza Library Archives, Kruger National Park, South Africa [ Links ]
Kioko, J., Muruthi, P., Omondi, P. & Chiyo, P.I., 2008, 'The performance of electric fences as elephant barriers in Amboseli, Kenya', South African Journal of Wildlife Research 38(1), 52-58. https://doi.org/10.3957/0379-4369-38.1.52 [ Links ]
Knapp, A.K., Burns, C.E., Fynn, R.W., Kirkman, K.P., Morris, C.D. & Smith, M.D., 2006, 'Convergence and contingency in production-precipitation relationships in North American and South African C 4 grasslands', Oecologia 149(3), 456-464. https://doi.org/10.1007/s00442-006-0468-2 [ Links ]
Knapp, A.K., Hoover, D.L., Blair, J.M., Buis, G., Burkepile, D.E., Chamberlain, A. et al., 2012, 'A test of two mechanisms proposed to optimize grassland aboveground primary productivity in response to grazing', Journal of Plant Ecology 5(4), 357-365. https://doi.org/10.1093/jpe/rts020 [ Links ]
Knapp, A.K., Smith, M.D., Collins, S.L., Zambatis, N., Peel, M., Emery, S. et al., 2004, 'Generality in ecology: Testing North American grassland rules in South African savannas', Frontiers in Ecology and the Environment 2(9), 483-491. https://doi.org/10.1890/1540-9295(2004)002[0483:GIETNA]2.0.CO;2 [ Links ]
Koerner, S.E., Burkepile, D.E., Fynn, R.W., Burns, C.E., Eby, S., Govender, N. et al., 2014, 'Plant community response to loss of large herbivores differs between North American and South African savanna grasslands', Ecology 95(4), 808-816. https://doi.org/10.1890/13-1828.1 [ Links ]
Koerner, S.E. & Collins, S.L., 2014, 'Interactive effects of grazing, drought, and fire on grassland plant communities in North America and South Africa', Ecology 95(1), 98-109. https://doi.org/10.1890/13-0526.1 [ Links ]
Kulmatiski, A. & Beard, K.H., 2013a, 'Root niche partitioning among grasses, saplings, and trees measured using a tracer technique', Oecologia 171(1), 25-37. https://doi.org/10.1007/s00442-012-2390-0 [ Links ]
Kulmatiski, A. & Beard, K.H., 2013b, 'Woody plant encroachment facilitated by increased precipitation intensity', Nature Climate Change 3(9), 833. https://doi.org/10.1038/nclimate1904 [ Links ]
Levick, S. & Rogers, K., 2008, 'Patch and species specific responses of savanna woody vegetation to browser exclusion', Biological Conservation 141(2), 489-498. https://doi.org/10.1016/j.biocon.2007.11.001 [ Links ]
Levick, S.R., Asner, G.P., Kennedy-Bowdoin, T. & Knapp, D.E., 2009, 'The relative influence of fire and herbivory on savanna three-dimensional vegetation structure', Biological Conservation 142(8), 1693-1700. https://doi.org/10.1016/j.biocon.2009.03.004 [ Links ]
Levick, S.R., Asner, G.P., Kennedy-Bowdoin, T. & Knapp, D.E., 2009, 'The spatial extent of termite influences on herbivore browsing in an African savanna', Biological Conservation 143(11), 2462-2467. https://doi.org/10.1016/j.biocon.2010.06.012 [ Links ]
MacFadyen, D.N., Avenant, N.L., Van Der Merwe, M. & Bredenkamp, G.J., 2012, 'The influence of fire on rodent abundance at the N'washitshumbe enclosure site, Kruger National Park, South Africa', African Zoology 47(1), 138-146. https://doi.org/10.3377/004.047.0116 [ Links ]
Malhi, Y., Doughty, C.E., Galetti, M., Smith, F.A., Svenning, J.-C. & Terborgh, J.W., 2016, 'Megafauna and ecosystem function from the Pleistocene to the Anthropocene', Proceedings of the National Academy of Sciences 113(4), 838-846. https://doi.org/10.1073/pnas.1502540113 [ Links ]
McCleery, R., Monadjem, A., Baiser, B., Fletcher Jr, R., Vickers, K. & Kruger, L., 2018, 'Animal diversity declines with broad-scale homogenization of canopy cover in African savannas', Biological Conservation 226, 54-62. https://doi.org/10.1016/j.biocon.2018.07.020 [ Links ]
Midgley, J.J., Coetzee, B.W., Tye, D. & Kruger, L.M., 2020, 'Mass sterilization of a common palm species by elephants in Kruger National Park, South Africa', Scientific Reports 10(1), 1-5. https://doi.org/10.1038/s41598-020-68679-8 [ Links ]
Midgley, J.J., Gallaher, K. & Kruger, L.M., 2012, 'The role of the elephant (Loxodonta africana) and the tree squirrel (Paraxerus cepapi) in marula (Sclerocarya birrea) seed predation, dispersal and germination', Journal of Tropical Ecology 28(2), 227-231. https://doi.org/10.1017/S0266467411000654 [ Links ]
Namah, J., Midgley, J.J. & Kruger, L.M., 2019, 'Reproductive biology of the sausage tree (Kigelia africana) in Kruger National Park, South Africa', koedoe 61(1), 1-7. https://doi.org/10.4102/koedoe.v61i1.1512 [ Links ]
O'Keefe, T. & Alard, G.F., 2002, 'Effects of herbivores and fire on riparian and upland savanna ecosystems: Field Operations Manual for Herbivore & Fire Exclosures on the Sabie and Letaba Rivers in the Kruger National Park', SANParks Report, Scientific Services, Skukuza, South African National Parks. [ Links ]
Paterson, D. & Steenekamp, P., 2003, Soil Survey of Nkuhlu exclosure, Agricultural Research Council, Institute for Soil, Climate and Water, Pretoria, South Africa. [ Links ]
Pellegrini, A.F., Refsland, T., Averill, C., Terrer, C., Staver, A.C., Brockway, D.G. et al., 2021, 'Decadal changes in fire frequencies shift tree communities and functional traits', Nature Ecology & Evolution 5(4), 504-512. https://doi.org/10.1038/s41559-021-01401-7 [ Links ]
Rogers, K.H., 2003, 'Adopting a heterogeneity paradigm: Implications for management of protected savannas', in J.T. Du Toit, K.H. Rogers & H.C. Biggs (eds.), The Kruger Experience. Ecology and management of savanna heterogeneity, pp. 41-58, Island Press, Washington, DC. [ Links ]
Rowan, J. & Faith, J.T., 2019, 'The Paleoecological impact of grazing and browsing: Consequences of the Late Quaternary large herbivore extinctions', in I.J Gordon & H.H.T Prins (eds.), The Ecology of Browsing and Grazing II, Ecological Studies (Analysis and Synthesis), p. 451, Springer, Cham [ Links ]
Scogings, P.F., 2011, 'Stem growth of woody species at the Nkuhlu exclosures, Kruger National Park: 2006-2010', koedoe 53(1), 01-09. https://doi.org/10.4102/koedoe.v53i1.1035 [ Links ]
Scogings, P.F., 2014, 'Large herbivores and season independently affect woody stem circumference increment in a semi-arid African savanna', Plant Ecology 215(12), 1433-1443. https://doi.org/10.1007/s11258-014-0400-5 [ Links ]
Scogings, P.F., Hattas, D., Skarpe, C., Hjältén, J., Dziba, L., Zobolo, A. et al., 2015, 'Seasonal variations in nutrients and secondary metabolites in semi-arid savannas depend on year and species', Journal of Arid Environments 114, 54-61. https://doi.org/10.1016/j.jaridenv.2014.11.003 [ Links ]
Scogings, P.F., Hjältén, J. & Skarpe, C., 2011, 'Secondary metabolites and nutrients of woody plants in relation to browsing intensity in African savannas', Oecologia 167(4), 1063-1073. https://doi.org/10.1007/s00442-011-2042-9 [ Links ]
Scogings, P.F., Hjältén, J. & Skarpe, C., 2013, 'Does large herbivore removal affect secondary metabolites, nutrients and shoot length in woody species in semi-arid savannas?', Journal of Arid Environments 88, 4-8. https://doi.org/10.1016/j.jaridenv.2012.08.010 [ Links ]
Scogings, P.F., Hjältén, J., Skarpe, C., Hattas, D., Zobolo, A., Dziba, L. et al., 2014, 'Nutrient and secondary metabolite concentrations in a savanna are independently affected by large herbivores and shoot growth rate', Plant Ecology 215(1), 73-82. https://doi.org/10.1007/s11258-013-0279-6 [ Links ]
Scogings, P.F., Johansson, T., Hjältén, J. & Kruger, J., 2012, 'Responses of woody vegetation to exclusion of large herbivores in semi-arid savannas', Austral Ecology 37(1), 56-66. https://doi.org/10.1111/j.1442-9993.2011.02249.x [ Links ]
Scogings, P.F., Mamashela, T.C. & Zobolo, A.M., 'Deciduous sapling responses to season and large herbivores in a semi-arid African savanna', Austral Ecology 38(5), 548-556. https://doi.org/10.1111/j.1442-9993.2012.02454.x [ Links ]
Siebert, F. & Eckhardt, H.C., 2008, 'The vegetation and floristics of the Nkhuhlu exclosures, Kruger National Park', Koedoe: African Protected Area Conservation and Science 50(1), 126-144. https://doi.org/10.4102/koedoe.v50i1.138 [ Links ]
Siebert, F., Siebert, S.J. & Eckhardt, H.C., 2010, 'The vegetation and floristics of the Letaba exclosures, Kruger National Park, South Africa', Koedoe: African Protected Area Conservation and Science 52(1), 1-12. https://doi.org/10.4102/koedoe.v52i1.777 [ Links ]
Siebert, F. & Scogings, P., 2015, 'Browsing intensity of herbaceous forbs across a semi-arid savanna catenal sequence', South African Journal of Botany 100, 69-74. https://doi.org/10.1016/j.sajb.2015.05.007 [ Links ]
Sisson, D., Hufschmid, J., Jolles, A., Beechler, B. & Jabbar, A., 2017, 'Molecular characterisation of anaplasma species from african buffalo (syncerus caffer) in kruger national park, South Africa', Ticks and Tick-borne Diseases 8(3), 400-406. https://doi.org/10.1016/j.ttbdis.2017.01.003 [ Links ]
Smit, I.P., Peel, M.J., Ferreira, S.M., Greaver, C. & Pienaar, D.J., 2020, 'Megaherbivore response to droughts under different management regimes: Lessons from a large African savanna', African Journal of Range & Forage Science 37(1), 65-80. https://doi.org/10.2989/10220119.2019.1700161 [ Links ]
Smit, I.P. & Prins, H.H., 2015, 'Predicting the effects of woody encroachment on mammal communities, grazing biomass and fire frequency in African savannas', PLoS One 10(9), e0137857. https://doi.org/10.1371/journal.pone.0137857 [ Links ]
Smit, I.P., Roux, D.J., Swemmer, L.K., Boshoff, N. & Novellie, P., 2017, 'Protected areas as outdoor classrooms and global laboratories: Intellectual ecosystem services flowing to-and-from a National Park', Ecosystem Services 28(Part B), 238-250. https://doi.org/10.1016/j.ecoser.2017.05.003 [ Links ]
Smith, F.A., Doughty, C.E., Svenning, J.-C. & Terborgh, J.W., 2016a, 'Megafauna in the earth system', Ecography 39(2), 99-108. https://doi.org/10.1111/ecog.02156 [ Links ]
Smith, M.D., Knapp, A.K., Collins, S.L., Burkepile, D.E., Kirkman, K.P., Koerner, S.E. et al., 2016b, 'Shared drivers but divergent ecological responses: Insights from long-term experiments in Mesic Savanna Grasslands', BioScience 66(8), 666-682. https://doi.org/10.1093/biosci/biw077 [ Links ]
Smith, M.K.S., Smit, I.P., Swemmer, L.K., Mokhatla, M.M., Freitag, S., Roux, D.J. et al., 2021, 'Sustainability of protected areas: Vulnerabilities and opportunities as revealed by COVID-19 in a national park management agency', Biological Conservation 255, 108985. https://doi.org/10.1016/j.biocon.2021.108985 [ Links ]
Steenekamp, P., 2003, Soil Survey of Nkuhlu exclosure, Agricultural Research Council, Institute for Soil, Climate and Water, Pretoria, South Africa. [ Links ]
Stanton, R.A. Jr., Boone IV, W.W., Soto-Shoender, J., Fletcher Jr, R.J., Blaum, N. & McCleery, R.A., 2018, 'Shrub encroachment and vertebrate diversity: A global meta-analysis', Global Ecology and Biogeography 27(3), 368-379. https://doi.org/10.1111/geb.12675 [ Links ]
Staver, A.C., Abraham, J.O., Hempson, G.P., Karp, A.T. & Faith, J.T., 2021, 'The past, present, and future of herbivore impacts on savanna vegetation', Journal of Ecology 109(8), 2804-2822. https://doi.org/10.1111/1365-2745.13685 [ Links ]
Swemmer, A.M., Bond, W.J., Donaldson, J., Hempson, G.P., Malherbe, J. & Smit, I.P., 2018, 'The ecology of drought-a workshop report', South African Journal of Science 114(9-10), 1-3. https://doi.org/10.17159/sajs.2018/5098 [ Links ]
Thoresen, J., Vermeire, M.-L., Venter, Z., Wolfaard, G., Krumins, J., Cramer, M. & Hawkins, H.J., 2021, 'Fire and herbivory shape soil arthropod communities through habitat heterogeneity and nutrient cycling in savannas', Global Ecology and Conservation 25, e01413. https://doi.org/10.1016/j.gecco.2020.e01413 [ Links ]
Treydte, A.C., Baumgartner, S., Heitkönig, I.M., Grant, C.C. & Getz, W.M., 2013, 'Herbaceous forage and selection patterns by ungulates across varying herbivore assemblages in a South African savanna', PLoS One 8(12), e82831. https://doi.org/10.1371/journal.pone.0082831 [ Links ]
Van Coller, H. & Siebert, F., 2015, 'Herbaceous biomass-species diversity relationships in nutrient hotspots of a semi-arid African riparian ecosystem', African Journal of Range & Forage Science 32(3), 213-223. https://doi.org/10.2989/10220119.2014.951394 [ Links ]
Van Coller, H. & Siebert, F., 2020, 'The impact of herbivore exclusion on forb diversity: Comparing species and functional responses during a drought', African Journal of Ecology 58(2), 236-250. https://doi.org/10.1111/aje.12676 [ Links ]
Van Coller, H., Siebert, F., Scogings, P.F. & Ellis, S., 2018, 'Herbaceous responses to herbivory, fire and rainfall variability differ between grasses and forbs', South African Journal of Botany 119, 94-103. https://doi.org/10.1016/j.sajb.2018.08.024 [ Links ]
Van Coller, H., Siebert, F. & Siebert, S.J., 2013, 'Herbaceous species diversity patterns across various treatments of herbivory and fire along the sodic zone of the Nkuhlu exclosures, Kruger National Park', Koedoe 55(1), 01-06. https://doi.org/10.4102/koedoe.v55i1.1112 [ Links ]
Vermeire, M.-L., Thoresen, J., Lennard, K., Vikram, S., Kirkman, K., Swemmer, A.M. et al., 2021, 'Fire and herbivory drive fungal and bacterial communities through distinct above-and belowground mechanisms', Science of the Total Environment 785, 147189. https://doi.org/10.1016/j.scitotenv.2021.147189 [ Links ]
Voysey, M.D., Archibald, S., Bond, W.J., Donaldson, J.E., Carla Staver, A. & Greve, M., 2021, 'The role of browsers in maintaining the openness of savanna grazing lawns', Journal of Ecology 109(2), 913-926. https://doi.org/10.1111/1365-2745.13518 [ Links ]
Wigley, B.J., Fritz, H., Coetsee, C. & Bond, W.J., 2014, 'Herbivores shape woody plant communities in the Kruger National Park: Lessons from three long-term exclosures', Koedoe 56(1), 1-12. https://doi.org/10.4102/koedoe.v56i1.1165 [ Links ]
Wigley-Coetsee, C. & Staver, A.C., 2020, 'Grass community responses to drought in an African savanna', African Journal of Range & Forage Science 37(1), 43-52. https://doi.org/10.2989/10220119.2020.1716072 [ Links ]
Wilcox, K.R., Koerner, S.E., Hoover, D.L., Borkenhagen, A.K., Burkepile, D.E., Collins, S.L. et al., 2020, 'Rapid recovery of ecosystem function following extreme drought in a South African savanna grassland', Ecology 101(4), e02983. https://doi.org/10.1002/ecy.2983 [ Links ]
 Correspondence:
Correspondence:
Corli Wigley-Coetsee
Corli.wigley-coetsee@sanparks.org
Received: 19 Feb. 2021
Accepted: 01 Dec. 2021
Published: 28 Feb. 2022


{kind=link}
{kind=link}