










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.59 n.2 Pretoria 2017
http://dx.doi.org/10.4102/koedoe.v59i2.1434
ORIGINAL RESEARCH
Spatial refuges buffer landscapes against homogenisation and degradation by large herbivore populations and facilitate vegetation heterogeneity
Keoikantse SiangaI; Mario van TelgenII; Jip VroomanII; Richard W.S. FynnI; Frank van LangeveldeII
IOkavango Research Institute, University of Botswana, Botswana
IIResource Ecology Group, Wageningen University, the Netherlands
ABSTRACT
Environmental heterogeneity across savanna landscapes, including different seasonal resources at different distances to water, may play a critical role in maintaining the size and diversity of wildlife populations and the sustainability of their resource base. We investigated whether extensive landscapes with functionally diverse seasonal resources and large waterless regions can mediate the effect of herbivory on plant composition, structure and diversity. Vegetation composition, structure and richness in two different vegetation types (mopane and sandveld woodland) at three distance zones (0 km - 5 km, 10 km - 15 km and > 20 km) from the permanent water of the Okavango Delta and Linyanti Swamps were surveyed. We modelled vegetation response of the most abundant species to herbivory in relation to distance from permanent water, and included fire frequency as a covariate. Trees favoured by elephants during the dry season occurred typically as immature, pollarded populations within 5 km of permanent water sources while mature tall populations of these species were found far from water (> 10 km - 15 km). Similarly, short high-quality grazing grasses were higher in abundance within 5 km of permanent water, whereas taller high-quality perennial grasses peaked in abundance beyond 20 km from permanent water. Trends in herbaceous richness with distance from water were contingent upon vegetation type, while tree richness did not change with distance from water. Spatial refuges in waterless regions of landscapes facilitate the creation of heterogeneity of vegetation structure, composition and richness by large herds of mammalian herbivores. Therefore, the extension of herbivore dry season foraging range, for example, by the creation of artificial water points (AWP) in backcountry woodlands, could seriously undermine the resilience of landscapes to herbivory by reducing the availability of spatial refuges. Consequently, it reduces the resilience of herbivore and predator populations that depend on these spatial refuges. We strongly advise that future scientific work, and management and policy actions should be focused on the identification and sustaining of these spatial refuges in wildlife areas.
CONSERVATION IMPLICATIONS: Management and policy actions should be focused on the identification and sustainability of spatial refuges in wildlife areas. Too many AWP in backcountry woodlands could undermine the resilience of landscapes to herbivory by reducing the proportion of landscapes beyond 15 km from permanent water.
Introduction
Environmental heterogeneity across savanna landscapes (spatial heterogeneity) may play a key role in determining plant (MacFayden et al. 2016), large mammal and bird diversity (Fuhlendorf et al. 2006; Harrington et al. 1999; Krook, Bond & Hockey 2007; Mills & Gorman 1997). This heterogeneity also determines the strength of density-dependent feedbacks on large herbivore population growth (Hobbs et al. 2008; Wang et al. 2006) and the viability of their populations (Hobbs et al. 2008; Hopcraft, Olff & Sinclair 2010; Illius & O'Connor 2000; Owen-Smith 2004). Spatial heterogeneity is determined by spatial variation in abiotic factors such as physical, chemical, topographic, hydrological, climatic and biotic factors (Fynn 2012; Hobbs et al. 2008; Hopcraft et al. 2010; Owen-Smith 2004).
Distance to water during the dry season, which affects herbivore foraging intensity, habitat structure and niche diversity, is another form of spatial heterogeneity. Plant species favoured by large herbivores may be spared from excessive herbivore impact in regions of landscapes far from permanent water (O'Connor, Goodman & Clegg 2007). Sable (Hippotragus niger) and roan (Hippotragus equinus) antelope depend upon waterless, backcountry parts of landscapes to avoid higher concentrations of predators and other herbivore species closer to permanent water (Harrington et al. 1999; Havemann 2014; Hensman et al. 2014). These species have specialised narrow mouths adapted to foraging on taller grasses in these low herbivore density backcountry habitats (Codron et al. 2008; Fynn et al. 2016). Similarly, mesocarnivores, such as African wild dog (Lycaon pictus), may also be forced by competition with larger carnivores to focus their hunting in regions of landscapes with lower abundances of their favoured prey (Mills & Gorman 1997). Thus, large distances from water during the dry season are a key characteristic of landscapes in African savannas, creating spatial refuges for various plant and animal species. For example, in the Kruger National Park, where artificial water provision (AWP) from boreholes in waterless, backcountry parts of landscapes removed these spatial refuges, which may have resulted in a collapse of rare herbivore populations (Harrington et al. 1999).
Fragmentation of ecosystems and landscapes by anthropogenic activities poses a threat to the viability of wildlife populations because of the restricted access to spatially heterogeneous landscapes, and by degrading resources caused by an increase in the strength of coupling of herbivores to their resources (Fynn 2012; Hopcraft et al. 2010). Herbivores become increasingly coupled to their resources as the opportunity for adaptive foraging declines with greater restrictions on movement and with decreasing heterogeneity (Fynn 2012; Hobbs et al. 2008; Hopcraft et al. 2010). Thus, extensive heterogeneous landscapes, where herbivores are able to move seasonally between functionally different habitats and avoid heavily utilised areas and where large distances from permanent water exist, are likely to be more resilient to the impacts of large herbivore populations but this resilience is likely declining with increasing modification of ecosystems by agriculture, fencing and AWP.
The African elephant (Loxodonta africana) is notorious for its effects on woody vegetation (O'Connor et al. 2007) and there is much concern that growing elephant populations may negatively influence ecosystem functioning and diversity in protected areas (Cumming et al. 1997). Elephants have induced dramatic changes in ecosystems by transforming woodlands into open savanna or grasslands (Laws 1970; Spinage 1994), effects that may be compounded by interactions with fire (O'Connor et al. 2007). Elephants can kill large trees and shrubs through pollarding and ring-barking, and young plants through uprooting (O'Connor et al. 2007; Vesey-FitzGerald 1973) reducing the density and structure of woody vegetation (Asner & Levick 2012; Ben-Shahar 1998; Cumming et al. 1997; Glover 1963; Teren & Owen-Smith 2010).
However, the ability of elephants to access food is strongly constrained by water availability, with bulls and breeding herds generally limited to foraging within 15 km and 5 km of water, respectively (O'Connor et al. 2007; Young & Van Aarde 2010). Thus, areas farther from water than the viable foraging range of elephants (> 15 km) are likely to represent a spatial refuge from elephant impact, for vegetation and herbivores that depend on these spatial refuges (O'Connor et al. 2007). The diet of elephants varies seasonally, with green grass and herbs being favoured during the wet season, browse during the early dry season and tree roots and bark by the late dry season (O'Connor et al. 2007). Certain woody species appear to be particularly favoured during the dry season, and it is expected that these species will be heavily impacted in areas within 5 km of water during the dry season (maximum foraging range of breeding herds) with little impact beyond 15 km from permanent water (maximum foraging range of bulls). We refer to this as the distance from water spatial refuge hypothesis (DWSR hypothesis) (O'Connor et al. 2007). The DWSR hypothesis is also applicable to large populations of other grazers, whose year-round grazing effects may eliminate high-quality tufted perennial grasses that require long periods of recovery from grazing (Fynn 2012). Thus, one may expect to find a higher abundance of high-quality, tufted perennial grasses within habitat types beyond the maximum foraging range of herbivores from water during the dry season and a higher abundance of more grazing-tolerant shorter grasses closer to available water. This suggests that herbivory gradients across unmodified landscapes facilitate greater compositional and structural heterogeneity than in landscapes where the distance to dry season water has been greatly modified by AWP.
Against this background, AWP is increasingly used in protected areas in African savannas, with the consequence that landscapes are losing spatial refuges from year-round herbivory for plants and animals, even in very large protected areas (e.g. Redfern et al. 2003). Owing to the potential for AWP to disrupt spatial refuges for plants and animals, it is becoming increasingly urgent to study the effects of herbivory on vegetation and animals in extensive, unfragmented landscapes that still have large areas without AWP and with large distances away from permanent water. The northern conservation area of Botswana is one of the few remaining open wildlife systems in Africa (about 100 000 km2 on the Botswana side and extending into protected areas in Namibia and Zimbabwe) supporting the largest African elephant population globally at around 130 000 (Chase 2011; Chase et al. 2016). This Savuti-Mababe-Linyanti ecosystem (SMLE) is a relatively pristine region of the northern conservation area containing extensive savanna woodlands sandwiched between the permanent water sources of the Okavango Delta and Linyanti Swamps, where large areas of woodland may occur > 20 km from permanent water sources, well beyond the maximum movement distance of the more mobile bull elephants during the dry season. Thus, the extensive unmodified landscapes of the SMLE with their large distances from available water during the dry season (> 20 km), their lack of anthropogenically induced barriers to herbivore movement, and their large populations of elephant, buffalo (Syncerus caffer) and zebra (Equus quagga) provide an ideal study site to gain insights into landscape-mediated herbivore controls of vegetation composition, structure and diversity.
Considering that a large proportion of the landscapes of the SMLE occurs beyond the reach of large herbivores during the dry season (spatial refuge) we hypothesise that greater year-round herbivore density within 5 km of permanent water will negatively affect the woody structure, grass cover and plant richness in this zone but have little effect in zones farther from permanent water. Thus at a landscape scale, herbivores are likely to create spatial heterogeneity in plant composition, diversity and structure, as opposed to homogenisation of plant composition and structure and a decline of species richness in less resilient landscapes.
Methods
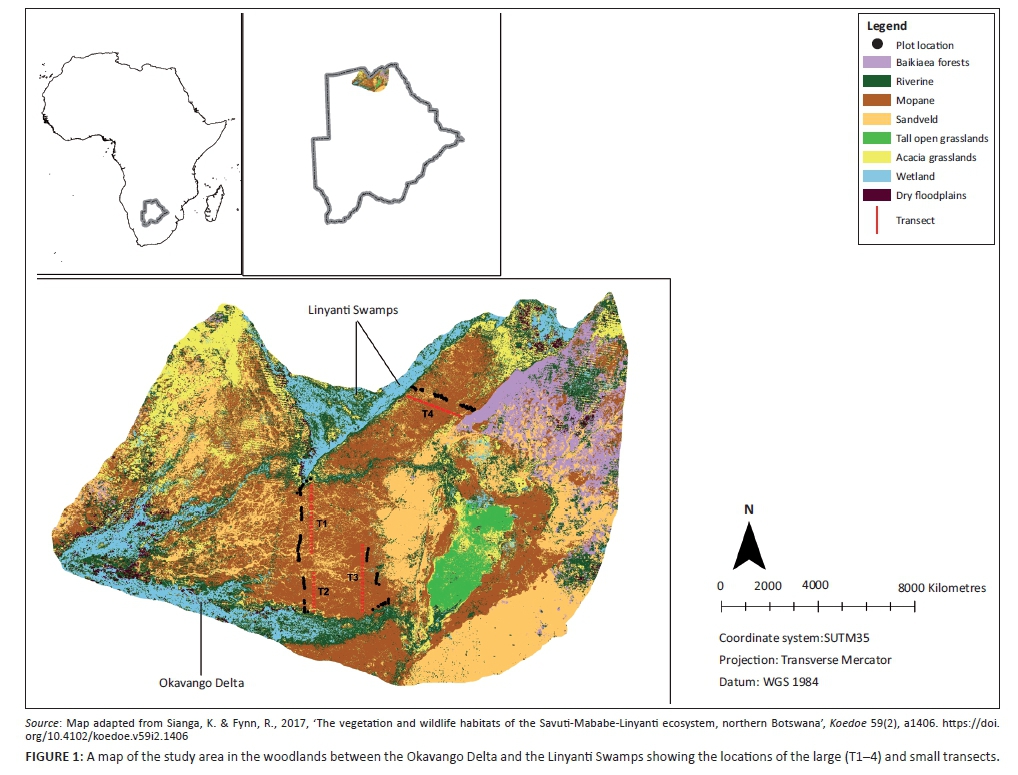
Study area
This study was conducted over the 2014 wet season in the northern conservation area of Botswana, in the extensive open woodland landscapes between the Okavango Delta in the south west and the Linyanti Swamps in the north east (Figure 1). Rainfall in the area averages between 500 mm and 600 mm in the Okavango region (western boundary of the SMLE) and Chobe Enclave region (eastern boundary of the SMLE), respectively (Fynn, Chase & Roder 2014). Rainfall is received between November and April (Botswana Meteorological Services), with maximum daily temperatures between 35 °C and 40 °C (Fynn et al. 2014). The Kwando River that feeds into the Linyanti Swamps and the Okavango River that feeds into the Okavango Delta are the primary sources of permanent water in the ecosystem (Figure 1). One artificial water point at Hyena Pan (± 11 km from water), near one of our large transects (T3), has been pumped consistently since 2013, about a year prior to our sampling, and as such unlikely to have affected the vegetation at that time. The major vegetation of the woodlands between the Okavango Delta and Linyanti Swamps is a mosaic of mopane woodland on alluvial soils and sandveld woodland on deep Kalahari sands, which have filled old river channels (Mendelsohn et al. 2010). Mopane woodland is dominated by Colophospermum mopane in the tree layer and Jasminum stenolobum, Zornia glochdiata, Kyllinga buchananii, Commelina forskaolii, Aristida adscensionis, Aristida scabrivalvis and Urochloa trichopus in the herbaceous layer, while sandveld woodland is dominated by Terminalia sericea and Philenoptera nelsii in the tree layer and Ipomoea chloroneura, Oxygonum alatum, Hibiscus mastersianus, Chamaecrista stricta, Aristida stipitata, Digitaria eriantha and Panicum maximum in the herbaceous layer (Sianga & Fynn 2017).
Vegetation sampling
Four large transects (about 27 km each) perpendicular to permanent water sources of the Linyanti Swamps and Okavango Delta (Figure 1) were sampled during the wet season (from January 2014 to end of March 2014) along easy access routes such as cutlines extending out from the two wetland systems and, therefore, across the major axis of the landscape (Figure 1). Each transect was divided into three distance zones as follows: near zone (0 km - 5 km), intermediate zone (10 km - 15 km) and far zone (> 20 km; up to 27 km) from permanent water. Thus, our four large transects represent landscapes with > 40% of their total area being beyond the maximum movement distance of elephant from water during the dry season. We controlled environmental effects by selecting only mopane and sandveld woodland because these are dryland plant communities that occur in all three distance zones. Thus, we avoided the confounding effects of soil type and soil moisture by keeping vegetation and soil types constant along the large transects (we avoided moist riverine communities directly adjacent to floodplains). The mopane-sandveld mosaic makes up over 90% of the total surface area of the dryland vegetation and so provides the greater bulk of the forage resources outside of the wetlands (Figure 1). This study aimed to assess the integrated effect of herbivory on vegetation in both the wet and dry season, which is manifest in wet season growth. Additionally, dry season sampling is impossible, since all species senesce and most herbaceous species disappear completely, making it impossible (and spurious) to determine composition and richness during this time of year. Therefore, vegetation sampling was done during the wet season, starting in January for easier identification of species (i.e. presence of flowers or inflorescence).
In each distance zone along the large transects, we laid down 6-7 small transects in each of the two vegetation types (mopane and sandveld). The position of small transects was identified by using two random numbers between 20 m and 100 m, with the first random number taken along an access route (cutline) and the second perpendicular from the cutline into the vegetation. A transect was set at a point where the second number fell. Each small transect was 100 m long and orientated perpendicular to the access road. Small transects were at least 500 m apart if in the same patch of woodland but we generally chose a new patch of sandveld or mopane woodland for each transect (the mopane and sandveld communities consists of a mosaic of patches, each generally several hundred metres wide, determined by patches of alluvial soils alternating with sand filled paleo-river channels).
Plant species richness
For plant species richness, all forb, grass and woody species were enumerated in one 40 m × 20 m releve plot per transect laid down over the first 40 m of each small transect. On large transect 3 (T3) in the > 20 km zone in mopane woodland, only three releve plots were used in the analysis instead of seven, owing to an error in plot sizes (50 m × 20 m instead of 40 m × 20 m) for four releve plots, which we discarded.
Woody vegetation
Woody vegetation structure was assessed at three points on the small transects (0 m, 50 m and 100 m) according to an adaptation of the point-centred quarter (PCQ) method (Mitchell 2010), whereby we added height classes of woody vegetation (0 m - 1 m, 1 m - 2 m, 2 m - 4 m and > 4 m). Per centre point, the area to be sampled was divided into quarters by imagining a line perpendicular to the transect line. For every quarter (hereafter sample point), the closest tree was identified in every height class. To avoid overlap between the sample points, a maximum sampling distance of 25 m to the nearest tree of a specific height class was chosen. For the sampled trees, distance from sampling point to the centre of tree trunk (with the use of a rangefinder) and height (with the use of a measuring pole) (Mitchell 2010) was recorded. Although plotless methods such as the point-centre quarter method may not be suitable for estimating the density of rare species, we restricted our analyses to the three most abundant species and our probability estimates are expected to be robust.
Grasses
Grasses were sampled at five points on the small transects (0 m, 25 m, 50 m, 75 m and 100 m) in 1 m2 quadrats (hereafter sample point). All grass species rooted within each quadrat were listed and their cover estimated (to within 5% accuracy if cover > 10% [Koerner et al. 2014], and to within 1% accuracy if cover < 10%, as not to overestimate the abundance of rare species). Grasses that could not be identified to the species level, mainly because of the absence of inflorescence or small growth stage of the plant in young grasses, were if possible identified to the genus level or else marked as 'unidentified'.
Unknown plants were pressed in the field and brought to the Peter Smith Herbarium (PSUB) collection at the Okavango Research Institute (Maun, Botswana) for identification. All vegetation nomenclature followed Germishuizen et al. (2006).
Herbivore density estimates
Herbivore dung was used to estimate herbivore density in this study area. Elephant and other herbivore dung heaps were counted within 2 m of each side of each small transect. Dung counts of elephants are a reliable index of elephant density (Barnes 2001), which is also true for the other mammalian herbivores (Cromsigt et al. 2009). Owing to the long-lasting nature of elephant dung, dung counts represent current wet season deposits as well as carry over from the previous dry season. Identification of herbivore species based on dung found in this study area was based on Walker (1996) and the expertise of various researchers.
Fire
To test the effects of fire, we used moderate resolution imaging spectroradiometer (MODIS) normalised difference vegetation index (NDVI) time-series to map fire events over the last 15 years. These data were used to determine the burn frequency of each small transect, which was used as a covariate in our statistical models.
Statistical analysis
Data from the two vegetation types (mopane and sandveld) were analysed separately. We analysed the relationships between each of the three response variables, (1) species richness of grasses, forbs and trees, (2) woody species structure (height class) and (3) grass cover in mopane and sandveld woodland, and four predictor variables, (1) distance zone from permanent water, (2) elephant dung count, (3) herbivore dung count and (4) fire frequency, and added one or two (see motivation below) random effects: (1) small transect and (2) sample point, using Bayesian generalized linear mixed models (BGLMMs) with Markov chain Monte Carlo (MCMC) estimation using the MCMCglmm package (Hadfield 2010) in R, Version 3.2.3 (R Core-Team 2013). We tested our hypothesis that greater elephant density closer to permanent water would negatively affect the woody structure, grass cover and plant richness using two approaches: (1) a model that considered distance zone from permanent water and fire frequency and their interaction as fixed effects, where the 0 km - 5 km zone was thought to be subject to year-round elephant occupancy in greater numbers, but occupancy and abundance would decline further from permanent water, especially beyond 20 km during the dry season (O'Connor et al. 2007; Young & Van Aarde 2010) and (2) a model that considered elephant dung count (or herbivore dung count) and fire frequency and their interaction as fixed effects (as elephant dung count strongly differed between distance zone and could cause collinearity in the model). We included fire frequency as a covariate because fire is considered to be a critical variable affecting the composition and structure of woody vegetation (Mudongo, Fynn & Bonyongo 2015). Furthermore, previous studies indicate that fire frequency interacts with herbivory in its effect on plant richness (Collins et al. 1998; Koerner et al. 2014).
Random effects were included in the models based on the rules of thumb: > 5-6 levels per random effect and > 10-20 samples per treatment level or experimental unit (Bolker et al. 2008). Thus, none of our models included the large transects (4 levels) in the random effect structure. The models for grass cover and species richness included only small transect (7 levels) in the random effect structure, while for woody species we also included sample point as a random effect (12 levels), nested within small transect.
For species richness, a Poisson distribution with a log-link (family 'poisson') was specified for both models. Initial models started with an inverse Wishart prior for both the residual and random effect (co)variances [R and G-structures: Pr (σ2) ~ IW (V = 1, nu = 0.002)]. For the response variables, woody structure and grass cover, binomial distributions with logit-links (families 'categorical' and 'multinomial2', respectively) were specified. Initial models started with a prior residual variance component fixed to 1 [R-structure: Pr(σ2) ~ IW(V = 1, fix = 1)] and an inverse Wishart prior for the random effect (co)variances [G-structure: Pr(σ2) ~ IW(V = 1, nu = 0.002)].
We ran all initial models with 500 000 iterations, a burn-in period of 100 000, and a thinning interval of 100 iterations to allow us to store 4000 samples. We examined sampling efficiency and convergence of each initial model through their mixing properties, using trace plots and calculated autocorrelation within and between all fixed and random effects. We only accepted models if the autocorrelation was < 0.1 (Hadfield 2015).
If model sampling efficiency and convergence was unsatisfactory, we used a stepwise process to improve the model's performance: first, by adding a redundant non-identified parameter to the G-structure of the initial prior (parameter expanded priors: alpha.mu = 0 and alpha.V = 1000); then, by using the slice sampling method of Damien, Wakefield and Walker (1999) in combination with both the initial and parameter expanded priors; and, when previous adjustments did not improve mixing properties of the models to a satisfying level, we improved mixing by varying the residual variance from 1 to a maximum of 10. These adjustments may improve mixing properties of the chain and lead to quicker model convergence (Hadfield 2015). If this still did not result in an acceptable level of autocorrelation, we increased the total number of iterations with steps of 500 000 iterations until it did, which required also increasing the thinning and burn-in proportionally (thin = 0.0002 and burn-in = 0.2 of total number of iterations) to maintain the total number of stored samples at 4000. Our final models are specified in supplementary material under Table 1-A1. We validated our models by running each final model three times and calculate their average autocorrelation and checked model convergence using the Gelman and Rubin's convergence diagnostic (Gelman & Rubin 1992), which compares within and between chain variance. Models had converged if the potential scale reduction (PSR) factor was less than 1.1. For woody structural and grass cover responses, we here present only species that were abundant enough to be analysed, for example, to give an adequate model convergence. Hence, hereafter we discuss only three tree species (C. mopane, P. nelsii and T. sericea) and 12 grass species (A. adscensionis, A. scrabrivalvis, A. stipitata, Dactyloctenium giganteum, D. eriantha, Digitaria milanjiana, Eragrostis rigidior, Eragrostis trichophora, P. maximum, Pogonarthria fleckii, Schmidtia pappophoroides and U. trichopus).
Results
We observed a total of 38 tree, 191 forb and 66 grass species. Unidentified grass species (including grasses that were identified only to the genus level) accounted for < 1% cover in both mopane and sandveld woodland.
Plant species richness
Grasses
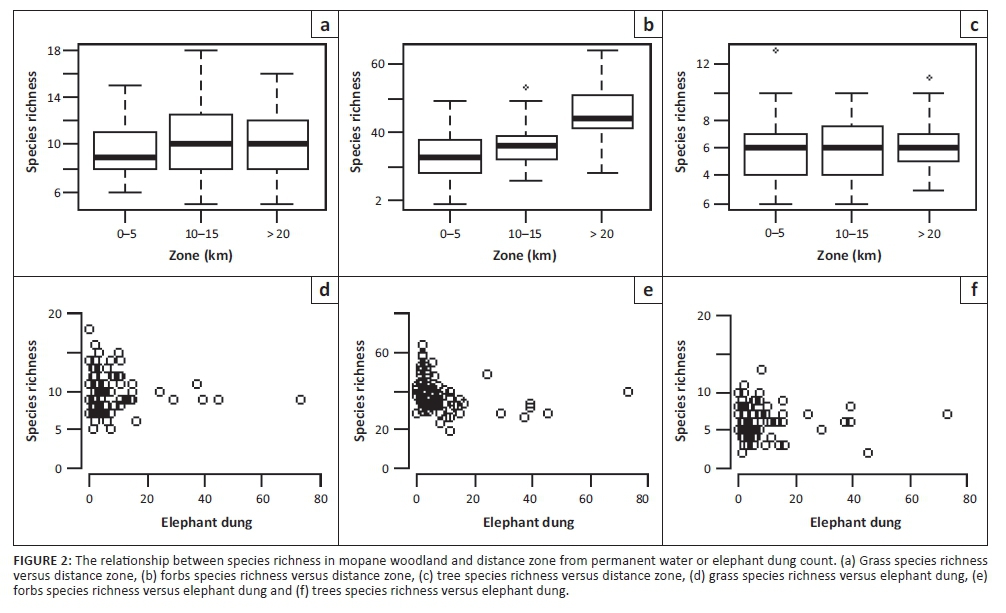
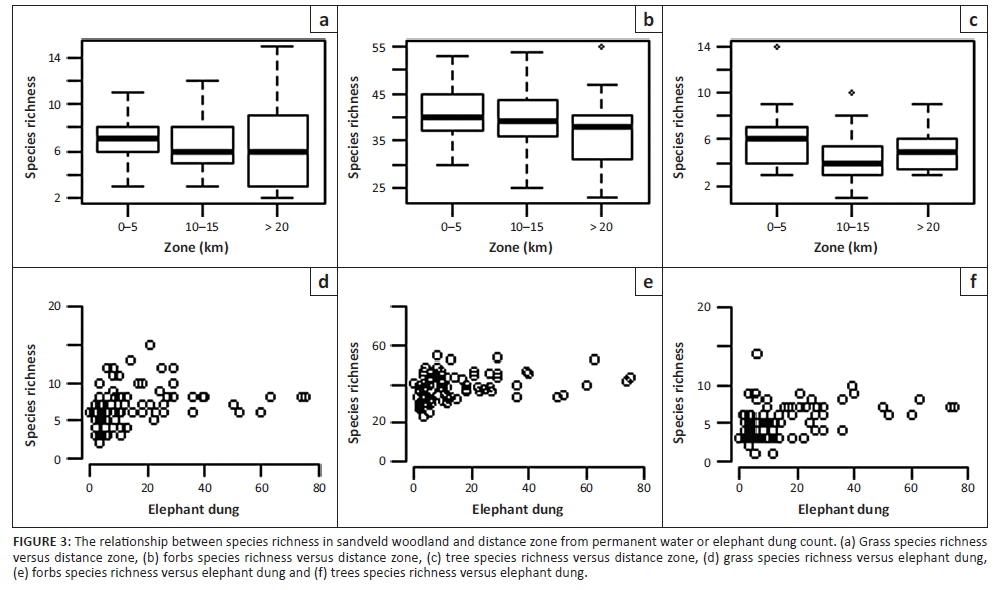
Species richness of grasses, forbs and trees responded differently to distance zone, elephant dung counts and fire frequency and between vegetation types (Table 1; Figures 2 and 3). In mopane woodland, grass richness was not significantly related to distance zone, elephant or herbivore dung count or fire, but in sandveld woodland the far zone (> 20 km) had significantly lower grass richness than the near zone (< 5 km) (pMCMC = 0.012) (Table 1 and Figure 2). Similar to distance zone, grass richness was greater in sites with higher elephant dung counts (but not herbivore dung counts) in sandveld woodland (pMCMC = 0.033), suggesting that higher richness in the near zone in sandveld is related to greater elephant impact in this zone (Table 1 and Figure 3). Similarly, fire frequency was positively related to grass richness in sandveld woodland (pMCMC = 0.0055) (Table 1).
Forbs
Forb richness showed similar responses to grasses being higher in the near zone of sandveld woodland (pMCMC = 0.0055) and increasing with increasing elephant dung count (pMCMC = 0.008) (Table 1 and Figure 3) but showed the opposite response in mopane woodland, being lower in the near than the far zone (pMCMC = 0.004) (Table 1; Figures 2 and 3). While there was no significant relationship between grass richness and herbivore dung count, for forb richness there was a significant positive relationship in sandveld woodland (pMCMC = 0.0095), similar to the relationship with elephant dung count (Table 1 and Figure 3). Thus, there appears to be a general trend of forb and grass richness being affected positively by disturbances from elephants (and to a lesser extent herbivores) in sandveld woodland, but with the opposite trend in mopane woodland.
Trees
In contrast to grasses and forbs, fire, elephant and herbivore dung count, and distance zone were not significantly related to tree richness in sandveld or mopane woodland (Table 1).
Woody species structural responses
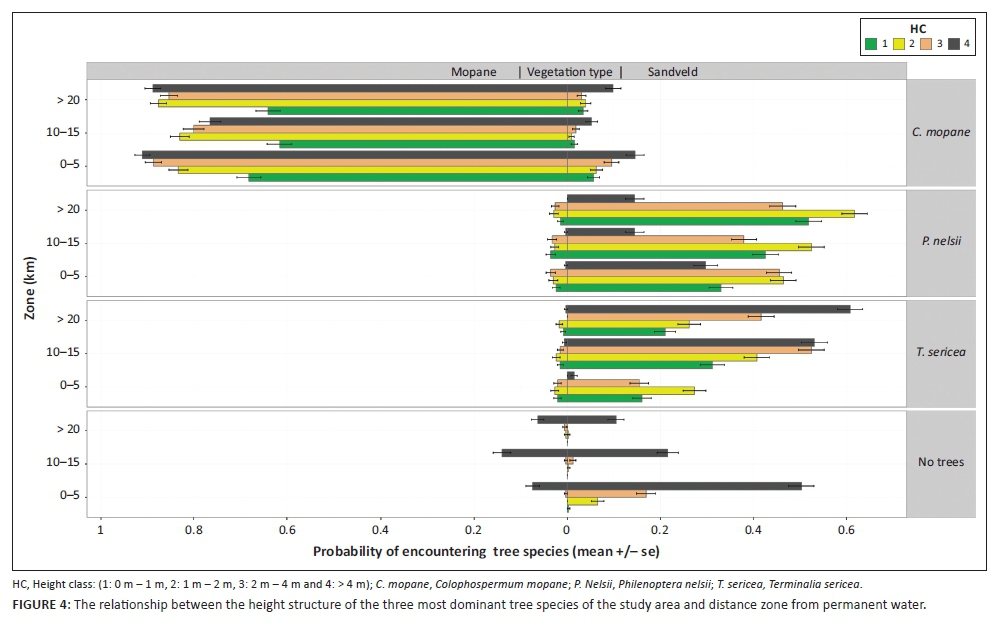
For C. mopane, there was a significant zone × height class 4 interaction where the > 20 km zone (posterior mean = 2.7973, pMCMC = 0.0080) had a significantly higher probability of finding individuals of C. mopane in the > 4 m height class (Table 2-A1; Figure 4). This was also reflected in the elephant dung count × height class interaction where the probability of finding individuals of C. mopane of the > 4 m height class was lower with higher elephant dung count (posterior mean = -0.0570, pMCMC = 0.0295) (Table 2-A1; Figure 4). However, these effects were small relative to those on T. sericea. Fire frequency was negatively related to the probability of observing C. mopane, especially in the taller height classes as shown by the fire frequency × height class interaction (Table 2-A1).
For P. nelsii, there was also a significant zone × height class 4 interaction but in the opposite direction to T. sericea, where the 10 km - 15 km zone (posterior mean = -2.3549, pMCMC = 0.0190) and the > 20 km zone (posterior mean = -4.1166, pMCMC = 0.0003) had significantly lower probability of containing individuals of P. nelsii > 4 m height class (Table 2-A1; Figure 4). In other words, taller trees of P. nelsii were more common in the 0 km - 5 km zone (Figure 4). There was a lower probability of finding individuals of P. nelsii > 4 m tall where fire frequency was greater (posterior mean = -0.9277, pMCMC = 0.0130) (Table 2-A1).
For T. sericea, there was a significant zone × height class 4 interaction, whereby the 10 km - 15 km zone (posterior mean = 6.4833, pMCMC = 0.0020) and the > 20 km zone (posterior mean = 6.3842, pMCMC = 0.0015) had significantly higher probability of containing individuals of T. sericea in the > 4 m height class (Table 2-A1; Figure 4). This was also reflected in the elephant dung count × height class interaction where the probability of finding individuals of T. sericea of the > 4 m height class was lower with higher elephant dung count (posterior mean = -0.1020, pMCMC = 0.0065) (Table 2-A1; Figure 4).
Grass cover responses
Distance from permanent water
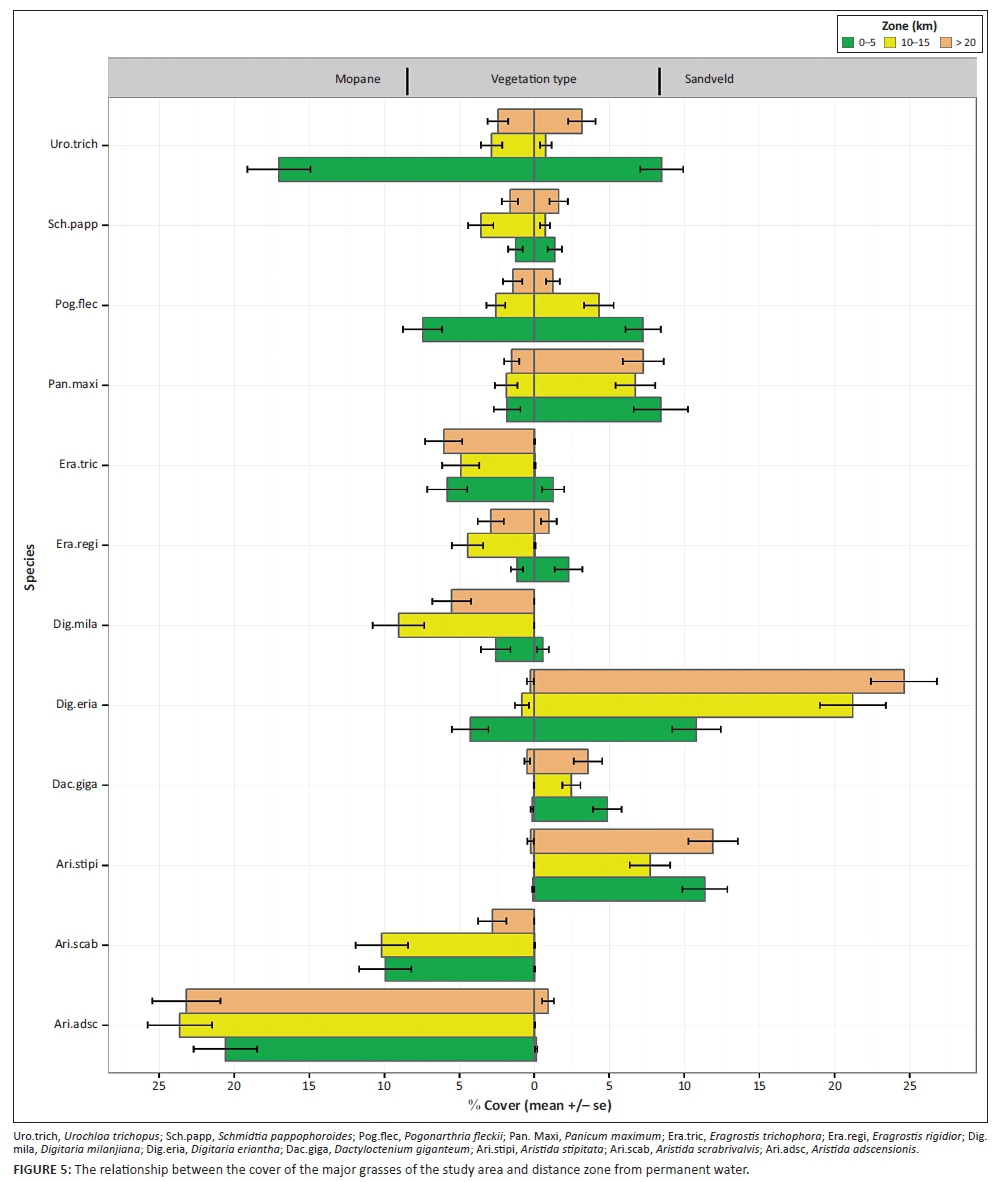
Digitaria eriantha, a high-quality medium or tall perennial grass, was not different across the three distance zones (0 km - 5 km: posterior mean= -14.08, pMCMC = 0.0003; 10 km - 15 km: posterior mean = 2.56, pMCMC = 0.365 and > 20 km: posterior mean = 0.966, pMCMC = 0.759) in mopane woodland (Figure 5; Table 3-A1). However, in its preferred habitat, sandveld, where it is more abundant, D. eriantha had lower cover in the near zone (0 km - 5 km) than the intermediate (10 km - 15 km) (posterior mean = 2.0178, pMCMC = 0.0295) and far zones (> 20 km) (posterior mean = 2.2788, pMCMC = 0.0105) (Table 3-A1; Figure 5) but for P. maximum, another high-quality tall perennial grass, cover in the 10 km - 15 km (posterior mean = 0.5020, pMCMC = 0.6980) and > 20 km (posterior mean = -0.5565, pMCMC = 0.6780) zones did not differ from the 0 km - 5 km zone in sandveld woodland (Figure 5; Table 3-A1). Urochloa trichopus, a high-quality short annual grass, had much greater cover in the 0 km - 5 km zone than the 10 km - 15 km (posterior mean = -6.8563, pMCMC = 0.0005) and > 20 km (posterior mean = -4.4625, pMCMC = 0.0120) zones in sandveld woodland, while in mopane woodland only the 10 km - 15 km (posterior mean = -4.0511, pMCMC = 0.0160) and not the > 20 km (posterior mean = -1.6514, pMCMC = 0.2730) zone was significantly lower than the 0 km - 5 km zone (Figure 5; Table 3-A1). For poor-quality grazing grasses, such as A. adscensionis, the cover of the 10 km - 15 km (posterior mean = 0.7943, pMCMC = 0.2900) and > 20 km (posterior mean = 0.3081, pMCMC = 0.6760) zones in mopane woodland did not differ from the 0 km - 5 km zone (Figure 5; Table 3-A1) as was the case with A. stipitata in sandveld woodland, where its cover in the 10 km - 15 km (posterior mean = -0.6306, pMCMC = 0.5960) and far > 20 km (posterior mean = 0.2058, pMCMC = 0.8750) zones did not differ from the 0 km - 5 km zone (Figure 5; Table 3-A1). However, cover of the poor-quality P. fleckii was greater in the 0 km - 5 km zone in both sandveld (posterior mean = -7.7248, pMCMC = 3e-04) and mopane woodlands (posterior mean = -6.0431, pMCMC = 0.0010) (Figure 5; Table 3-A1).
Elephant dung count
Cover of poor-quality grazing grass such as P. fleckii increased with increasing elephant dung count in mopane (posterior mean = 0.1983, pMCMC = 0.0040) and sandveld woodlands (posterior mean = 0.2602, pMCMC = 0.0003), respectively, but that of A. adscensionis and E. trichophora decreased with increasing elephant dung count in mopane woodland (posterior mean = -0.0513, pMCMC = 0.0100 and posterior mean = -0.5610, pMCMC = 0.0005), respectively (Table 3-A1). The high-quality annual grass D. giganteum also decreased with increasing elephant dung count in sandveld woodland (posterior mean = -0.0881, pMCMC = 0.0235) (Figure 5; Table 3-A1).
Herbivore dung count
Aristida adscensionis, E. rigidior, E. trichophora and S. pappophoroides (all in mopane woodland) decreased with herbivore dung (posterior mean = -0.3058, pMCMC = 0.0340, posterior mean = -1.2515, pMCMC = 0.0320, posterior mean = -1.5979, pMCMC = 0.0145 and posterior mean = -3.0292, pMCMC = 0.0010), respectively (Table 3-A1) while that of U. trichopus in sandveld woodland increased with herbivore dung (posterior mean = 0.5256, pMCMC = 0.0345) (Table 3-A1).
Fire
Fire had no effect on A. adscensionis, U. trichopus, E. rigidior, E. trichophora, P. maximum and P. fleckii in mopane woodland and D. giganteum, D. eriantha, P. maximum, U. trichopus and P. fleckii in sandveld, respectively (Table 3-A1). Grasses such as D. eriantha and S. pappophoroides increased with fire in mopane woodland (Table 3-A1).
Discussion
This study demonstrated that spatial refuges far from permanent water in the relatively unmodified landscapes of this study area provided a buffering effect against homogenisation of vegetation composition, structure and diversity across the landscape by a massive elephant population and large populations of buffalo and other large herbivores. This supported our hypothesis that landscapes that have a large proportion of their area beyond the maximum foraging distance from water during the dry season for most large herbivores (> 15 km) would be resilient to the impacts of large herbivore populations. There is great concern that growing populations of elephant may eventually reach levels that will negatively affect ecosystem function and diversity, and thereby populations of other herbivore species (Cumming et al. 1997). Northern Botswana has by far the largest population of elephants in Africa at approximately 130 000 (Chase 2011; Chase et al. 2016) and there is therefore potential for elephant to homogenise vegetation structure and reduce diversity across landscapes (e.g. Asner & Levick 2012). However, in the landscapes of our study area, which extend to at least 25 km from available water during the dry season, we did not observe homogenisation of vegetation composition, structure or richness but rather heterogeneity in these variables. For example, woody species favoured over the dry season such as T. sericea (Ben-shahar 1993) occurred as immature populations of short, pollarded shrubs within 5 km of water but tall mature populations of these species were typical of the landscape beyond 20 km from permanent water although present within as little as 10 km of water (Figures 4 and 6). By contrast, elephants are impacting favoured woody species right across landscapes in the Kruger National Park (Asner & Levick 2012), where artificial water points have reduced the average distance to available water in the dry season to around 6 km (Redfern et al. 2003), well under our proposed distance of 15 km, thereby eliminating spatial refuges in the landscape for favoured woody species. Similarly, elephants are impacting trees up to 60 km from the Chobe River (Fullman & Child 2013) because of many artificial water points in those distant back country woodlands, enabling elephant to spend all year in the backcountry areas (no spatial refuge available).

In addition, there was no change in woody species richness in sandveld and mopane woodland with distance from water or with elephant dung count (Table 1), suggesting that in these extensive landscapes most woody species populations are not extirpated under high elephant impact but rather survive in a shorter structural state. Thus, in accordance with our hypothesis, it appears that in extensive, heterogeneous landscapes without fences restricting movement, where elephants are able to move seasonally between functionally different habitats and shift their foraging away from over-utilised areas, woody plants are able to persist under a large elephant population (albeit in a shorter state), such that woody diversity is not negatively impacted (sensu Fynn 2012). By contrast, richness of woody species was found to be significantly lower in the elephant-impacted zone (< 10 km) of the Chobe River Front (Fullman & Child 2013) and inside the southern buffalo fence of the Okavango Delta (Cassidy, Fynn & Sethebe 2013). The much greater effect of elephants on tree richness in sandveld and mopane woodlands (the same communities examined in this study) along the southern buffalo fence than in this study could possibly have been caused by elephant movement ranges in these woodlands being constrained to a relatively narrow band of woodland between the floodplains of the Okavango Delta and the southern buffalo fence; hence, no spatial refuges are possible and the southern buffalo fence is likely to further concentrate elephant foraging. Thus in these extensive, relatively unfragmented woodland landscapes, where large parts of the landscape occur greater than 15 km from permanent water (spatial refuges), elephants have not homogenised woody vegetation structure or reduced diversity but rather have created structural heterogeneity across the landscape, as predicted for large unmodified landscapes (Du Toit, Skarpe & Moe 2014).
The contrasting responses of forb and grass richness to herbivory and distance from permanent water in mopane and sandveld woodlands appear to be related to well-known interactions of herbivory and habitat productivity on grass and forb richness, with richness generally being decreased by herbivory in less productive habitats and increased by herbivory in more productive habitats, observed globally (Bakker et al. 2006; Osem, Perevolotsky & Kigel 2002; Proulx & Mazumder 1998) and in African savannas (Burkepile et al. 2017). This is because in more productive habitats herbivory reduces competition from dominant herbaceous species while in unproductive habitats dominants are not productive enough to exclude other species but instead herbivory increases stress on many species (Burkepile et al. 2017). Sandveld woodland generally has a taller and denser grass layer than mopane woodland, which has a lot of bare ground. For example, during the wet season, the total cover of grasses in the > 20 km zone of sandveld woodland was 83.4% ± 29.3%, whereas in mopane woodland total grass cover was 56.9% ± 12.9%. Thus it appears that insufficient fire or herbivory in sandveld woodland results in large dominant herbaceous species excluding small grasses and forbs (e.g. Koerner et al. 2014). Thus overall, richness does not appear to have undergone catastrophic declines under foraging and trampling impacts of a large herbivore biomass, but rather responds to herbivory positively or negatively depending upon habitat productivity and the potential for competitive exclusion (Bakker et al. 2006; Burkepile et al. 2017; Osem et al. 2002; Proulx & Mazumder 1998).
Similarly to patterns of woody vegetation structure with distance from water, we found that taller tufted high-quality perennial grasses such as D. eriantha were more abundant far from water (> 20 km) but that shorter, high-quality grasses such as U. trichopus were most abundant within 5 km of water (Figure 5) demonstrating that gradients of herbivory and trampling by elephant and other large herbivores with increasing distance from permanent water created both compositional and structural heterogeneity in the grass layer. Had there been significant numbers of artificial water points in the backcountry parts of these landscapes, however, we would predict that elephant and other large herbivores would have homogenised the structure of favoured woody species as well as composition and structure of grasses right across the landscape. A decline in the abundance of high-quality tall grasses with year-round grazing in the backcountry woodlands would be expected to negatively impact tall-grass grazers such as buffalo, sable and roan antelope (Fynn et al. 2016). This might have had negative knock-on effects on other biota such as various birds, bats, insects, et cetera. that rely on mature woodland or taller grasses as optimal habitat (e.g. Cumming et al. 1997). Creation of heterogeneity in woodland structure (zones of short, immature and tall, mature woodland) and in grass composition and structure may be expected to maximise niches for various biota, some of which may favour different structural and compositional states of woodland and grassland, as observed for birds with woodland structural heterogeneity (Bradbury et al. 2005) and for birds and insects with grassland structural heterogeneity (Chambers & Samways 1998; Fuhlendorf et al. 2006; Krook et al. 2007).
In addition, creation of heterogeneity in grassland structure facilitates adaptive foraging options and more stable and productive herbivore populations (Hopcraft et al. 2010; Owen-Smith 2002, 2004). Grassland structural heterogeneity also facilitates niche diversity for medium and tall-grass grazers such as roan and sable antelope and buffalo, having a spatial refuge far from water and short-grass grazers such as wildebeest and impala having a special niche closer to water (Fynn et al. 2016). Several studies in this landscape have shown that roan and sable antelope prefer these backcountry areas far from water in both wet and dry seasons, though they will walk to water every 3 or 4 days to drink during the dry season (Havemann 2014; Hensman et al. 2014). In addition, it has been observed that immediately once the rains arrive African buffalo leave the floodplains and move far out from water, presumably to benefit from the higher abundance of D. eriantha far from water (Sianga, Fynn & Bonyongo 2017). These effects of herbivores on composition and structural heterogeneity across these landscapes are likely to be further enhanced by the observation that herbivory also increased heterogeneity in grass and forb richness across the landscape (Table 1). Richness may be important for adaptive foraging options for herbivores in the form of diet breadth expansion (Owen-Smith 2002). Owing to the spatial refuges for tall grasses in this study area, roan and sable populations are indeed maintained here at healthy numbers (Chase 2011; Havemann 2014; Hensman et al. 2014) but have collapsed in Kruger National Park where spatial refuges in landscapes have been eliminated by AWP (Harrington et al. 1999).
Conclusion
It is clear that herbivory on the large distance gradients away from permanent water (> 20 km) in the SMLE has created key diversity, compositional and structural heterogeneity in grass, forb and woody species that is likely to result in greater niche diversity and adaptive foraging options that will enhance biodiversity and herbivore population stability and productivity. In this regard, our findings show that spatial refuges for both grass and woody species operated beyond 15 km from permanent water, with the implication that managers should avoid AWP in backcountry woodlands, because water points will reduce the distance to available water during the dry season and consequently eliminate spatial refuges for plants and animals. If AWP cannot be avoided, for whatever reason, then artificial water points should be spaced at least 50 km apart to ensure a 25 km distance away from dry season water, providing for a large proportion (around 40%) of the landscape area as a spatial refuge from herbivory during the dry season (15 km - 25 km = 40% of the landscape). Consequently, too many artificial water points in backcountry woodlands could seriously undermine the resilience of landscapes to herbivory by reducing the proportion of landscapes beyond 15 km from permanent water (reduced area of spatial refuges), which will reduce the resilience of plant, herbivore and predator populations that depend on these spatial refuges. We strongly advise that future scientific work, and management and policy actions should be focused on the identification and sustaining of these spatial refuges in wildlife areas.
Acknowledgements
The authors are grateful to Great Plains Safaris (Selinda Reserve) for their logistical support during field work. They also thank the Ministry of Environment, Wildlife and Tourism and the Department of Wildlife and National Parks (Botswana) for permitting us to conduct this research in northern Botswana (EWT 8/36/4 XXV [16]). The authors acknowledge the Southern African Science Service Centre for Climate Change and Adaptive Land Management (SASSCAL) for funding this research.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
Authors contributed equally to this article.
References
Asner, G.P. & Levick, S.R., 2012, 'Landscape-scale effects of herbivores on treefall in African savannas', Ecology Letters 15, 1211-1217. https://doi.org/10.1111/j.1461-0248.2012.01842.x [ Links ]
Bakker, E.S., Ritchie, M.E., Olff, H., Milchunas, D.G. & Knops, J.M.H., 2006, 'Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size', Ecology Letters 9, 780-788. https://doi.org/10.1111/j.1461-0248.2006.00925.x [ Links ]
Barnes, R.F.W., 2001, 'How reliable are dung counts for estimating elephant numbers?', African Journal of Ecology 39, 1-9. https://doi.org/10.1111/j.1365-2028.2001.00266.x [ Links ]
Ben-Shahar, R., 1993, 'Patterns of elephant damage to vegetation in northern Botswana', Biological Conservation 65, 249-256. https://doi.org/10.1016/0006-3207(93)90057-8 [ Links ]
Ben-Shahar, R., 1998, 'Changes in structure of Savanna Woodlands in Northern Botswana following the impacts of elephants and fire', Plant Ecology 136, 189-194. https://doi.org/10.1023/A:1009708021735 [ Links ]
Bolker, B.M., Brooks, M.E., Clark, C.J., Geange, S.W., Poulson, J.R., Stevens, M.H.M. et al., 2008, 'Generalized linear mixed models: A practical guide for ecology and evolution', Ecology Letters 24, 127-135. [ Links ]
Bradbury, R.B., Hill, R.A., Mason, D.C., Hinsley, S.A., Wilson, J.D., Balzter, H.A. et al., 2005, 'Modelling relationships between birds and vegetation structure using airborne LiDAR data: A review with case studies from agricultural and woodland environments', Ibis 147, 443-452. https://doi.org/10.1111/j.1474-919x.2005.00438.x [ Links ]
Burkepile, D.E., Fynn, R.W., Thompson, D.I., Lemoine, N.P., Koerner, S.E., Eby, S. et al., 2017, 'Herbivore size matters for productivity-richness relationships in African savannas', Journal of Ecology. https://doi.org//10.1111/1365-2745.12714 [ Links ]
Cassidy, L., Fynn, R. & Sethebe, B., 2013, 'Effects of restriction of wild herbivore movement on woody and herbaceous vegetation in the Okavango Delta Botswana', African Journal of Ecology 51, 513-527. https://doi.org/10.1111/aje.12061 [ Links ]
Chambers, B.Q. & Samways, M.J., 1998, 'Grasshopper response to a 40-year experimental burning and mowing regime, with recommendations for invertebrate conservation management', Biodiversity & Conservation 7, 985-1012. https://doi.org/10.1023/A:1008803513740 [ Links ]
Chase, M. 2011. 'Dry season fixed-wing aerial survey of elephants and wildlife in northern Botswana', Elephants without Borders, San Diego Zoological Society, Department of Wildlife and National Parks. Report, 138. [ Links ]
Chase, M., Schlossberg, S., Griffin, C.R., Philippe J.C., Bouché, P.J.C., Djene, S.W. et al., 2016, 'Continent-wide survey reveals massive decline in African savannah elephants', Peer J 4, e2354. https://doi.org/10.7717/peerj.2354 [ Links ]
Codron, D., Brink, J.S., Rossouw, L., Clauss, M., Codron, J., Lee-thorp, J. et al., 2008, 'Functional differentiation of African grazing ruminants: An example of specialized adaptations to very small changes in diet', Journal of Biology 94, 755-764. https://doi.org/10.1111/j.1095-8312.2008.01028.x [ Links ]
Collins, S.L., Knapp, A.K., Briggs, J.M., Blair, J.M. & Steinauer, E.M., 1998, 'Modulation of diversity by grazing and mowing in native tallgrass prairie', Science 280, 745-747. https://doi.org/10.1126/science.280.5364.745 [ Links ]
Cromsigt, J.P., Van Rensburg, S.J., Etienne, R.S. & Olff, H., 2009, 'Monitoring large herbivore diversity at different scales: comparing direct and indirect methods', Biodiversity and Conservation 18.5, 1219-1231. [ Links ]
Cumming, D.H.M., Fenton, M.B., Rautenbach, I.L., Taylor, R.D., Cumming, G.S., Cumming, M.S. et al., 1997, 'Elephants, woodlands and biodiversity in southern Africa', South African Journal of Science 93, 231-236. [ Links ]
Damien, P., Wakefield, J. & Walker, S., 1999, 'Gibbs sampling for Bayesian non-conjugate and hierarchical models by using auxiliary variables', Journal of the Royal Statistical Society Series B-Statistical Methodology 61, 331-344. https://doi.org/10.1111/1467-9868.00179 [ Links ]
Du Toit, J.T., Skarpe, C. & Moe, S.R., 2014, 'Elephants and heterogeneity in savanna landscapes', in C. Skarpe, J.T. Du Toit & S.R. Moe (eds.), Elephants and savanna woodland ecosystems, pp. 289-298, Wiley Blackwell, Oxford. [ Links ]
Fuhlendorf, S.D., Harrell, W.C., Engle, D.M., Hamilton, R.G., Davis, C.A. & Leslie, D.M., 2006, 'Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing', Ecological Applications 16, 1706-1716. https://doi.org/10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2 [ Links ]
Fullman, T.J. & Child, B., 2013, 'Water distribution at local and landscape scales affects tree utilization by elephants in Chobe National Park, Botswana', African Journal of Ecology 51, 235-243. https://doi.org/10.1111/aje.12026 [ Links ]
Fynn, R.W.S., 2012, 'Functional resource heterogeneity increases livestock and rangeland productivity', Rangeland Ecology & Management 65, 319-329. https://doi.org/10.2111/REM-D-11-00141.1 [ Links ]
Fynn, R.W.S., Augustine, D.J., Peel, M.J.S. & De Garine-Wichatitsky, M., 2016, 'Strategic management of livestock to improve biodiversity conservation in African savannahs: A conceptual basis for wildlife-livestock coexistence', Journal of Applied Ecology 53, 388-397. https://doi.org/10.1111/1365-2664.12591 [ Links ]
Fynn, R.W.S., Chase, M. & Roder, A., 2014, 'Functional habitat heterogeneity and large herbivore seasonal habitat selection in northern Botswana', South African Journal of Wildlife Research 44, 1-15. https://doi.org/10.3957/056.044.0103 [ Links ]
Gelman, A. & Rubin, D.B., 1992, 'Inference from iterative simulation using multiple sequences', Statistical Science 7, 457-472. https://doi.org/10.1214/ss/1177011136 [ Links ]
Germishuizen, G., Meyer, N.L., Steenkamp, Y. & Keith, M., 2006, A checklist of South African plants, SABONET/SANBI, SABONET, Pretoria. [ Links ]
Glover, J., 1963, 'The elephant problem at Tsavo', African Journal of Ecology 1, 30-39. https://doi.org/10.1111/j.1365-2028.1963.tb00176.x [ Links ]
Hadfield, J.D., 2010, 'MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R package', Journal of Statistical Software 33, 1-22. https://doi.org/10.18637/jss.v033.i02 [ Links ]
Hadfield, J.D., 2015, MCMCglmm course notes. [ Links ]
Harrington, R., Owen-Smith, N., Viljoen, P.C., Biggs, H.C, Mason, D.R. & Funston, P., 1999, 'Establishing the causes of the roan antelope decline in the Kruger National Park, South Africa', Biological Conservation 90, 69-78. https://doi.org/10.1016/S0006-3207(98)00120-7 [ Links ]
Havemann, C.P., 2014, 'Population dynamics and foraging ecology of Roan antelope in Northern Botswana', MSc thesis, University of Pretoria, Pretoria, South Africa. [ Links ]
Hensman, M.C., Owen-Smith, N., Parrini, F. & Bonyongo, C.M., 2014, 'Home range occupation and habitat use of sable antelope in the Okavango Delta region of northern Botswana', African Journal of Ecology 52, 237-245. https://doi.org/10.1111/aje.12111 [ Links ]
Hobbs, N.T., Galvin, K.A., Stokes, C.J., Lackett, J.M., Ash, A.J., Boone, R.B. et al., 2008, 'Fragmentation of rangelands: Implications for humans, animals, and landscapes', Global Environmental Change-Human & Policy Dimensions 18, 776-785. https://doi.org/10.1016/j.gloenvcha.2008.07.011 [ Links ]
Hopcraft, J.G.C., Olff, H. & Sinclair, A., 2010, 'Herbivores, resources and risks: Alternating regulation along primary environmental gradients in savannas', Trends in Ecology & Evolution 25, 119-128. https://doi.org/10.1016/j.tree.2009.08.001 [ Links ]
Illius, A.W. & O'Connor, T.G., 2000, 'Resource heterogeneity and ungulate population dynamics', Oikos 89, 283-294. https://doi.org/10.1034/j.1600-0706.2000.890209.x [ Links ]
Koerner, S.E., Burkepile, D.E., Fynn, R.W.S., Burns, C.E., Eby, S., Govender, N. et al., 2014, 'Plant community response to loss of large herbivores differs between North American and South African savanna grasslands', Ecology 95, 808-816. https://doi.org/10.1890/13-1828.1 [ Links ]
Krook, K., Bond, W.J. & Hockey, P.A., 2007, 'The effect of grassland shifts on the avifauna of a South African savanna', Ostrich-Journal of African Ornithology 78, 271-279. https://doi.org/10.2989/OSTRICH.2007.78.2.24.104 [ Links ]
Laws, R.M., 1970, 'Elephants as agents of habitat and landscape change in East Africa', Oikos 21, 1-15. https://doi.org/10.2307/3543832 [ Links ]
MacFayden, S., Hui, C., Verberg, P.H. & Van Teeffelen, A.J.A., 2016, 'Quantifying spatiotemporal drivers of environmental heterogeneity in Kruger National Park, South Africa', Landscape Ecology 31, 2013-2029. https://doi.org/10.1007/s10980-016-0378-6 [ Links ]
Mendelsohn, J., Vanderpost, C., Ramberg, L., Murray-Hudson, M., Wolski, P. & Mosepele, K., 2010, Okavango Delta: Floods of life, Raison, Windhoek. [ Links ]
Mills, M.G.L. & Gorman, M., 1997, 'Factors affecting the density and distribution of wild dogs in the Kruger National Park', Conservation Biology 11, 1397-1406. https://doi.org/10.1046/j.1523-1739.1997.96252.x [ Links ]
Mitchell, K., 2010, 'Quantitative analysis by the point-centered quarter method', arXiv preprint arXiv, 1010.3303. [ Links ]
Mudongo, E., Fynn, R. & Bonyongo, M.C., 2015, 'Influence of fire on woody vegetation density, cover and structure at Tiisa Kalahari Ranch in western Botswana', Grassland Science 62, 3-11. https://doi.org/10.1111/grs.12110 [ Links ]
O'Connor, T.G., Goodman, P.S. & Clegg, B., 2007, 'A functional hypothesis of the threat of local extirpation of woody plant species by elephant in Africa', Biological Conservation 136, 329-345. https://doi.org/10.1016/j.biocon.2006.12.014 [ Links ]
Osem, Y., Perevolotsky, A. & Kigel, J., 2002, 'Grazing effect on diversity of annual plant communities in a semi-arid rangeland: Interactions with small-scale spatial and temporal variation in primary productivity', Journal of Ecology 90, 936-946. https://doi.org/10.1046/j.1365-2745.2002.00730.x [ Links ]
Owen-Smith, N., 2002, Adaptive herbivore ecology: From resources to populations in variable environments, Cambridge University Press, United Kingdom. [ Links ]
Owen-Smith, N., 2004, 'Functional heterogeneity in resources within landscapes and herbivore population dynamics', Landscape Ecology 19, 761-771. https://doi.org/10.1007/s10980-005-0247-2 [ Links ]
Proulx, M. & Mazumder, A., 1998, 'Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems', Ecology 79, 2581-2592. https://doi.org/10.1890/0012-9658(1998)079[2581:ROGIOP]2.0.CO;2 [ Links ]
R Core-Team, 2013, R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, viewed 11 April 2015, from http://www.R-project.org [ Links ]
Redfern, J.V., Grant, R., Biggs, H. & Getz, W.M., 2003, 'Surface-water constraints on herbivore foraging in the Kruger National Park, South Africa', Ecology 84, 2092-2107. https://doi.org/10.1890/01-0625 [ Links ]
Sianga, K. & Fynn, R., 2017, 'The vegetation and wildlife habitats of the Savuti-Mababe-Linyanti ecosystem, northern Botswana', Koedoe 59(2), a1406. https://doi.org/10.4102/koedoe.v59i2.1406 [ Links ]
Sianga, K., Fynn, R.W.S. & Bonyongo, M.C., 2017, 'Seasonal habitat selection by African buffalo Syncerus caffer in the Savuti-Mababe-Linyanti ecosystem of northern Botswana', Koedoe 59(2), a1382. https://doi.org/10.4102/koedoe.v59i2.1382 [ Links ]
Spinage, C.A., 1994, Elephants, Poyser, T. and Poyser, AD Ltd., London. [ Links ]
Teren, G. & Owen-Smith, N., 2010, 'Elephants and riparian woodland changes in the Linyanti region, northern Botswana', Pachyderm 47, 18-25. [ Links ]
Vesey-Fitzgerald, D.F., 1973, 'Animal impact on vegetation and plant succession in lake Manyara National Park, Tanzania', Oikos 24, 314-324. https://doi.org/10.2307/3543890 [ Links ]
Walker, C., 1996, 'Signs of the wild', Struik Publishers, Cape Town, South Africa. [ Links ]
Wang, G., Hobbs, N.T., Boone, R.B, Illius, A.W., Gordon, I.J., Gross, J.E. et al., 2006, 'Spatial and temporal variability modify density dependence in populations of large herbivores', Ecology 87, 95-102. https://doi.org/10.1890/05-0355 [ Links ]
Young, K.D. & Van Aarde, R.J., 2010, 'Density as an explanatory variable of movements and calf survival in savanna elephants across Southern Africa', Journal of Animal Ecology 79, 662-673. https://doi.org/10.1111/j.1365-2656.2010.01667.x [ Links ]
 Correspondence:
Correspondence:
Keoikantse Sianga
keosianga@gmail.com
Received: 08 Aug. 2016
Accepted: 01 Feb. 2017
Published: 23 May 2017
Note: Additional supporting information may be found in the online version of this article as Online Appendix 1: https://doi.org/10.4102/koedoe.v59i2.1434-1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}