










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.59 n.1 Pretoria 2017
http://dx.doi.org/10.4102/koedoe.v59i1.1432
REVIEW ARTICLE
Tsetse flies should remain in protected areas in KwaZulu-Natal
Adrian J. Armstrong; Andy Blackmore
Scientific Services, Ezemvelo KZN Wildlife, South Africa
ABSTRACT
The proposal to eradicate tsetse flies from South Africa, including its protected areas, via the sequential aerosol technique combined with the sterile insect technique to reduce trypanosomiasis in cattle did not present an appropriate analysis of the impacts that implementation of the proposal would have on biodiversity. Not only would the implementation of the proposal be contrary to South African laws protecting and conserving biodiversity, but it would also have negative consequences for the conservation of biodiversity. Some of the negative consequences are reviewed, including extirpations and negative impacts on ecological and ecosystem processes and services. Alternative strategies to control trypanosomiasis in cattle effectively in a more environment-friendly manner are presently available and others will almost certainly become available in the not-too-distant future.
CONSERVATION IMPLICATIONS: Environmental protection, promotion of conservation and sustainable use of the environment are all deeply seated in South Africa's law. Rural livestock husbandry considerations and biodiversity conservation are not mutually exclusive and the importance of one cannot supersede the other. The eradication proposal is seen to be environmentally damaging and therefore it is concluded that the purpose of this proposed eradication exercise is unconstitutional, contrary to various multilateral agreements South Africa has entered into and contrary to good environmental governance.
Introduction
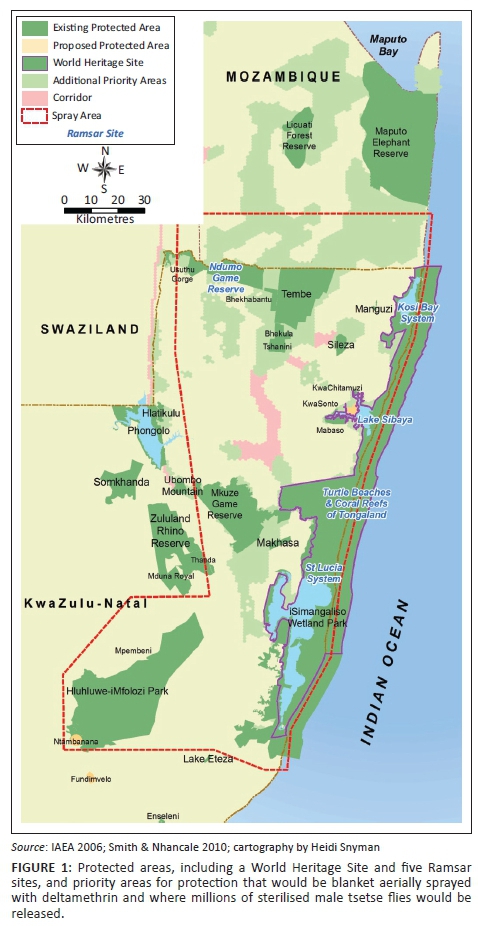
Tsetse flies are the most important vectors for African animal trypanosomiasis (AAT) in cattle. Although indigenous wild mammals have resistance to trypanosomes, susceptible cattle develop clinical trypanosomiasis. The eradication of tsetse flies from KwaZulu-Natal (and therefore South Africa) has been proposed by several authors (International Atomic Energy Agency [IAEA] 2006; Kappmeier Green, Potgieter & Vreysen 2007). The IAEA has a Technical Cooperation Project, 'Supporting a Feasibility Study to Eradicate Tsetse from Southern Mozambique, South Africa and Swaziland', under the Joint Food and Agriculture Organization (FAO)/IAEA Programme, 'Nuclear Techniques in Food and Agriculture', as part of its peaceful use of nuclear technology theme (IAEA n.d.). This eradication proposal (EP) recommends using the sequential aerosol technique (SAT) over north-eastern KwaZulu-Natal (neKZN; Figure 1), a region with a number of very important protected areas, including a World Heritage Site (WHS) and five Ramsar sites. SAT involves blanket spraying an insecticide from the air several times to suppress tsetse fly populations. Deltamethrin, a synthetic pyrethroid, at a low concentration of active ingredient, has been proposed as the insecticide of choice (IAEA 2006). The SAT would be followed by the sterile insect technique (SIT) in which millions of sterilised male tsetse flies are released to eradicate the flies through suppression of reproduction.
Although generally believed to have low environmental build-up and greater specificity to invertebrates, deltamethrin is pernicious in aquatic systems. It has been recorded in fish liver, water, and sediment samples in South Africa, and it has been found in trace amounts in the Ndumo Game Reserve and Tembe Elephant Park (Ansara-Ross et al. 2012), both of which are in the proposed spray region. Further contamination by deltamethrin of protected areas that are held in public trust for the conservation of biodiversity should not be permitted. Deltamethrin poses a significant risk to non-target species, including aquatic insects and macrocrustaceans (Ansara-Ross et al. 2012). Implementation of the EP would have detrimental consequences that are not restricted to tsetse flies and trypanosomes, but will include several non-target species of conservation concern and others of great biodiversity and ecosystem service value. Area-wide eradication of tsetse flies in KwaZulu-Natal and the adjacent part of Mozambique and Swaziland is still promoted in the scientific literature (e.g. De Beer, Venter & Vreysen 2015). The study by Perkins and Ramberg (2004) on the impacts of deltamethrin applied via the SAT in 2001 and 2002 to eradicate tsetse flies from the Okavango Delta (OD) and the recovery study in 2003 have been used to justify why the EP would be acceptable from an environmental viewpoint. Little critique of, or research on, the negative impacts that the eradication course of action would have on biodiversity has been published. Hence, this review gives reasons why the EP should not be carried out from a biodiversity conservation viewpoint, and indeed may not be carried out in protected areas.
The biodiversity case against implementation of the eradication proposal
Various South African legislations promulgated to conserve biodiversity would be contravened
The consideration and implementation of a strategy to eradicate tsetse flies within a region of South Africa, and importantly in existing protected areas, is seen to contravene South Africa's constitutional and environmental laws and some African and global multilateral agreements this country has entered into.
The Environmental Right in the Bill of Rights in the Constitution of South Africa affords a right to present and future generations to have, inter alia, protection and conservation of the environment which is devoid of ecological degradation, and that all use of the environment must be ecologically sustainable within a context of justifiable economic and social development. Therefore, the EP presents a significant risk of contravening the supreme law of the country. Derived from the Environmental Right, the National Environmental Management Act (No. 107 of 1998) (NEMA) provides for a set of environmental principles that:
serve as guidelines by reference to which any organ of state must exercise any function when taking any decision in terms of this Act or any statutory provision concerning the protection of the environment. (Republic of South Africa Government 1998, s. 2, ss. 1)
The application of these principles is binding on all organs of state. Within these, NEMA brings into South African law the public trust doctrine which is defined as:
The environment is held in public trust for the people, the beneficial use of environmental resources must serve the public interest and the environment must be protected as the people's common heritage. (Republic of South Africa Government 1998, s. 2, ss. 4o)
The remainder of the environmental principles provide the guidance and the tools necessary to ensure that the trust entity (the environment) is not depleted or degraded (Blackmore 2015a). The application of the public trust doctrine is further reinforced by section 3 of the National Environmental Management: Biodiversity Act (No. 10 of 2004) (NEMBA) and the National Environmental Management: Protected Areas Act (No. 57 of 2003) (NEMPAA) in which both biodiversity and protected areas are held, by the State, in trust for the people of South Africa, thereby clearly bringing biodiversity, and in this case the tsetse flies, into the public trust entity (the environment). Thus, the State has fiduciary duty to ensure, through all of its organs of state, the protection of the tsetse flies as species within South Africa as well as a component of biodiversity occurring within the country's protected areas (Blackmore 2014).
The EP is in conflict with the 2003 African Convention on the Conservation of Nature and Natural Resources (the Maputo Convention) that requires state parties to undertake measures, inter alia, to avoid or eliminate risks to biodiversity that manifest at species and habitat levels (Article X). Furthermore, the parties to the Convention are to ensure that they, inter alia:
to the maximum extent possible, take all necessary measures to ensure that development activities and projects are based on sound environmental policies and do not have adverse effects on natural resources and the environment in general. (Article XIV, s. 2.a; IUCN 2004)
The 1999 Southern African Development Community (SADC) Protocol on Wildlife Conservation and Law Enforcement requires each state party to apply their fiduciary duty to ensure that its wildlife resources are conserved and used sustainably (Southern African Development Community 1999, Article 3, s. 1.). The protocol also requires SADC countries to refrain from causing 'damage to the wildlife resources of other states or in areas beyond the limits of national jurisdiction' (Southern African Development Community 1999, Article 3, s. 1.). The request or expectation of South Africa to eradicate tsetse flies in this country, by neighbouring countries, would conflict with this provision (Blackmore 2015b).
The EP is also in conflict with the Convention on Biological Diversity and the Ramsar and World Heritage Conventions, to which South Africa is a signatory. Given that the South African distribution of tsetse flies includes protected areas that are Ramsar and WHS sites (Figure 1), the provisions of these conventions also apply. The global recognition of these sites is founded mainly on the species and habitat assemblages that occur within these protected areas. South Africa has an obligation to protect the Outstanding Universal Values of the iSimangaliso Wetland Park WHS: it fulfilled, inter alia, the criterion of containing 'the most important and significant natural habitats for in-situ conservation of biological diversity' (United Nations Educational, Scientific and Cultural Organisation 2015, s. II.D, ss. 77[x]); the prerequisite condition of integrity included the condition that 'The properties should contain habitats for maintaining the most diverse fauna and flora characteristic of the bio-geographic province and ecosystems under consideration' (United Nations Educational, Scientific and Cultural Organisation 2015, s. II.E, ss. 95). It is thus common cause that the WHS would include a complete assemblage of co-evolved animals, including indigenous mammals, tsetse flies and trypanosomes.
By entering into these multilateral agreements, and thereafter having applied for both listing and the inscription of these protected areas, the State has fundamentally bound itself to conserving and protecting all indigenous species and habitats that comprise the biodiversity of these sites (section 231 of the Constitution). Furthermore, these multilateral agreements have also been brought into South Africa's jurisprudence by way of the NEMA (section 2[4][n]) and the NEMBA (section 5), and within an array of explicit provisions in the NEMPAA. The NEMPAA further requires the State to ensure that the management of a protected area is commensurate with the purpose it was declared as such (Blackmore 2015a, 2015b). Finally, by way of a schedule to the World Heritage Convention Act (No. 49 of 1999), the Convention is explicitly brought into South African law. The entire Convention and all provisions of this Convention are, therefore, binding on all organs of state. Even outside protected areas, the State has a duty to conserve indigenous biodiversity.
Tsetse flies and trypanosomes are part of biodiversity
Tsetse flies and trypanosomes are ancient and remarkable organisms, being components of indigenous biodiversity, and have intrinsic existence value. A maximum of about nine final instar larvae are born live per female and so tsetse flies are 'K-selected' (Nagel 1995), rather like humans and elephants. They are the only insects that live entirely on blood, and they have become model organisms in biology (Snyder & Rio 2013). Trypanosomes are remarkable for their ability to live in two hosts, and to evade the mammalian immune system, mediated through changes in their surface glycoproteins (Matthews 2005). The trypanosome has become a model organism in the study of cell biology and genetics.
Ecosystem health and functioning would be compromised
Indigenous parasites and their vectors are important components of biodiversity. Tsetse flies are eaten by many animals (Nagel 1995), and so if they are eradicated, the food-web structure will be altered, which would result in a less stable and resilient animal community. The elimination of a parasite from an ecosystem can strongly affect the interactions between a diverse range of species in the community, both hosts and non-hosts, and hence affect biodiversity (Lebarbenchon et al. 2007). Parasites such as trypanosomes are an integral part, and are vital to the maintenance, of biodiversity. Parasitic species increase the species richness of an area dramatically, and parasites alter the outcomes of competition between, and therefore the relative abundance of, other animal species.
Parasites increase the number of linkages in food webs and therefore the connectivity and cohesion of the food webs, as well as increase the average number of links per species and the lengths of food chains. These factors increase the stability of food webs and therefore of biotic communities. Animals at higher trophic levels are less vulnerable to predators but more vulnerable to parasites. All of these factors cause an increase in ecosystem stability (Lafferty, Dobson & Kuris 2006). Parasites can divert host energy towards investment in immune function, reduce reproductive success and increase susceptibility to predators or to stress. Many parasites have significant effects on the survival, behaviour, growth and competitive ability of their animal hosts, and therefore play a role in natural population regulation and have an important influence on the flow of energy between various trophic levels. Parasites influence speciation through changes in animal life histories and gene flow and by influencing the ways in which species interact. The end result is that parasites increase the health of ecosystems (Hudson, Dobson & Lafferty 2006).
The following local example is illustrative of a role of tsetse flies and trypanosomes in ecosystems. R.H.T.P. Harris discovered in Zululand in the 1920s that shape and shading were important in the attraction of tsetse flies to their hosts. A unicoloured mammal of horizontal, cylindrical shape with its undersurface in shadow attracts tsetse flies; the horizontal cylindrical body profile of a zebra is 'broken up' by vertical black and white contrasting stripes, rendering them unattractive to biting flies, and they are seldom bitten. Therefore, the characteristic striping of zebras evolved to avoid the attention of biting flies, especially tsetse flies (Caro et al. 2014; Egri et al. 2012; Waage 1981). Classic equine symptoms of trypanosomiasis are known in zebras, and the disease may come at a greater fitness cost than for wildebeest (Caro et al. 2014; Ford 1971; Glover 1965; Neitz 1931; Rurangirwa et al. 1986). Therefore, the striped coats of zebras appear to have an adaptive function to reduce the risk of parasitism by trypanosomes.
Direct pathological effects of trypanosomes on populations of wild mammals will most likely occur when animals are stressed (Ford 1971; Glover 1965; Molyneux 1982), such as in times of drought and poor food availability or high competition with other species (cf. Melton 1987). In a prolonged drought situation, reduction in population numbers may be crucial to avoid severe habitat degradation (Walker et al. 1987). Trypanosomes may play a role in reducing populations of certain species more quickly under these circumstances.
Non-target organisms would be detrimentally impacted
The OD study documented that the SAT had immediate, significant, measurable impacts on non-target organisms, some of which were long lasting (Perkins & Ramberg 2004; Ramberg et al. 2006). Abundance of aquatic invertebrates and terrestrial invertebrates was reduced by up to 50% and 70%, respectively, and the losses of higher aquatic invertebrate taxa and of species in the canopies of various tree species was up to 30% and 40%, respectively, after spraying. There was a significant effect of deltamethrin spray on the abundance and community composition of non-target invertebrate organisms (Perkins & Ramberg 2004). Although recovery of assemblages to near those present pre-spray was documented in the recovery study of 2003, some of the spray-affected families of aquatic organisms remained at reduced levels and 10% of the identified species may have been lost because of spraying. In the terrestrial invertebrate recovery study, the long-term fate of 18 rare species flagged as missing could not be determined. However, only four tree species out of the approximately 1300 vascular plant species and subspecies in the OD were sampled and five major invertebrate groups analysed (Perkins & Ramberg 2004; Ramberg et al. 2006). Given that many invertebrates are host specific, even at lower taxonomic levels, or are microhabitat specific, the long-term impact of the SAT on the total terrestrial invertebrate assemblage of the OD could not be determined from such a small sample.
Little follow-up study was done in the OD to determine whether any 'lost' species may have been extirpated from the sprayed area. For example, various Odonata were severely affected by the aerial spraying and the disappearance of 22 species of dragonflies and damselflies from the Delta was plausibly the result of the aerial spraying (Ramberg et al. 2006). Kipping (2010) recorded some of the 'lost' species again in the OD, as would be expected for non-endemic mobile species, but some species historically recorded in the OD were not recorded after the spraying. Kurugundla, Kgori and Moleele (2012) summarised the impacts of the SAT using deltamethrin that was carried out in the Kwando-Linyanti region in 2006, which included severe detrimental impacts on orthopterans.
The biodiversity of the north-eastern KwaZulu-Natal is more susceptible to the sequential aerosol technique than that of the Okavango Delta
Although the proposed number of insecticide sprays is fewer than in the OD because of the proposal to use the SIT in conjunction with insecticide sprays, severe negative impacts can still be expected if the EP were to be implemented. The OD is part of a vast, relatively uniform savanna, and generally the species there are widely distributed. No species were known to be endemic to the OD (Cowling & Hilton-Taylor 1997; Ramberg et al. 2006). The re-appearance of hardy and mobile species after the spraying could be reasonably expected. In contrast, Maputaland is hemmed in by the sea on one side and by mountains on another, and consists of a diverse mosaic of habitats. The Maputaland Centre of Endemism is part of the Maputaland-Pondoland-Albany global biodiversity hotspot (Cowling & Hilton-Taylor 1997; Steenkamp et al. 2004). The negative impacts of the SAT would likely be greater in Maputaland than in the OD owing to greater habitat heterogeneity, environmental stability, numbers of localised endemics and perhaps more specific relationships between certain invertebrates and particular host plant species or localised vegetation types. For example, moth species assemblages differ between Ndumo Game Reserve, Tembe Elephant Park, Manguzi Forest Reserve and Kosi Bay which are all in relatively close proximity (Figure 1; Staude 2016; see Staude 1999 for other comparisons). Seventy-three per cent of the total 322 species recorded in these protected areas have only been found in one reserve. Conversely, in the northern Kalahari, there is very little change in looper moth composition between Shakawe and Maun, which are about 250 km apart (Staude 2016). Re-establishment of populations of species that were vulnerable to deltamethrin would depend on nearby sources of immigrants and the lack of barriers (e.g. Longley et al. 1997). These conditions would not be fulfilled for Maputaland endemics and likely not for species with patchy distributions in a transformed and fragmented landscape.
Extirpation of non-target organisms in north-eastern KwaZulu-Natal: Extermination of tsetse flies can have repercussions for other elements of biodiversity. Predators of tsetse flies may be forced to feed on other prey (Nagel 1995) and may be reduced in number. Parasites (including trypanosomes) and parasitoids of tsetse flies may be concomitantly exterminated or reduced in number. Parasitoids and predators are important for the control of tsetse and other fly populations (Glasgow 1963). The mourning bee fly Exhyalanthrax lugens, which parasitises tsetse fly pupae, was eradicated from areas of neKZN that were aerially sprayed with DDT even before the pale-footed tsetse fly Glossina pallidipes itself was eradicated (Fiedler, Du Toit & Kluge 1954; Fiedler & Kluge 1954). This bee fly did not return to the sprayed areas, nor was it and another parasite of tsetse fly pupae, the abrupt bee fly Exhyalanthrax abruptus, recorded at the uMkhuze Game Reserve in an extended survey carried out between 2002 and 2005 (Greathead et al. 2006). This suggests that the spraying had long-term impacts on some non-target species and that the control of other tsetse flies may have been affected, perhaps allowing populations of other tsetse fly species to expand (cf. Esterhuizen et al. 2005). Parasitoids endemic to Maputaland (Figure 2) will be susceptible to reduction in numbers and eventual elimination, depending on their specificity to the tsetse fly hosts and sensitivity to the SAT. Ecological processes such as the population control of other flies may therefore be affected.
Additionally, some lepidopteran species disappeared from the sprayed area to the south and east of the Lebombo Mountains. For example, Rhodometra satura (Figure 3) has not been seen again in the sprayed areas despite the presence of appropriate habitat, host plants and populations to the north. Chionopora tarachodes (Figure 3) is one of several other examples of Maputaland-endemic looper moths that may be vulnerable to SAT (Staude 2016).
Endemic invertebrates on the Red List: Some Rare, non-endemic invertebrate species were present in the OD before the commencement of the SAT in 2001, but no threatened endemics (Ramberg et al. 2006). In contrast, the Critically Endangered Zulu ambush katydid Peringueyella zulu, the Endangered Zululand black millipede Doratogonus zuluensis, the Endangered orange wisp damselfly Agriocnemis ruberrima, the vulnerable Umsingazi sprite damselfly Pseudagrion coeleste umsingaziense (Figure 4) and the vulnerable blue river crab Potamonautes lividus are endemic or near-endemic to neKZN and the wider Maputaland region (IUCN 2016; Samways 2006). Most, if not all, of these species would be severely affected were the EP implemented. Some Rare butterfly species, including the white spotted sapphire Iolaus lulua (Figure 4), Pennington's white mimic Ornipholidotos peucetia penningtoni and Zulu buff Teriomima zuluana, that complete their whole life-cycles on plants, are also endemic to the same region (eds. Mecenero et al. 2013). Deltamethrin is known to cause mortality of butterfly larvae at very low doses, including relatively long-term delayed mortality of larvae and pupae through sublethal effects, as well as loss in fitness in survivors (Çigli & Jepson 1995). Other non-endemic Red List species occur in neKZN. The threat status of most invertebrate groups in South Africa is unknown, but some species have their entire life-cycles in the canopy of trees or on bushes in the region, and these may be susceptible to local extinction if the EP were to be implemented.
Some examples of other susceptible invertebrates: Dragonflies and damselflies, beetles, grasshoppers and crickets, and spiders were some groups negatively affected by the SAT in the OD (Kurugundla et al. 2012; Perkins & Ramberg 2004; Ramberg et al. 2006). Owing to the diversity of habitats, neKZN is an important area for Odonata (50% of the species in South Africa are known to occur there) which contribute to various ecosystem services (Hart et al. 2014). Implementation of the proposed SAT would have negative repercussions for Odonata. Amongst the beetle families in neKZN, leaf beetles (Family Chrysomelidae) may be severely affected by the SAT. Many species in the subfamilies Chrysomelinae and Cassidinae have traits that make them very vulnerable to heavy mortality or extinction through aerial application of insecticides, including traits such as host specificity (often at plant species or genus level), living exposed on plants for most or all life-cycle stages and winglessness (Chaboo 2007; Grobbelaar 2016; Heron & Borowiec 1997; Figure 5). Not only do leaf beetles contribute to ecosystem functioning through their herbivory, but many good flyers are also pollinators. Orthoptera are important herbivores in the savannas (Scholes & Walker 1993), and at least seven genera of grasshoppers and three genera of crickets have flightless species endemic or near-endemic to Maputaland. Spiders are very important predators of other invertebrates, and some spiders that live above-ground, including on tree trunks and on grass, are known to be sensitive and even very sensitive to deltamethrin (Nagel 1995). Maputaland has a high diversity of spiders (e.g. Haddad & Russell-Smith 2009). Declines in populations of at least some species of spider, if not extinction, would occur if the EP were to be implemented, and non-lethal effects would affect invertebrate community structure and ecological processes to some degree.
The negative effects of factors such as climate change on the persistence of elements of biodiversity may either be added to or multiplied with the negative effects of other factors, such as habitat loss (Jewitt et al. 2015a), increased pollution, increased use of pesticides and drought, to cause a greater negative effect than if just one factor was operating on biodiversity. The impact of insecticide spray, acting additively or synergistically with other factors, may push some species towards extinction or lead to local extirpation through decrease in population size, including through various genetic effects in small and isolated populations (e.g. Pekin 2013). Conservation is required in various parts of the range of a species, particularly for endemic species that occur at low densities and in restricted habitats.
Negative ecosystem effects are likely
Negative ecosystem effects in the aquatic environment
The use of deltamethrin in the OD affected the full spectrum of aquatic habitats and animals, from water surface predators to sediment-dwelling mayflies, with negative impacts to a wide range of non-target aquatic organisms, representing many of the functional feeding groups of aquatic macroinvertebrates (Perkins & Ramberg 2004). Community recovery was affected, with some changes to community structure and function. A good example of impacts from the OD study concerns freshwater shrimps Caridina species that were severely negatively affected by the deltamethrin applications and showed poor recovery. Freshwater shrimps (Figure 6) are important in various freshwater lakes in neKZN: they are the dominant zoobenthic species of the littoral zone of Lake Sibaya in terms of biomass, and occur in nearly every marginal habitat, and are abundant in various habitats at Lake Mgobezeleni (Bruton 1980; Hart 1979, 1980). These detritivores will be important in the process of the mineralisation of detritus. Lake Sibaya and many of the wetlands in neKZN are closed systems, being rain fed and with drainage internal to the region, so the shrimps will not be able to re-colonise them from elsewhere should the EP be implemented.

In Lake Sibaya and in pans on the Pongolo River floodplain, Caridina was found to be a main food in the diet of the tank goby Glossogobius giurus, manyspined climbing perch Ctenopoma multispine and smaller sharp-tooth catfish Clarias gariepinus, and together with aquatic insects form part of the diet of other fish such as young tigerfish Hydrocynus vittatus, silver catfish Schilbe intermedius, snake catfish Clarias theodore, imberi Alestes imberi, southern redbreast tilapia Tilapia rendalli and adult Mozambique tilapia Oreochromis mossambicus (Bruton 1979; Bruton & Kok 1980; Kok 1980). Deltamethrin is known to be toxic to even hardy fish, and can cause mortality of, or non-lethal effects in, fish (e.g. Huang et al. 2014). Cascading negative ecosystem effects might occur in neKZN should the EP be implemented.
Impact on livelihoods of poor rural people: Fish are important in the diets of poor rural people in Maputaland, and the fish species mentioned in the preceding paragraph are important for the local fishermen (Bruton & Kok 1980; Coetzee et al. 2015; Kok 1980). The livelihoods of these poor rural people in the spray area would likely be negatively impacted were the EP to be implemented, through impact on the prey base of fish species that are important in their diets. Actions that reduce the provision of ecosystem services must be properly weighed up against the benefits of the actions.
Negative ecosystem effects in the terrestrial environment
Invertebrates play very important roles in the functioning of ecosystems. Implementation of the EP will have impacts on ecological processes and ecosystem services, as illustrated by the selected examples below.
Nutrient (carbon, nitrogen and phosphorus) cycling from leaves to the soil: Herbivorous invertebrate communities play very important roles in the recycling of carbon, nitrogen and phosphorus from vegetation to the soil, making these nutrients more readily available to plants and other micro-organisms than would otherwise happen (Metcalfe et al. 2013). The detrimental impacts that the EP would have on insect herbivore communities could negatively affect ecosystem carbon sequestration and soil fertility, particularly in sub-tropical forests and grasslands, at least in the short term. Scholes and Walker (1993) reported that leaf-feeding grasshoppers and lepidopteran larvae removed about 5% of the foliar primary production in an infertile, sub-tropical, broad-leafed savanna, and a greater proportion of foliar primary production in fertile sub-tropical, fine-leafed savanna. This biomass is returned to the soil as faeces, bodies and cut pieces of leaves that are more easily processed by detritivores and bacteria than abscised leaves. The plant nutrients are thereby more efficiently recycled to the soil to assist plant growth.
Pollination of flowering plants: Reduction in pollination efficiency may occur if non-target pollinators (such as bees, bee flies, beetles and tabanid flies) are reduced in number or extirpated. Pollination limitation may lead to a reduction in or absence of recruitment of certain plant species through poor or absent seed production (Potts et al. 2010), a reduction in the preferred food supply of certain herbivores, and so on. Some wild pollinators, particularly bees, have been declining globally, in part owing to the lethal and sublethal effects of insecticides, which may act synergistically with other drivers such as habitat loss and disease (Potts et al. 2010). An estimated 94% of plant species in the latitudinal band 29° - 0° are biotically pollinated, mainly by insects (Ollerton, Winfree & Tarrant 2011), and the impact on the pollination of some of the plant species of Maputaland could be severe if the EP were to be implemented. Vamosi et al. (2006) show that animal-pollinated plants in plant hotspots have reduced seed-sets owing to greater pollen-limitation than elsewhere, and human-mediated impacts on pollinators may eventually lead to the extinction of plant species. Bees may be killed or disabled by deltamethrin, depending on the genus or species of bee (Desneux, Decourtye & Delpuech 2007; Scott-Dupree, Conroy & Harris 2009). Other pollinating insect groups have not been studied the way bees have been, but the same general impacts are likely. Toxicity of a particular insecticide to different pollinators can vary markedly. However, Scott-Dupree et al. (2009) found that deltamethrin is highly toxic to alfalfa leaf-cutting bees Megachile rotundata, an important indigenous North American pollinator of native plants and cultivated crops. At least 20 species of Megachile bees occur in neKZN (Eardley 2013 and references therein) and deltamethrin may be highly toxic to them.
Parasitism and predation: Parasitoid and predator insects are important for controlling insect populations. Deltamethrin is known to have lethal and a wide variety of sublethal effects on insect parasitoids and predators (e.g. Desneux et al. 2007; Longley et al. 1997). A small change in the reproductive potential of parasitoids can disrupt biological control and make it less successful. Deltamethrin can act synergistically with another pesticide in terms of detrimental effects (Desneux et al. 2007).
Ecosystem services relied on by poor rural people: Implementation of the EP would affect ecosystem services needed by, or important to, poor rural people. Poor rural communities in the proposed spray region are reliant on indigenous plants to maintain health and treat disease, and use indigenous plants and other vegetables and fruits as food (Cunningham 1988a, 1988b; Ellis, Myers & Ricketts 2015). Many of these are insect-pollinated and a variety of wild insect pollinators are vital for good fruit production in many pollinator-dependent vegetable and fruit crops (Garibaldi et al. 2011, 2013). Managed honeybee pollination cannot replace wild insect pollination for many of these crops. Predatory insects and insect parasitoids that assist with the control of pests of crops (e.g. Samways 1988), including those of poor rural people, would be adversely affected by broad-scale SAT using deltamethrin, with likely detrimental impacts on crop production, food security and traditional medicine resources. Crop production may be affected where poor rural people rely on ecosystem services rather than fertilisers for soil fertility. Many rural people in neKZN rely directly or indirectly on activities associated with wildlife for income (Aylward & Lutz 2003). Protected areas are reservoirs of natural resources, including invertebrates that mediate ecosystem processes and services.
Biological control: Alien plant biological control projects in neKZN might be negatively affected were the EP implemented. One of the probable reasons why neKZN does not have great problems with feral and alien invasive mammals is that these mammals are susceptible to diseases such as trypanosomiasis. If trypanosomes, and in future other disease-causing parasites and their vectors, are deliberately exterminated in neKZN, feral and alien invasive mammal problems would likely increase, with corresponding increased impacts on indigenous species. Various domesticated animals that have the ability to become feral are susceptible to trypanosomiasis (Nwoha 2013), and when controlling factors for those species are reduced or eliminated, the feral species are released from population control and can cause changes in community structure and function, including extinctions of indigenous species (Hollings et al. 2016). Controlling feral and alien mammals would be costly.
Stable host-parasite relationships: Eradication of tsetse flies and certain trypanosome species would disrupt the stable host-parasite relationship between the trypanosomes and the native mammalian wildlife. Should these tsetse flies and trypanosomes return to the region after a certain period, severe clinical trypanosomiasis would be likely in wild mammals. Immunity against local trypanosomes but not against foreign trypanosomes is a known phenomenon (Ford 1971). South African rhinos that were translocated to areas where different trypanosomes were encountered, and rhinos brought into Maputaland from areas that do not have trypanosomes, suffered from trypanosomiasis. Some died and others had to be treated. Also, if tsetse fly parasites have been eliminated, tsetse fly populations could build up rapidly again. The reasoning that tsetse flies would not come back to the region if the whole population was eradicated can also be used to support the claim that the eradication would then be unnecessary as neither would the population expand northwards.
The eradication of trypanosomes without proper cattle population controls would likely exacerbate the destruction of biodiversity
Forty-four vegetation types occur in the approximate area of occurrence of tsetse flies in KwaZulu-Natal. Ten of these vegetation types are classed as Critically Endangered, eight as Endangered, five as vulnerable and 21 as Least Threatened. Targets for protection have been reached in only 14 of the 44 vegetation types; two of the Critically Endangered vegetation types are not protected. Existing protected areas fall short of conserving a representative sample of the diversity of the region, and accordingly emphasise the importance of areas outside protected areas for meeting South Africa's and Mozambique's and Swaziland's conservation commitments (Smith et al. 2008). The rate of land transformation is high at an average of 1.2% of KwaZulu-Natal per annum between 1994 and 2011 (Jewitt et al. 2015b). Therefore, reaching the conservation targets in the future is becoming less likely. Risks associated with tsetse fly eradication include increasing cattle and other livestock numbers, leading to a reduction of vegetation cover, increasing runoff and erosion, increased pressure on conserved areas, increased transmission of livestock diseases to wild mammals and vice versa, and reduction in biodiversity and increase in alien plants through overgrazing. Contrary to the EP (IAEA 2006), the argument for not eradicating tsetse flies has in fact become stronger in this respect.
Environment-friendly methods and options for the control of trypanosomiasis in cattle
Alternative options currently exist to reduce the incidence and prevalence of trypanosomiasis in cattle without causing extinction of indigenous species, and other options will be available in the future.
Normal veterinary measures that include trypanosomiasis surveillance, dipping of cattle using appropriate dips or else the use of pour-ons and treatment of infected cattle with trypanocides have proved effective in the past (Kappmeier, Nevill & Bagnall 1998). Trypanosomiasis is not the only, nor the most important, disease in cattle in neKZN. Tick-borne diseases also have to be controlled using the same methods. Decentralised control of animal husbandry at the farmer level using environment-friendly methods could be the ultimate goal. Rather than area-wide eradication, creation of local grazing areas free of tsetse flies would assist the farmer and would be more environment-friendly.
For many centuries, communities practicing agriculture have been able to reside successfully in areas where tsetse flies existed via local adaptations in the ways the communities formed and lived (e.g. Anderson et al. 2015; Ford 1971). Colonialism and the impacts of its practices and pastoralist migrations often had adverse impacts on these ways of life (e.g. Anderson et al. 2015; Ford 1971). The consequences of the gradual encroachment of people and cattle into an area where tsetse flies occur on the epidemiology of bovine trypanosomiasis are well known (Molyneux 1982; Van Den Bossche 2001). Hunting of indigenous mammals and continuous clearing of vegetation eventually removed the hosts and breeding habitat of the tsetse flies, which then disappeared from the area. This enabled farming with crops and cattle to occur even within the tsetse belts (Ford 1971; Kent 1926; Van Den Bossche 2001). Trypanosomiasis in cattle is likely to be more severe where the cattle are close to the boundaries of protected areas or other areas of suitable tsetse fly habitat with wild mammal hosts (De Beer et al. 2016; Van Den Bossche 2001). Glossina austeni has a short flight distance and is predominantly restricted to forest in KwaZulu-Natal (Esterhuizen et al. 2005). Keeping cattle away from forest or bush would greatly reduce the incidence of trypanosomiasis in cattle. This could be achieved by grazing cattle on fenced pastures and by keeping cattle out of protected area buffers or away from protected area boundaries. Trypanosomiasis in cattle is more severe where poor nutrition or other stress factors are present (Ford 1971; Holmes 2013). Improvement in the immune system and general health of cattle will occur with improvement in the condition of grazing lands and maintenance of these in good condition through rehabilitation of grazing lands, adherence to appropriate stocking and land resting rates, and implementation of an integrated land-use plan with designated grazing areas (e.g. pastures) where tsetse flies cannot breed. Traps and targets are effective for tsetse flies in KwaZulu-Natal (Esterhuizen et al. 2006; Kappmeier & Nevill 1999), and so could be used to good effect on cattle grazing lands that are within the normal flight range of tsetse flies, especially Glossina brevipalpis. Certain species of trypanosomes such as Trypanosoma vivax are transmitted via the proboscis of blood-feeding flies, including tsetse flies, horse flies (Family Tabanidae) and stable flies (Genus Stomoxys). So the eradication of tsetse flies alone will not prevent trypanosomiasis in cattle (Ahmed et al. 2016).
Replacing susceptible cattle by cattle with resistance to trypanosomes in a recapitalisation programme would be fruitful (Department of Agriculture 1950; Holmes 2013). Maintenance of heterozygosity by rotation of breeding bulls should be practiced to decrease susceptibility of calves to transmittable diseases such as trypanosomiasis (Murray et al. 2013). Maintenance of physiological adaptation of the cattle to trypanosomes, as occurred in Nguni cattle (Department of Agriculture 1950), would be required. Stress may cause cattle to lose resistance, so it may be necessary to use other means than cattle for ploughing. Tractors are alternatives to cattle for draught power and are preferred in terms of modernisation of agriculture. In addition, modern agricultural thinking emphasises the value of no-till farming which has grown remarkably in KwaZulu-Natal because of its benefits for the farmer and carbon storage. Cattle coat colour selection could be used to good effect. White horse models are very unattractive to tabanid flies (Egri et al. 2012), and blue and black are most attractive to tsetse flies in neKZN (Kappmeier & Nevill 1999). If the majority of Nguni cattle near the boundaries of protected areas had light coats, the incidence of trypanosomiasis may be reduced further. White-coloured cattle were kept by the king of the Mtetwa clan, King Dingiswayo, in the iMfolozi Game Reserve area before it was proclaimed a reserve (Foster 1955). Perhaps this was a local adaptation to the presence of tsetse flies and horse flies, the white coats of the cattle not being attractive to them. Maputaland is not South Africa's major beef-producing region, and nature tourism plays a major economic and development role in neKZN (eds. Aylward & Lutz 2003). The dependence on cattle as a monetary system could be reduced through diversifying income streams, including small business development around wildlife or the green economy.
Environmental costs (many of which are not financial) have not been quantified or incorporated into a cost-benefit analysis in terms of the EP. Further, opportunity costs, depending on what are the pressing social needs, and costs and benefits of alternative management strategies would also have to be included. A significant contribution to livestock productivity could be achieved by investing in the control of invasive alien plants that are invading and reducing the grazing capacity in neKZN, such as famine weed Parthenium hysterophorus. With the sequencing of the trypanosome genome complete, and genetic research on trypanosomes proceeding well, there is a good chance that biochemical and molecular genetic techniques can be used to prevent or reduce trypanosomiasis in cattle in the future (e.g. Caljon, De Vooght & Van Den Abbeele 2013). The development of vaccines could be one outcome. Owing to the great strides that are being made in understanding the biology of trypanosomes, long-lasting environment-friendly solutions for the prevention of trypanosomiasis in cattle are likely.
Conclusion
The proposed mass killing of invertebrates, including parasites, as mooted in the tsetse fly EP, would be illegal and detrimental to the biodiversity of South Africa, as was the destruction of 'game' in the middle of the 20th century. Eradication of tsetse flies from protected areas and the use of aerial spraying of insecticides over vast regions is no longer an option in a country that prides itself on its conservation principals with innovative and advanced environmental protection laws. A total ban on insecticide use and the SIT is not what is being proposed here, but rather conservative and environmentally judicial use where needed at local grazing land scales, if appropriate. A variety of techniques are now available to combat trypanosomiasis in cattle, and in future others should become available. The more environment-friendly ones should be prioritised for use at the grazing land scale.
Acknowledgements
We thank Ezemvelo KZN Wildlife for assisting with the research and publication of this work. The ideas, arguments and opinions expressed in this manuscript are that of the authors and do not necessarily represent those of Ezemvelo KZN Wildlife. We thank the following people for their valuable contributions to this article: Beth Grobbelaar for information relating to leaf beetles and for comments on the manuscript, Camille Lebarbenchon for information on the role of parasites in ecosystems, Hermann Staude for information on and photographs of looper moths in north-eastern KwaZulu-Natal, Nick Rivers-Moore for assistance with aquatic aspects of the article, Roger Porter for information on World Heritage Site matters, and Michael Samways and Steve Woodhall for permitting the use of their photographs. We also thank our colleagues Debbie Jewitt, Dave Cooper, Ian Rushworth and John Craigie for information on vegetation types, trypanosomes in non-resistant rhinos, alien plant invasion and for providing a photograph, respectively.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
A.J.A. was the project leader and conceptualised, designed and wrote the review. A.B. contributed the legal section.
References
Ahmed, S.K., Rahman, A.H., Hassan, M.A., Salih, E.M., Paone, M. & Cecchi, G., 2016, 'An atlas of tsetse and bovine trypanosomosis in Sudan', Parasites & Vectors 9, 194-201. http://dx.doi.org/10.1186/s13071-016-1485-6 [ Links ]
Anderson, N.E., Mubanga, J., Machila, N., Atkinson, P.M., Dzingirai, V. & Welburn, S.C., 2015, 'Sleeping sickness and its relationship with development and biodiversity conservation in the Luangwa Valley, Zambia', Parasites & Vectors 8, 224. http://dx.doi.org/10.1186/s13071-015-0827-0 [ Links ]
Ansara-Ross, T.M., Wepener, V., Van den Brink, P.J. & Ross, M.J., 2012, 'Pesticides in South African fresh waters', African Journal of Aquatic Science 37(1), 1-16. http://dx.doi.org/10.2989/16085914.2012.666336 [ Links ]
Aylward, B. & Lutz, E. (eds.), 2003, Nature tourism, conservation, and development in KwaZulu-Natal, South Africa, The World Bank, Washington, DC. [ Links ]
Blackmore, A., 2014, 'The interplay between the public trust doctrine and biodiversity and cultural resource legislation in South Africa: The case of the Shembe Church Worship Site in Tembe Elephant Park in KwaZulu-Natal', Law, Environment and Development Journal 10(1), 1-15. [ Links ]
Blackmore, A., 2015a, 'The relationship between the NEMA and the public trust doctrine: The importance of the NEMA principles in safeguarding South Africa's biodiversity', South African Journal of Environmental Law and Policy 20(2), 89-118. [ Links ]
Blackmore, A., 2015b, 'Legal and public trust considerations for the Ndumo Game Reserve and South Africa-Mozambique border, following the migration of the Usuthu River', Journal of Southern African Public Law 30(2), 347-379. [ Links ]
Bruton, M.N., 1979, 'The fishes of Lake Sibaya', in B.R. Allanson (ed.), Lake Sibaya, pp. 162-245, Dr W. Junk bv Publishers, The Hague. (Monographiae Biologicae 36). [ Links ]
Bruton, M.N., 1980, 'An outline of the ecology of the Mgobezeleni lake system at Sodwana, with emphasis on the mangrove community', in M.N. Bruton & K.H. Cooper (eds.), Studies on the ecology of Maputaland, pp. 408-426, Rhodes University, Grahamstown. [ Links ]
Bruton, M.N. & Kok, H.M., 1980, 'The freshwater fishes of Maputaland', in M.N. Bruton & K.H. Cooper (eds.), Studies on the ecology of Maputaland, pp. 210-244, Rhodes University, Grahamstown. [ Links ]
Caljon, G., De Vooght, L. & Van Den Abbeele, J., 2013, 'Options for the delivery of anti-pathogen molecules in arthropod vectors', Journal of Invertebrate Pathology 112(suppl. 1), 75-82. http://dx.doi.org/10.1016/j.jip.2012.07.013 [ Links ]
Caro, T., Izzo, A., Reiner, R.C., Walker, H. & Stankowich, T., 2014, 'The function of zebra stripes', Nature Communications 5, 3535. http://dx.doi.org/10.1038/ncomms4535 [ Links ]
Chaboo, C., 2007, 'Biology and phylogeny of Cassidinae Gyllenhal (tortoise and leaf-mining beetles) (Coleoptera: Chrysomelidae)', Bulletin of the American Museum of Natural History 305, 1-250. http://dx.doi.org/10.1206/0003-0090(2007)305[1:BAPOTC]2.0.CO;2 [ Links ]
Çigli, T. & Jepson, P.C., 1995, 'The risks posed by deltamethrin drift to hedgerow butterflies', Environmental Pollution 87, 1-9. [ Links ]
Coetzee, H.C., Nell, W., Van Eeden, E.S. & De Crom, E.P., 2015, 'Artisanal fisheries in the Ndumo Area of the lower Phongolo River Floodplain, South Africa', Koedoe 57(1), Art. #1248, 1-6. http://dx.doi.org/10.4102/koedoe.v57i1.1248 [ Links ]
Cowling, R.M. & Hilton-Taylor, C., 1997, 'Phytogeography, flora and endemism', in R.M. Cowling, D.M. Richardson & S.M. Pierce (eds.), Vegetation of Southern Africa, pp. 43-61, Cambridge University Press, Cambridge. [ Links ]
Cunningham, A.B., 1988a, 'An investigation of the herbal medicine trade in Natal/KwaZulu', Investigational report no. 29, Institute of Natural Resources, University of Natal, Pietermaritzburg. [ Links ]
Cunningham, A.B., 1988b, 'Collection of wild plant foods in Tembe Thonga society: A guide to Iron Age gathering activities?', Annals of the Natal Museum 29(2), 433-446. [ Links ]
De Beer, C.J., Venter, G.J., Kappmeier Green, K., Esterhuizen, J., De Klerk, D.G., Ntshangase, J. et al., 2016, 'An update of the tsetse fly (Diptera: Glossinidae) distribution and African animal trypanosomosis prevalence in north-eastern KwaZulu-Natal, South Africa', Onderstepoort Journal of Veterinary Research 83(1), a1172. http://dx.doi.org/10.4102/ojvr.v83i1.1172 [ Links ]
De Beer, C.J., Venter, G.J. & Vreysen, M.J.B., 2015, 'Determination of the optimal mating age of colonized Glossina brevipalpis and Glossina austeni using walk-in field cages in South Africa', Parasites & Vectors 8, 467-476. http://dx.doi.org/10.1186/s13071-015-1073-1 [ Links ]
Department of Agriculture, 1950, 'Nguni-cattle', Report on indigenous cattle in South Africa, Agricultural Research Institute series No. 22, Bulletin No. 311, The Government Printer, Pretoria. [ Links ]
Desneux, N., Decourtye, A. & Delpuech, J.-M., 2007, 'The sublethal effects of pesticides on beneficial arthropods', Annual Review of Entomology 52, 81-106. http://dx.doi.org/10.1146/annurev.ento.52.110405.091440 [ Links ]
Eardley, C., 2013, 'A taxonomic revision of the southern African leaf-cutter bees, Megachile Latreille sensu stricto and Heriadopsis Cockerell (Hymenoptera: Apoidea: Megachilidae)', Zootaxa 3601(1), 1-133. http://dx.doi.org/10.11646/zootaxa.3601.1.1 [ Links ]
Egri, Á., Blahó, M., Kriska, G., Farkas, R., Gyurkovszky, M., Åkesson, S. et al., 2012, 'Polarotactic tabanids find striped patterns with brightness and/or polarization modulation least attractive: An advantage of zebra stripes', The Journal of Experimental Biology 215, 736-745. http://dx.doi.org/10.1242/jeb.065540 [ Links ]
Ellis, A.M., Myers, S.S. & Ricketts, T.H., 2015, 'Do pollinators contribute to nutritional health?', PLoS One 10(1), e114805. http://dx.doi.org/10.1371/journal.pone.0114805 [ Links ]
Esterhuizen, J., Kappmeier Green, K., Marcotty, T. & Van Den Bossche, P., 2005, 'Abundance and distribution of the tsetse flies, Glossina austeni and G. brevipalpis, in different habitats in South Africa', Medical and Veterinary Entomology 19, 367-371. http://dx.doi.org/10.1111/j.1365-2915.2005.00582.x [ Links ]
Esterhuizen, J., Kappmeier Green, K., Nevill, E.M. & Van Den Bossche, P., 2006, 'Selective use of odour-baited, insecticide-treated targets to control tsetse flies Glossina austeni and G. brevipalpis in South Africa', Medical and Veterinary Entomology 20, 464-469. http://dx.doi.org/10.1111/j.1365-2915.2006.00650.x [ Links ]
Fiedler, O.G.H., Du Toit, R. & Kluge, E.B., 1954, 'The influence of the tsetse fly eradication campaign on the breeding activity of Glossinae and their parasites in Zululand', Onderstepoort Journal of Veterinary Research 26, 389-397. [ Links ]
Fiedler, O.G.H. & Kluge, E.B., 1954, 'The parasites of tsetse flies in Zululand with special reference to the influence of the hosts upon them', Onderstepoort Journal of Veterinary Research 26, 399-404. [ Links ]
Ford, J., 1971, The role of the trypanosomiases in African ecology. A study of the tsetse fly problem, Clarendon Press, Oxford. [ Links ]
Foster, W.E., 1955, History of the Umfolozi Game Reserve, Unpublished report, Natal Parks, Game and Fish Preservation Board, Pietermaritzburg. [ Links ]
Garibaldi, L.A., Aizen, M.A., Klein, A.M., Cunningham, S.A. & Harder, L.D., 2011, 'Global growth and stability of agricultural yield decrease with pollinator dependence', PNAS 108, 5909-5914. http://dx.doi.org/10.1073/pnas.1012431108 [ Links ]
Garibaldi, L.A., Steffan-Dewenter, I., Winfree, R., Aizen, M.A., Bommarco, R., Cunningham, S.A. et al., 2013, 'Wild pollinators enhance fruit set of crops regardless of honey bee abundance', Science 339, 1608-1611. http://dx.doi.org/10.1126/science.1230200 [ Links ]
Glasgow, J.P., 1963, The distribution and abundance of tsetse, Pergamon Press, Oxford. [ Links ]
Glover, P.E., 1965, Review and recent knowledge on the relationship between the tsetse fly and its vertebrate hosts, Fauna Protection Society, London. [ Links ]
Greathead, D., Lovell, S., Barraclough, D., Slotow, R., Hamer, M. & Herbert, D., 2006, 'An ecological and conservation assessment of the fauna of Bombyliidae (Diptera) occurring in the Mkuze, Phinda and False Bay reserves, KwaZulu-Natal, South Africa', African Invertebrates 47, 185-206. [ Links ]
Grobbelaar, E., 2016, Biology of Chrysomelidae: Cassidinae, Chrysomelinae and Galerucinae, Unpublished report to Ezemvelo KZN Wildlife, Pietermaritzburg. [ Links ]
Haddad, C.R. & Russell-Smith, A., 2009, 'Spider family composition and species richness patterns in two savannah habitats along the eastern coastal plain of Africa', African Journal of Ecology 48, 418-427. http://dx.doi.org/10.1111/j.1365-2028.2009.01128.x [ Links ]
Hart, L.A., Bowker, M.B., Tarboton, W. & Downs, C.T., 2014, 'Species composition, distribution and habitat types of Odonata in the iSimangaliso Wetland Park, KwaZulu-Natal, South Africa and the associated conservation implications', PLoS One 9(3), e92588. http://dx.doi.org/10.1371/journal.pone.0092588 [ Links ]
Hart, R.C., 1979, 'The invertebrate communities: Zooplankton, zoobenthos and littoral fauna', in B.R. Allanson (ed.), Lake Sibaya, pp. 108-161, Dr W. Junk bv Publishers, The Hague. (Monographiae Biologicae). [ Links ]
Hart, R.C., 1980, 'The aquatic invertebrates of Lake Sibaya', in M.N. Bruton & K.H. Cooper (eds.), Studies on the ecology of Maputaland, pp. 114-122, Rhodes University, Grahamstown. [ Links ]
Heron, H. & Borowiec, L., 1997, 'Host plants and feeding patterns of some South African tortoise beetles (Coleoptera: Chrysomelidae: cassidoid Hispinae)', Genus 8, 625-658. [ Links ]
Hollings, T., Jones, M., Mooney, N. & McCallum, H., 2016, 'Disease-induced decline of an apex predator drives invasive dominated states and threatens biodiversity', Ecology 97(2), 394-405. [ Links ]
Holmes, P., 2013, 'Tsetse-transmitted trypanosomes - Their biology, disease impact and control', Journal of Invertebrate Pathology, 112(suppl. 1), 11-14. http://dx.doi.org/10.1016/j.jip.2012.07.014 [ Links ]
Huang, Y., Zhang, J., Han, X. & Huang, T., 2014, 'The use of zebrafish (Danio rerio) behavioral responses in identifying sublethal exposures to deltamethrin', International Journal of Environmental Research and Public Health 11, 3650-3660. http://dx.doi.org/10.3390/ijerph110403650 [ Links ]
Hudson, P.J., Dobson, A.P. & Lafferty, K.D., 2006, 'Is a healthy ecosystem one that is rich in parasites?', Trends in Ecology and Evolution 21, 381-385. http://dx.doi.org/10.1016/j.tree.2006.04.007 [ Links ]
International Atomic Energy Agency (IAEA), 2006, Feasibility study - The creation of a tsetse-free zone in the Republic of South Africa and in the Southern Part of the Republic of Mozambique, International Atomic Energy Association, Vienna. [ Links ]
International Atomic Energy Agency (IAEA), n.d., Supporting a feasibility study to eradicate tsetse from Southern Mozambique, South Africa and Swaziland, viewed 17 October 2016, from https://www.iaea.org/projects/tc/raf5069 [ Links ]
IUCN, 2004, An Introduction to the African Convention on the Conservation of Nature and Natural Resources, IUCN, Gland, Switzerland and Cambridge, UK, viewed n.d., from https://portals.iucn.org/library/sites/library/files/documents/EPLP-056.pdf [ Links ]
IUCN, 2016, The IUCN Red List of Threatened Species. Version 2016-1, viewed 04 August 2016, from http://www.iucnredlist.org [ Links ]
Jewitt, D., Erasmus, B.F.N., Goodman, P.S., O'Connor, T.G., Hargrove, W.W., Maddalena, D.M. et al., 2015a, 'Climate-induced change of environmentally defined floristic domains: A conservation based vulnerability framework', Applied Geography 63, 33-42. http://dx.doi.org/10.1016/j.apgeog.2015.06.004 [ Links ]
Jewitt, D., Goodman, P.S., Erasmus, B.F.N., O'Connor, T.G. & Witkowski, E.T.F., 2015b, 'Systematic land-cover change in KwaZulu-Natal, South Africa: Implications for biodiversity', South African Journal of Science 111(9/10), Art. #2015-0019, 1-9. http://dx.doi.org/10.17159/sajs.2015/20150019 [ Links ]
Kappmeier, K. & Nevill, E.M., 1999, 'Evaluation of coloured targets for the attraction of Glossina brevipalpis and Glossina austeni (Diptera: Glossinidae) in South Africa', Onderstepoort Journal of Veterinary Research 66, 291-305. [ Links ]
Kappmeier, K., Nevill, E.M. & Bagnall, R.J., 1998, 'Review of tsetse flies and trypanosomosis in South Africa', Onderstepoort Journal of Veterinary Research 65, 195-203. [ Links ]
Kappmeier Green, K., Potgieter, F.T. & Vreysen, M.J.B., 2007, 'A strategy for an area-wide control campaign with an SIT component to establish a tsetse-(Glossina austeni and Glossina brevipalpis) free South Africa', in M.J.B. Vreysen, A.S. Robinson & J. Hendrichs (eds.), Area-wide control of insect pests, pp. 309-323, IAEA, Vienna. [ Links ]
Kent, C.C., 1926, 'Field notes on the Mfolozi Game Reserve', South African Journal of Natural History 6, 38-48. [ Links ]
Kipping, J., 2010, 'The dragonflies and damselflies of Botswana - An annotated checklist with notes on distribution, phenology, habitats and Red List status of the species (Insecta: Odonata)', Mauritiana (Altenburg) 21, 126-204. [ Links ]
Kok, H.M., 1980, 'Ecological studies of some important fish species of the Pongolo floodplain, KwaZulu, South Africa', PhD thesis, University of KwaZulu-Natal, Pietermaritzburg. [ Links ]
Kurugundla, C.N., Kgori, P.M. & Moleele, N., 2012, 'Management of tsetse fly using insecticides in Northern Botswana', in F. Perveen (ed.), Insecticides - Pest engineering, pp. 449-476, InTech, Rijeka, viewed 12 August 2016, from http://www.intechopen.com/books/insecticides-pest-engineering/management-of-tsetse-fly-using-insecticides-in-northern-botswana [ Links ]
Lafferty, K.D., Dobson, A.P. & Kuris, A.M., 2006, 'Parasites dominate food web links', PNAS 103, 11211-11216. http://dx.doi.org/10.1073/pnas.0604755103 [ Links ]
Lebarbenchon, C., Poulin, R., Gauthier-Clerc, M. & Thomas, F. 2007, 'Parasitological consequences of overcrowding in protected areas', EcoHealth 3, 303-307. http://dx.doi.org/10.1007/s10393-006-0067-z [ Links ]
Longley, M., Jepson, P.C., Izquierdo, J. & Sotherton, N., 1997, 'Temporal and spatial changes in aphid and parasitoid populations following applications of deltamethrin in winter wheat', Entomologia Experimentalis et Applicata 83, 41-52. http://dx.doi.org/10.1046/j.1570-7458.1997.00155.x [ Links ]
Matthews, K.R., 2005, 'The developmental cell biology of Trypanosoma brucei', Journal of Cell Science 118, 283-290. http://dx.doi.org/10.1242/jcs.01649 [ Links ]
Mecenero, S., Ball, J.B., Edge, D.A., Hamer, M.L., Henning, G.A., Kruger, M. et al., (eds.), 2013, Conservation assessment of butterflies of South Africa, Lesotho and Swaziland: Red List and Atlas, Saftronics (Pty) Ltd., Johannesburg & Animal Demography Unit, Cape Town. [ Links ]
Melton, D.A., 1987, 'Waterbuck (Kobus ellipsipyrmnus) population dynamics: The testing of a hypothesis', African Journal of Ecology 25, 133-145. http://dx.doi.org/10.1111/j.1365-2028.1987.tb01100.x [ Links ]
Metcalfe, D.B., Asner, G.P., Martin, R.E., Silva Espejo, J.E., Huaraca Huasco, W., Farfán Amézquita, F.F. et al., 2013, 'Herbivory makes major contributions to ecosystem carbon and nutrient cycling in tropical forests', Ecology Letters 17(3), 324-332. http://dx.doi.org/10.1111/ele.12233 [ Links ]
Molyneux, D.H., 1982, 'Trypanosomes, trypanosomiasis, and tsetse control: Impact on wildlife and its conservation', in A. Marcia Edwards & U. McDonnell (eds.), Animal disease in relation to animal conservation: Symposia of the zoological society of London, vol. 50, pp. 29-55, Academic Press, London. [ Links ]
Murray, G.G.R., Woolhouse, M.E.J., Tapio, M., Mbole-Kariuki, M.N., Thumbi, S.M., Jennings, A.E. et al., 2013, 'Genetic susceptibility to infectious disease in East African Shorthorn Zebu: A genome-wide analysis of the effect of heterozygosity and exotic introgression', BMC Evolutionary Biology 13, 246. http://dx.doi.org/10.1186/1471-2148-13-246 [ Links ]
Nagel, P., 1995, Environmental monitoring handbook for tsetse control operations, The Scientific Environmental Monitoring Group, Margraf Verlag, Weikersheim. [ Links ]
Neitz, W.O., 1931, Blood parasites of game in Zululand, Preliminary Report, 17th Report of the Director of Veterinary Services and Animal Industry, Union of South Africa, Pretoria. [ Links ]
Nwoha, R.I.O., 2013, 'A review on trypanosomosis in dogs and cats', African Journal of Biotechnology 12(46), 6432-6442. http://dx.doi.org/10.5897/AJB2013.12093 [ Links ]
Ollerton, J., Winfree, R. & Tarrant, S., 2011, 'How many flowering plants are pollinated by animals?', Oikos 120, 321-326. http://dx.doi.org/10.1111/j.1600-0706.2010.18644.x [ Links ]
Pekin, B.K., 2013, 'Effect of widespread agricultural chemical use on butterfly diversity across Turkish provinces', Conservation Biology 27(6), 1439-1448. http://dx.doi.org/10.1111/cobi.12103 [ Links ]
Perkins, J.S. & Ramberg, L. (eds.), 2004, 'Environmental recovery monitoring of tsetse fly spraying impacts in the Okavango Delta - 2003 Final report May 2004', The Harry Oppenheimer Okavango Research Centre, University of Botswana, Maun. [ Links ]
Potts, S.G., Biesmeijer, J.C., Kremen, C., Neumann, P., Schweiger, O. & Kunin, W.E., 2010, 'Global pollinator declines: Trends, impacts and drivers', Trends in Ecology and Evolution 25(6), 345-353. http://dx.doi.org/10.1016/j.tree.2010.01.007 [ Links ]
Ramberg, L., Hancock, P., Lindholm, M., Meyer, T., Ringrose, S., Sliva, J. et al., 2006, 'Species diversity of the Okavango Delta, Botswana', Aquatic Sciences 68, 310-337. http://dx.doi.org/10.1007/s00027-006-0857-y [ Links ]
Republic of South Africa Government, 1998, National Environmental Management Act 107, Republic of South Africa Government, Pretoria, viewed n.d., from https://www.environment.gov.za/sites/default/files/legislations/nema_amendment_act107.pdf [ Links ]
Rurangirwa, F.R., Musoke, A.J., Nantulya, V.M., Nkonge, C., Njuguna, L., Mushi, E.Z. et al., 1986, 'Immune effector mechanisms involved in the control of parasitaemia in Trypanosoma brucei-infected wildebeest (Connochaetes taurinus)', Immunology 58, 231-237. [ Links ]
Samways, M.J., 1988, 'A pictorial model of the impact of natural enemies on the population growth rate of the scale insect Aonidiella aurantii', South African Journal of Science 84, 270-272. [ Links ]
Samways, M.J., 2006, 'National Red List of South African Odonata', Odonatologica 35, 341-368. [ Links ]
Scholes, R.J. & Walker, B.H., 1993, An African Savanna: Synthesis of the Nylsvley study, Cambridge University Press, Cambridge. [ Links ]
Scott-Dupree, C.D., Conroy, L. & Harris, C.R., 2009, 'Impact of currently used or potentially useful insecticides for canola agroecosystems on Bombus impatiens (Hymenoptera: Apidae), Megachile rotundata (Hymentoptera: Megachilidae), and Osmia lignaria (Hymenoptera: Megachilidae)', Journal of Economic Entomology 102(1), 177-182. http://dx.doi.org/10.1603/029.102.0125 [ Links ]
Smith, B. & Nhancale, B., 2010, 'Appendix 1: A conservation assessment for the Lubombo Transfrontier Conservation Area', in B. Smith (ed.), Final report: TFCA data collation and data gap analysis, pp. 1-23, Report to TFCA - CU Conservation Planning Systems (CPS) for the Chimanimani, Great Limpopo and Lubombo TFCAs, Durrell Institute of Conservation and Ecology, University of Kent. [ Links ]
Smith, R.J., Easton, J., Nhancale, B.A., Armstrong, A.J., Culverwell, J., Dlamini, S.D. et al., 2008, 'Designing a transfrontier conservation landscape for the Maputaland centre of endemism using biodiversity, economic and threat data', Biological Conservation 141, 2127-2138. http://dx.doi.org/10.1016/j.biocon.2008.06.010 [ Links ]
Snyder, A.K. & Rio, R.V.M., 2013, 'Interwoven biology of the tsetse holobiont', Journal of Bacteriology 195(19), 4322-4330. http://dx.doi.org/10.1128/JB.00487-13 [ Links ]
Southern African Development Community, 1999, Protocol on Wildlife Conservation and Law Enforcement, Southern African Development Community, Maputo, viewed n.d., from http://www.sadc.int/documents-publications/show/813 [ Links ]
Staude, H., 2016, An analysis of the moth species composition of the northern KwaZulu-Natal Reserves: Ndumo Game Reserve, Tembe Elephant Park, Manguzi Forest Reserve and Kosi Bay Reserve, indicating a high degree of local endemism, Unpublished report to Ezemvelo KZN Wildlife, Pietermaritzburg. [ Links ]
Staude, H.S., 1999, 'An illustrated report of 510 geometrid moth taxa (Lepidoptera: Geometridae) recorded from 28 protected areas from the northern and eastern parts of South Africa', Metamorphosis 10, 97-148. [ Links ]
Steenkamp, Y., Van Wyk, B., Victor, J., Hoare, D., Smith, G., Dold, T. et al., 2004, 'Maputaland-Pondoland-Albany', in R.A. Mittermeier, P.R. Gil, M. Hoffmann, J. Pilgrim, T. Brooks, C.G. Mittermeier et al. (eds.), Hotspots revisited: Earth's biologically richest and most endangered ecoregions, pp. 219-228, CEMEX, Mexico City. [ Links ]
United Nations Educational, Scientific and Cultural Organisation, 2015, Operational Guidelines for the Implementation of the World Heritage Convention, Intergovernmental Committee for the Protection of the World Cultural and Natural Heritage, United Nations Educational, Scientific and Cultural Organisation World Heritage Centre, Paris, viewed n.d., from http://whc.unesco.org/en/guidelines/ [ Links ]
Vamosi, J.C., Knight, T.M., Steets, J.A., Mazer, S.J., Burd, M. & Ashman, T.-L., 2006, 'Pollination decays in biodiversity hotspots', PNAS 103(4), 956-961. http://dx.doi.org/10.1073/pnas.0507165103 [ Links ]
Van den Bossche, P., 2001, 'Some general aspects of the distribution and epidemiology of bovine trypanosomosis in southern Africa', International Journal for Parasitology 31, 592-598. http://dx.doi.org/10.1016/S0020-7519(01)00146-1 [ Links ]
Waage, J.K., 1981, 'How the zebra got its stripes - Biting flies as selective agents in the evolution of zebra coloration', Journal of the Entomological Society of southern Africa 44, 351-358. [ Links ]
Walker, B.H., Emslie, R.H., Owen-Smith, R.N. & Scholes, R.J., 1987, 'To cull or not to cull: Lessons from a southern African drought', Journal of Applied Ecology 24, 381-401. http://dx.doi.org/10.2307/2403882 [ Links ]
 Correspondence:
Correspondence:
Adrian Armstrong
adrian.armstrong@kznwildlife.com
Received: 08 Aug. 2016
Accepted: 20 Oct. 2016
Published: 30 Jan. 2017


{kind=link}
{kind=link}
{kind=link}
{kind=link}