










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkKoedoe
versión On-line ISSN 2071-0771
versión impresa ISSN 0075-6458
Koedoe vol.56 no.1 Pretoria ene. 2014
http://dx.doi.org/10.4102/koedoe.v56i1.1231
SHORT COMMUNICATION
Impact of elephant on two woody trees, Boscia oleoides and Pappea capensis, in an arid thicket-Nama Karoo mosaic, Greater Addo Elephant National Park
Marietjie LandmanI; Angela GaylardII; Thando MendelaII; Graham I.H. KerleyI
IDepartment of Zoology, Nelson Mandela Metropolitan University, South Africa
IISouth African National Parks Scientific Services, Garden Route Regional Office, Knysna, South Africa
ABSTRACT
Despite extensive evidence of the influences of elephant on woody trees in savannah habitats, effects on trees in the succulent thickets of the Eastern Cape are relatively poorly described. Our study investigates the role and intensity of elephant impacts on Pappea capensis and the relatively rare Boscia oleoides in an arid thicket-Nama Karoo mosaic habitat of the Greater Addo Elephant National Park. We show that roughly 19% of the B. oleoides and nearly half of the P. capensisindividuals recorded showed signs of elephant impact. Elephant often toppled our study trees, and where these individuals were uprooted, mortalities occurred: B. oleoides ∼ 44% of the impacted trees (4 individuals); P. capensis ∼ 22% of the impacted trees (29 individuals).
CONSERVATION IMPLICATIONS: Whilst this study is restricted by limited spatial and temporal replication, P. capensis mortalities caused by elephant occurred at a rate exceeding that of other processes. Our results provide insight into the severity of the measured changes and the need to reduce the impacts. However, it would be critically important to establish the specific driver of elephant-tree interactions before any management intervention is implemented
Introduction
Understanding the distribution of elephant effects amongst elements of biological diversity is important for the conservation management of both elephant and biodiversity (Landman et al. in press). In particular, such an understanding provides key insights into the scale of monitoring needed to avoid mismatch at the management scale (Lindenmayer & Likens 2009). In savannah habitats, the effects of elephant on woody trees are well described; elephant (and fire) are considered to be the primary drivers of tree mortalities, changing habitat composition and structure (Kerley et al. 2008; O'Connor, Goodman & Clegg 2007). In the succulent thickets of the Eastern Cape, however, impacts on trees are relatively poorly described (notable exceptions include effects on Euclea undulata and Schotia afra [Landman et al.in press; Stuart-Hill 1992]). This is probably because research has focused almost exclusively on coastal thicket types (see the most recent review by Kerley & Landman2006), with a relatively poorly developed tree component. For these thicket types, elephant appear to have a disproportionate impact on rare and endemic low-growing succulents and geophytes, potentially causing local extinctions (Lombard et al. 2001). However, shifts in the relative importance of tree species (between thicket types) may also mean shifting impacts and increased vulnerability of trees.
The Kuzuko section of the Greater Addo Elephant National Park comprises a mosaic of arid thicket and Nama Karoo types, with a relatively well-developed woody tree component. Following an absence of nearly 100 years (Boshoff et al. 2002), elephant were re-introduced into the park during 2005 and have since had substantial impacts on this component of the flora: Pappea capensis and the relatively rare Boscia oleoides appear to be particularly vulnerable, suffering extensive toppling effects (Figure 1). This raises significant concerns regarding their mid- to long-term longevity, particularly as both species are exceptionally long-lived (some experts estimate up to ∼ 600 years) and form a key structural element of the landscape (Lechmere-Oertel, Kerley & Cowling 2005). This article describes the role and intensity of elephant effects on these woody trees in the study area.

Methods
Study area
We conducted the study in the Kuzuko section (covering ∼ 110 km2) of the Greater Addo Elephant National Park, located north of the Zuurberg Mountains in the Eastern Cape, South Africa. Sixteen elephant were re-introduced into the area in 2005, with the population increasing to twenty-one individuals during 2011. The region is semi-arid, with 200 mm-300 mm rainfall annually, often experiencing extended droughts. More than half of the study area comprises a series of low, undulating hills dominated by a mosaic of arid thicket and Nama Karoo types, whilst the steeper slopes of the Zuurberg mostly give way to arid thicket with a varying abundance of the tree-succulent Portulacaria afra (Boshoff et al. 2002).
Field survey
During January 2011 we located 17 transects across the arid thicket-Nama Karoo habitats of the study area. Transects were 10 m wide and 1 km long. Along each transect we recorded all B. oleoides and P. capensis individuals, distinguishing between individuals that (1) showed no impacts (No impact), (2) were impacted by elephant (Elephant; easily differentiated given the severity of the impacts) or (3) were affected by other or unknown factors (Other/Unknown). For all the affected trees (i.e. 2 and 3 above), we also determined the intensity of the impact according to three categories: (1) Broken branches only, (2) Toppled, but alive and (3) Dead.
Results
In total, we recorded 47 B. oleoides (0-10 indiv.transect−1) and 291 P. capensis (6-26 indiv.transect−1) trees (all mature) along transects. Roughly 19% of the B. oleoides and nearly half of the P. capensis individuals recorded showed signs of elephant impact (Figure 2). Whilst impacts in the Other/Unknown category appeared extensive, exceeding that caused by elephant in the case of B. oleoides, these were generally less severe (i.e. broken branches accounted for most of the impacts;Figure 3). Importantly, the impacts in this category never resulted in tree mortalities.

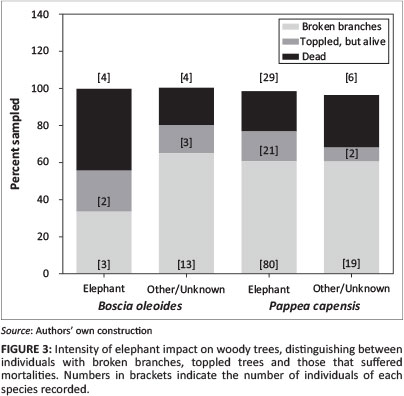
Elephant often toppled our study trees (Figure 3), and where these individual trees were uprooted, mortalities occurred: B. oleoides ∼ 44% of the impacted trees (4 individuals); P. capensis ∼ 22% of the impacted trees (29 individuals). It is not known which of the toppled trees that were alive at the time of sampling later died.
Because of their size, elephant are the primary drivers of tree toppling in our study area. Thus, it is very likely that the remaining toppled and dead individuals recorded (see the Other/Unknown category in Figure 2 and Figure 3) reflect background mortalities; in other words, mortalities related to tree demographics or other abiotic factors (e.g. lightning). For P. capensis, mortalities caused by elephant were nearly five times that caused by other processes (i.e. from 6 to 29 individuals). Our small B. oleoides sample population precluded any robust conclusions in this regard.
Discussion
Our results show the extensive impacts of elephant, particularly on P. capensis, in our study area. Interestingly, and despite numerous descriptions of the influences of elephant on savannah trees (e.g. Kerley et al. 2008; O'Connor, Goodman & Clegg 2007) and the wide geographic distribution of our study species, we provide the first record of elephant effects on these species. Thus, our study contributes towards understanding the distribution of elephant effects between plant species in succulent thickets and elsewhere.
However, our study is restricted by limited spatial and temporal replication. That is, transects were located in a single habitat, on relatively even terrain and with abundant water provisioning, which probably elevated the observed intensity of the impact (e.g. Kerley et al. 2008; Landman et al. in press). The steeper slopes of the Zuurberg Mountains may be inaccessible to elephant, providing a refuge habitat. Furthermore, we only measured the impacts once and consequently could not determine rates of toppling or mortality. Nevertheless, for P. capensis at least, mortalities caused by elephant occurred at a rate exceeding that of other processes.
It is important to recognise that whilst elephant were historically abundant on the coastal lowlands and along the river valleys of the Eastern Cape, the inland interfluves - such as Kuzuko - were only utilised ephemerally (Boshoff et al. 2002). This suggests that the woody trees in the region probably escaped intensive utilisation, both spatially and temporally. Also, most thicket plants showed limited recruitment from seeds (Lechmere-Oertel, Kerley & Cowling 2005), which could mean that the vulnerable, slow-growing and long-lived mature trees may not be readily replaced following these impacts. This provides insight into the severity of the measured changes and the need to reduce the impacts.
Management interventions that may be implemented to reduce the impacts should be based on robust cause-and-effect relationships between elephant and the impacts (Landman, Kerley & Schoeman 2008). Without this, conservation management interventions may fail in their objectives. As an example, the culling of elephant in the Kruger National Park did not prevent the loss of tall trees (Owen-Smith et al. 2006). Interestingly, tree toppling in our study area was often not associated with other foraging impacts (e.g. extensive branch breaking; M. Landman pers. obs., n.d.). Thus, reducing elephant numbers per se may be a costly intervention if trees are toppled during the strength training of mature bulls (Midgley, Balfour & Kerley 2005). Thus, it would be critically important to establish the specific driver of elephant-tree interactions in Kuzuko before any management interventions are implemented.
Recommendations for monitoring
Our study only provides a first approximation of the impact of elephant on two key species in the study area. It is important to expand this work to be representative of the habitat and topographic heterogeneity of the area, including information on the age structure and levels of recruitment of trees. Furthermore, there currently exists no information on the consequences of the impacts for other components of biodiversity. From Figure 1 it is clear that the toppling effects cause disturbance to soil processes and probably ecological functioning. In addition, these effects may (at least initially) facilitate access to forage for co-occurring browsing herbivores. Clearly, these knock-on effects require further research.
Acknowledgments
South African National Parks provided financial support to conduct the study. Izak Smit provided comments that improved the manuscript.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
M.L. (Nelson Mandela Metropolitan University) and G.I.H.K. (Nelson Mandela Metropolitan University) analysed the data and wrote the article. A.G. (Garden Route Regional Office) and T.M. (Garden Route Regional Office) designed the experiment and conducted the field survey.
References
Boshoff, A.F., Kerley, G.I.H., Cowling, R.M. & Wilson, S.L., 2002, 'The potential distributions, and estimated spatial requirements and population sizes, of the medium to large-sized mammals in the planning domain of the Greater Addo Elephant National Park project', Koedoe 45, 85-116. http://dx.doi.org/10.4102/koedoe.v45i2.19 [ Links ]
Kerley, G.I.H. & Landman, M., 2006, 'The impacts of elephants on biodiversity in the Eastern Cape Subtropical Thickets', South African Journal of Science 102, 395-402. [ Links ]
Kerley, G.I.H., Landman, M., Kruger, L., Owen-Smith, N., Balfour, D., De Boer, W.F. et al., 2008, 'Effects of elephant on ecosystems and biodiversity', in R.J. Scholes & K.G. Mennell (eds.), Elephant management: A scientific assessment for South Africa, pp. 146-204, Wits University Press, Johannesburg. [ Links ]
Landman, M., Kerley, G.I.H. & Schoeman, D.S., 2008, 'Relevance of elephant herbivory as a threat to Important Plants in the Addo Elephant National Park, South Africa',Journal of Zoology 274, 51-58. [ Links ]
Landman, M., Schoeman, D.S., Hall-Martin, A.J. & Kerley, G.I.H., in press, 'Long-term monitoring reveals differing impacts of elephant on elements of a canopy shrub community', Ecological Applications. [ Links ]
Lechmere-Oertel, R.G., Kerley, G.I.H. & Cowling, R.M., 2005, 'Patterns and implications of transformation in semi-arid succulent thicket, South Africa', Journal of Arid Environments 62, 459-474. http://dx.doi.org/10.1016/j.jaridenv.2004.11.016 [ Links ]
Lindenmayer, D.B. & Likens, G.E., 2009, 'Adaptive monitoring: A new paradigm for long-term research and monitoring', Trends in Ecology and Evolution 24, 482-486. http://dx.doi.org/10.1016/j.tree.2009.03.005 [ Links ]
Lombard, A.T., Johnson, C.F., Cowling, R.M. & Pressey, R.L., 2001, 'Protecting plants from elephants: Botanical reserve scenarios within the Addo Elephant National Park, South Africa', Biological Conservation 102, 191-203. http://dx.doi.org/10.1016/S0006-3207(01)00056-8 [ Links ]
Midgley, J.J., Balfour, D. & Kerley, G.I.H., 2005, 'Why do elephants damage savanna trees?', South African Journal of Science 101, 213-215. [ Links ]
O'Connor, T.G., Goodman, P.S. & Clegg, B., 2007, 'A functional hypothesis of the threat of local extirpation of woody plant species by elephant in Africa', Biological Conservation 136, 329-345. http://dx.doi.org/10.1016/j.biocon.2006.12.014 [ Links ]
Owen-Smith, N., Kerley, G.I.H., Page, G., Slotow, R. & Van Aarde, R.J., 2006, 'A scientific perspective on the management of elephants in the Kruger National Park and elsewhere', South African Journal of Science 102, 389-394. [ Links ]
Stuart-Hill, G.C., 1992, 'Effects of elephants and goats on the Kaffrarian Succulent Thicket of the Eastern Cape, South Africa', Journal of Applied Ecology 29, 699-710. http://dx.doi.org/10.2307/2404479 [ Links ]
 Correspondence:
Correspondence:
Marietjie Landman
PO Box 77000
Port Elizabeth 6031
South Africa
Email: marietjie.landman@nmmu.ac.za
Received: 28 Mar. 2014
Accepted: 31 Aug. 2014
Published: 17 Nov. 2014

