










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkKoedoe
versión On-line ISSN 2071-0771
versión impresa ISSN 0075-6458
Koedoe vol.55 no.1 Pretoria ene. 2013
ORIGINAL RESEARCH
Monitoring of the eggs of the Karkloof blue butterfly, Orachrysops ariadne, for its conservation management
Adrian J. ArmstrongI; Sharon L. LouwII
IBiodiversity Research & Assessment Division, Ezemvelo KwaZulu-Natal Wildlife, South Africa
IIEcological Advice, Ezemvelo KwaZulu-Natal Wildlife, South Africa
ABSTRACT
The Endangered Orachrysops ariadne (Butler 1898) (Karkloof blue butterfly) is endemic to the Endangered Moist Midlands Grassland in KwaZulu-Natal, South Africa, and is extant at four sites. The results from the monitoring of the eggs laid by O. ariadne in a grassland area that is frequently burnt by poor rural people to ensure that palatable grass is available to their livestock, suggested the implementation of management interventions (fencing and firebreak burning) to prevent the local extinction of the butterfly. The number of eggs at the monitoring site declined dramatically between 2002 and 2003 and fluctuated after the management interventions were initiated properly in 2008, but had nearly reached the target number of 250 by 2013. An index count method for the monitoring of O. ariadne eggs at the other three known colonies, where plant invasion rather than uncontrolled burning is a major threat, was developed and shown to be efficient with regard to time relative to the number of eggs sampled. The host ant Camponotus natalensis (F. Smith 1858) (Natal sugar ant) was found to be present in all the host-plant patches at one colony site, indicating that all host-plant patches are likely to be breeding areas for the butterfly. Invasive plant control at and appropriate burning of the habitat of O. ariadne should assist in ensuring the survival of these colonies.
CONSERVATION IMPLICATIONS: Adaptive monitoring and management of threatened endemic invertebrates and their habitats may be crucial for their continued survival. The development of efficient methods for the monitoring of such species is required where resources are limited, as threats to the species may cause sudden and irreversible declines in population size.
Introduction
Endangered species should not lose any more of their habitat if they are to survive in the wild in the long term (Pfab, Victor & Armstrong 2011). Monitoring of populations of threatened lepidopteran species that have obligatory relationships with host plants or ants will almost always be required for their effective conservation, especially where land transformation or habitat degradation is proceeding apace (Jewitt 2012; Murphy & Weiss 1988). Adaptive monitoring that enables adaptive management interventions to be initiated as and when appropriate, and that informs managers of the nature of the threats to the species and potential counters to those threats, would be very useful (Campbell et al. 2002; Greenwood & Robinson 2006; Lindenmayer & Likens 2009; Lindenmayer et al. 2011). Monitoring can be costly and therefore should be initiated only if clear and tangible benefits for the conservation of the species will result (Field et al. 2007).
The threats to a threatened species and knowledge of the biology of the species should guide the development of a monitoring programme and implementation of management actions for that species (Campbell et al. 2002), although certain threats may be detected only once a monitoring programme is underway. Monitoring of lepidoteran species needs to be long term, as responses of threatened butterflies to management interventions may not be immediate and may depend on other interacting factors such as weather conditions (Murphy & Weiss 1988; Smee et al. 2011; Thomas 2005). Long-term monitoring of threatened butterfly species is well established on other continents (e.g. Swengel et al. 2011; Thomas 2005), but is only in its infancy in Africa (for South Africa, see Edge et al. 2013; McGeoch et al. 2011).
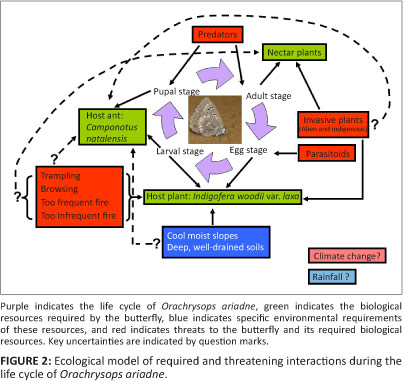
The Endangered Orachrysops ariadne (Butler 1898) (Karkloof blue butterfly) is known from only four colonies, all of which are in the highly transformed and Endangered Midlands Mistbelt Grassland in KwaZulu-Natal, South Africa (Figure 1; Armstrong et al. 1998; Armstrong & Edge 2013; Henning, Terblanche & Ball 2009; Lu & Samways 2001, 2002a, 2002b; Mucina & Rutherford 2006). None of the colonies has protected area status yet. The butterfly has an obligate host relationship with Indigofera woodii Bolus var. laxa Bolus (lax variety of Wood's indigo plant) and Camponotus natalensis (F. Smith 1858) (Natal sugar ant), laying eggs only on the host plant and apparently only where the host ant is present (Lu & Samways 2001). (See Pierce et al. [2002] for further discussion of ant-dependent oviposition, but cf. Edge, Cilliers & Terblanche [2008] for the related Orachrysops niobe [Trimen 1862] [Brenton blue butterfly]). The larvae feed only on the host plant and the adults feed on a limited number of nectar-plant species that flower during their flight period (Lu & Samways 2001). Any conservation action implemented for the butterfly must therefore conserve the butterfly's host-plant, host-ant and nectar-plant relationships and other required habitat conditions for it to be successful (Swengel et al. 2011; Tudor et al. 2004). The work by Lu and Samways (2001, 2002a, 2002b) elucidated some of the life history and behavioural ecology of the butterfly and enabled the formulation of conservation management recommendations. A conceptual ecological model of required and threatening interactions during the life cycle of O. ariadne is presented in Figure 2.
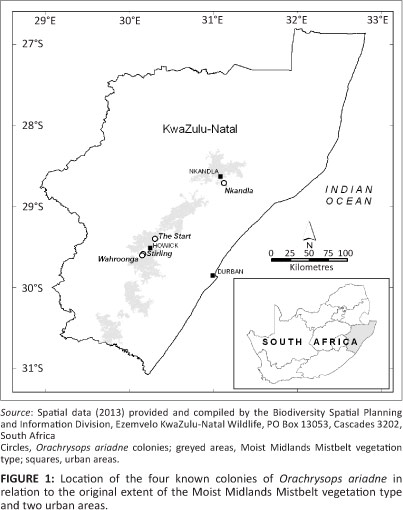
At least one known colony of O. ariadne, near Balgowan, became extinct last century and from verbal reports and the literature it seems probable that other colonies have also been extirpated through land transformation or habitat degradation (Armstrong 2010; Henning et al. 2009; Pennington 1946; Pringle, Henning & Ball 1994; Samways & Lu 2007). Mixed-species alien plant infestation was observed at the extinct colony site by the first author. Potential threats and other factors relating to the four extant colonies are presented in Table 1. All current landowners or land occupiers are very supportive of the monitoring of the colonies and management of the colony sites for the conservation of O. ariadne.
Monitoring of the O. ariadne colonies would be an important component of the management programme for this species, because it enables the current threats to the colonies to be determined, the response of the populations to management actions to be ascertained and timely corrective action to be initiated and maintained (Campbell et al. 2002; Greenwood & Robinson 2006). Monitoring of the eggs is a simple and efficient way of determining the trend in a population of O. ariadne, as well as ascertaining the factors that are causing a decline in the population (Lu & Samways 2001). The eggs are clearly visible on the host plant as they are deposited near the ends of growth tips of the mature host plant, which are not covered by other vegetation, and the eggs whiten with age from their initial green colour (Lu & Samways 2001; pers. obs.). Monitoring of non-adult life cycle stages can be, and has been, used to detect population size changes in some butterfly species (e.g. Hula et al. 2004; Murphy & Weiss 1988; Smee et al. 2011). Monitoring methods that are inexpensive and easy to implement by the managers or inhabitants of the land on which the colonies occur would be very useful(McGeoch et al. 2011).
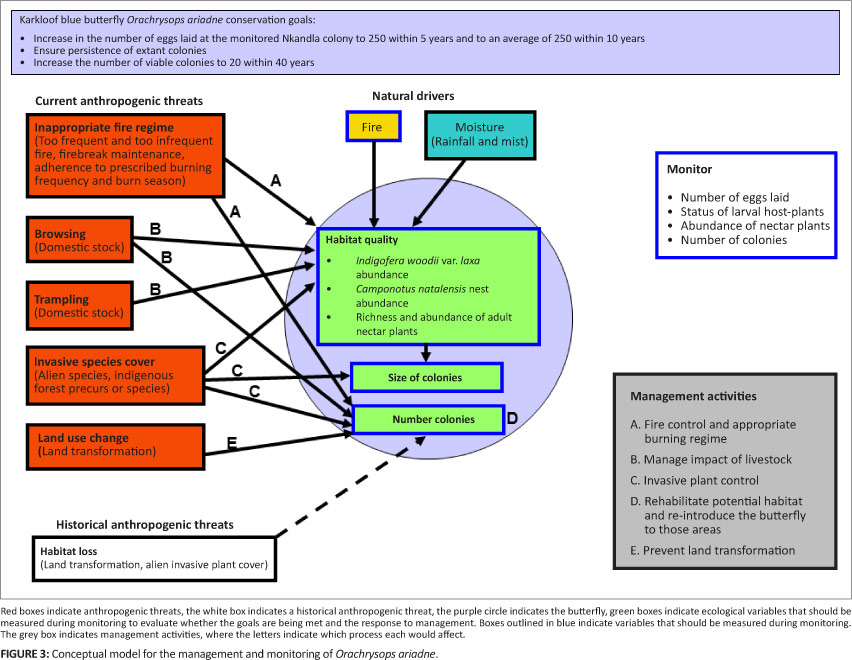
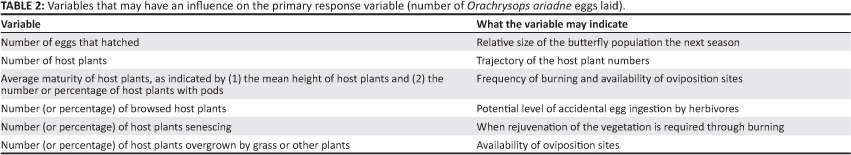
A conceptual model for the management and monitoring of O. ariadne is presented in Figure 3. The management objectives for O. ariadne will be achieved through various means, including adaptive management of the fire regime, fire control, livestock exclusion, invasive plant control, and through restoration of habitat for and re-introductions of O. ariadne. The key performance indicator for success of the monitoring and management programme is a sustained increase in the number of eggs laid (the primary response variable) until a set target number is reached, and thereafter the maintenance of egg numbers near this target indefinitely (Figure 3). Some variables that may have an influence on the value of the primary response variable are presented in Table 2. These variables may be measured during monitoring.
Only a short monitoring period for O. ariadne is consistently available each year as the species has a short flight period and embryonic development period (Lu & Samways 2001). Few personnel will be available to undertake the monitoring. These constraints would have to be taken into account when developing the monitoring programme. The objectives of the present study were to:
- determine whether management of threats to the Nkandla O. ariadne colony that caused its rapid decline, as ascertained through the long-term monitoring of egg numbers at the colony site, would change the trajectory of egg numbers towards a target of at least the number of eggs recorded during the first monitoring occasion
- develop and test a cost-effective monitoring index method for the other O. ariadne colonies to ensure that major declines in population size can be detected in time to prevent possible extirpation of the colonies.
Research methods and design
Monitoring area at the Orachrysops ariadne colony near Nkandla
A small valley, with an intermittent stream, was chosen as the monitoring area (0.24 ha) for the Nkandla colony, the breeding areas of which are discrete patches distributed in nearly 4 ha of grassland. This monitoring site was chosen because of its accessibility and the large number of host plants present and because the number of eggs present on the host plants could be counted during a day by a team of at least six staff. The latter requirement refers to the amount of time and number of personnel that are available to do the monitoring. Host plants growing in the several other small valleys inhabited by the butterfly were not included in the monitoring owing to resource constraints and to avoid excessive disturbance of the breeding habitat (Samways, McGeoch & New 2010). The boundary of the monitoring area was taken to be the edge of the valley (i.e. the top of the small ridge or watershed on both sides) and where the host plant distribution ended below the cliff at the top and above the road bank at the bottom.
The adult O. ariadne are on the wing from early March to late April, with male butterflies eclosing before the female insects. The maturation period for the developing larvae ranges from about 18-30 days in the field (Lu & Samways 2001). Monitoring was carried out 1 month after the last eggs were likely to have been laid in order to minimise the number of eggs that might not have hatched yet and also the number of eggshells that might be dislodged by frost or other factors. The monitoring commenced in 2002. All host plants considered large enough to be suitable for oviposition were searched for eggs; that is, single-stemmed seedlings were excluded. The precise number of I. woodii var. laxa surveyed in 2002 was not recorded as this was not deemed necessary then. Avoidance of double counting was implemented through awareness, communication between the monitors and the vigilance of the recorder. The number of monitors varied over the years, from 3 in 2003 to 19 in 2009 (median and mode: 10 monitors; mean ± 1 SD = 9.5 ± 4.25 monitors; n = 12). There was no significant correlation between egg numbers and number of monitors (r = -0.172; n = 12).
Eggs were classified visually according to the system of Lu and Samways (2001), using the naked eye or a magnifying glass. An egg from which a larva has successfully hatched had a round, clean-edged exit hole. We classified eggs that showed no signs of damage or parasitism as unhatched, being either infertile or still unhatched (sometimes the head capsule of the unhatched larva could be distinguished). Eggs that had a minute round hole on the surface in addition to the micropyle were classified as parasitised and combined with the damaged class, and all other eggs were classified as damaged (e.g. those that had ragged openings or were misshapen).
I. woodii var. laxa were considered overgrown if covered by surrounding vegetation without the protrusion of at least two I. woodii var. laxa shoots on which the adult female butterflies could lay. I. woodii var. laxa were classified as senescing if yellowing leaves, leaf fall and withered stems were evident, and as browsed by mammals where nipped shoots (shoots with obvious obliquely truncated ends) were evident. Plant heights were estimated to the nearest 10 cm and the presence of seed pods was recorded.
The management interventions at the Nkandla monitoring site included the burning of a firebreak around the site in 2006 and most years thereafter, and the fencing of the site in 2008. Alien plant control occurred once (2006) during the monitoring period. Since the approach taken is not experimental, the success of management interventions can only be inferred from the data until further factors that could potentially have caused the observed changes have been measured and a long-term dataset is obtained.
Development of a monitoring method for the three Orachrysops ariadne colony sites near Howick
The monitoring of adult O. ariadne using a mark-recapture method was shown not to be feasible (Samways & Lu 2007) because of the cryptic habits of the female butterflies. A total count of eggs laid at the three more southern colonies is impractical because their breeding areas are much greater than at the monitoring area of the Nkandla colony, a more limited number of monitors will be available, and the daily period of daylight suitable for monitoring eggs is less. Therefore, a subsample of the total egg number at a colony would have to be monitored. The boundary of the host plant distribution at each colony would have to be mapped and the distribution of the host ant be determined in relation to that of the host plant to prevent potential bias in where sampling takes place. Both hosts are essential to the completion of the butterfly's life cycle and the mobility of early instar O. ariadne larvae is likely to be limited. As a result, the eggs may be patchily distributed. These aspects of the study were investigated using the following methods.
Mapping the host plant distribution boundaries at the three colony sites near Howick
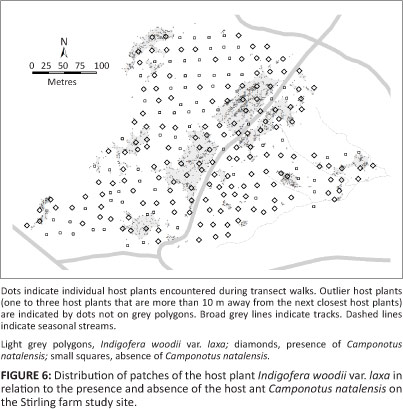
The boundary of the I. woodii var. laxa distribution was determined at each of the three colonies near Howick by searching for the host plants at the edge of suitable habitat and recording the positions of the edge host plants using a GPS receiver. The boundary was mapped by connecting the outer host plant positions that were at most 10 m apart. Single-stemmed seedlings were again ignored, as there was no certainty as to which would survive to maturity. The potential area of the O. ariadne breeding area could then be calculated. At Stirling, I. woodii var. laxa were surveyed by walking transects and recording the positions of host plants encountered between April 2007 and January 2008. Two to four observers walked approximately 2 m - 3 m abreast along transects, recording I. woodii var. laxa locations on and to about a metre on either side of the transects. These transects included straight transects through the habitat and zigzag transects along the apparent boundaries of the host plant patches to delimit the boundaries more accurately. The aim was not to record every single I. woodii var. laxa, but rather to define the limits of the various patches of host plants on the site. 'Outlier' host plants were those more than 10 m away from the next nearest host plants, including groups of up to three host plants that were more than 10 m away from the next nearest host plant. From visual observations, the distribution of the I. woodii var. laxa appeared to be closely correlated to the distribution of the Miscanthus capensis (Nees) Andersson (daba grass) on the site. Therefore areas of this grass were targeted for searching during the survey. Other areas were also searched, and from the data it became apparent that the distributions of I. woodii var. laxa and M. capensis were closely related, although the host plant was not as widely distributed as the grass species.
Pilot study at The Start Orachrysops ariadne colony
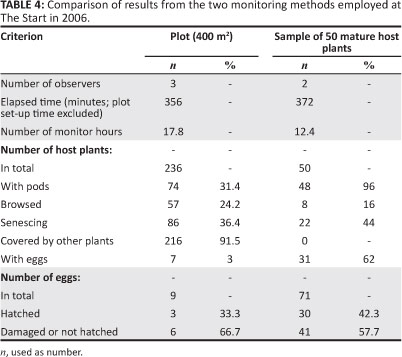
Both a plot method and an index method were compared for efficiency. The plot method might permit an estimate of the total number of eggs laid at the colony as the areas of both the plot and of the host plant distribution were known. It would also give an indication of whether a full egg count at the site was feasible given the constraints.
A plot of 20 m χ 20 m was placed roughly in the middle of The Start study area (approximately 1090 m.a.s.l.), where the setup was facilitated by the reasonably even slope. The plot was divided into four smaller quadrants (10 m χ 10 m each), identified as quadrants A (south-east quadrant), B (south-west quadrant), C (north-west quadrant) and D (north-east quadrant). This enabled accurate searching for host plants and detection of variation in host plant densities and egg densities over the whole plot. Each quadrant was systematically searched for I. woodii var. laxa and O. ariadne eggs by three people walking roughly parallel along the contours of the hillside and recording the number of host plants and eggs. Only plants rooted inside the plot were counted. The following features of the I. woodii var. laxa were recorded: presence of seed pods, signs of browsing by mammals and signs of senescence. Whether the surrounding vegetation completely covered the host plant was also recorded, as were the number of flowering indigenous encroacher Phymaspermum acerosum (DC.) Kallersjö (Currys Post plant) and the number of alien invader Rubus cuneifolius Pursh (American bramble) plants. The start and finish times were recorded for each quadrant. Since 0.038 ha of the colony's breeding area fell within the plot, the densities of host plants and eggs in this area were used to estimate the number of host plants and eggs in the whole breeding area of the colony, respectively. The densities of the invader or increaser plant species were calculated from the numbers of each species recorded in the entire 0.04-ha plot.
An index monitoring method might be very efficient in that specific host plants could be targeted to increase the number of eggs recorded per unit effort. When developing an index monitoring method it will have to be considered that:
- the number of host plants suitable for oviposition will vary from year to year according to the post-burn age of the vegetation
- the density of host plants will vary over the colony area
- the spatial distribution of the host ant is not likely to change in response to fire as the ants live underground and therefore should not be affected directly by fire
- as a result of these and perhaps other factors, the distribution of O. ariadne eggs is likely to change somewhat each year (Figure 4).
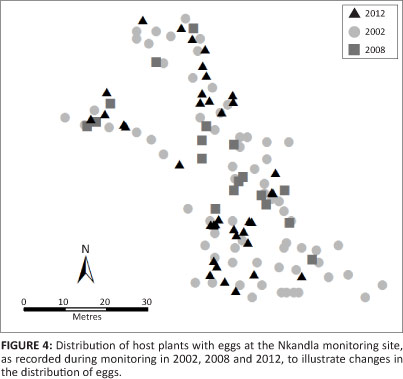
Therefore, the index monitoring method will have to include some randomness in its spatial component and the number of host plants that can be searched for eggs in a day will be one of the necessary considerations. During the pilot phase of the study, two observers walked through the colony's breeding area and searched 50 large (mature) host plants suitable for oviposition as an indexing method, as they were encountered randomly in various parts of the site outside the plot. The data recorded were similar to those during the plot count (excluding the number of encroacher and invader plants), but also included a visual estimate (to the nearest 10 cm) of the relative height of each I. woodii var. laxa, as assessed against an observer's height.
Camponotus natalensis distribution in relation to the distribution of Indigofera woodii var. laxa
The distribution of the host ant, C. natalensis, in relation to the host plant was investigated at Stirling farm. Sampling for the host ant was initiated in February 2008 and was completed by mid-March 2008. Weather conditions were variable during the survey period and nights were frequently misty and wet. Sampling was abandoned in rain. A grid of points 20 m apart was laid out over the site encompassing the area where I. woodii var. laxa grew. Each point was used as a baiting station and the actual coordinates on the ground were captured using a hand-held GPS receiver. Each baiting station was a Perspex square of 10 cm χ 10 cm placed on cleared ground of approximately 30 cm in diameter. Bait was applied in the late afternoon and left for 90 min before surveying. The bait consisted of a honey-and-brown sugar mix, diluted with water to give a watery consistency, and a small amount of Bovril added to provide protein. The baiting stations were checked for ant activity after dark. Each baiting site (including the baiting station, the bare ground surrounding the station and the peripheral vegetation, up to the top of the grass layer) was searched very carefully for about 1 min and the C. natalensis were counted.
Implementation of the 'mature host plants index method' at The Start
The 'mature host plants index method' was refined by determining the number of mature host plants that should be sampled at The Start to ensure an appropriate egg sample size. An appropriate egg sample size was estimated using the formula (Watson 2001):
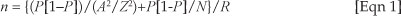
where n = sample size required, N = estimated number of eggs in the population, P = estimated variance in the population, A = the precision desired (± 5%), Z is the confidence level (95%, i.e. 1.96), and R is the estimated response rate (100%, i.e. 1.0).
An estimate of the number of reproductively mature host plants that should be searched to obtain the required sample size of eggs was then calculated from the data collected during the pilot study at The Start in 2006. Two observers searched for eggs on this number of mature host plants
(i.e. those with pods) at The Start on 16 and 17 May 2012. They used a hand-held GPS receiver to guide them along predetermined transects spaced 20 m apart and running north-south through the colony's breeding area. These transects were numbered consecutively from west to east. The order of transects to be walked until the calculated number of mature host plants had been searched for eggs was determined via a random number generator. Mature host plants observed within 2 m of the centre line of each transect walked were searched for eggs.
Statistical analyses and geographic information systems
All statistical analyses were performed using Statistica 10 (StatSoft, Inc., Tulsa, Oklahoma). Spatial data building was performed using CartaLinx (Clark Labs, Clark University, Worcester MA). The Idrisi 15 Andes Version (Clark Labs, Clark University, Worcester MA, USA) and ArcView 3.2 (Esri, Inc., Redlands CA) geographic information systems were used for geographical coverage analyses.
Results
Monitoring at the Nkandla colony
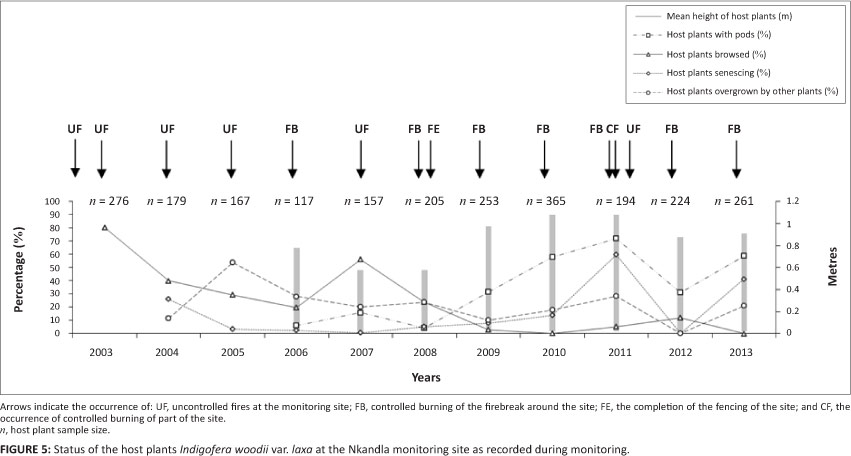
Browsing, fire and host plants
The relatively extensive browsing of the host plants was curtailed by the erection of a fence in 2008, after which no or little browsing was recorded (Figure 5). Uncontrolled fires burnt through the monitoring site each year between 2002 and 2005 (Figure 5). The establishment of the firebreak enabled the fires and their effects on the host plants to be controlled most years thereafter. Exceptions included the burning of the site by an uncontrolled fire in July 2007 after the egg monitoring took place that year, and in 2011, again after the egg count. During the latter fire, about half the site was burnt in August as part of the site management programme, followed later by an unauthorised fire that burnt the rest of the site.
A decline in the number of I. woodii var. laxa was recorded between 2004 and 2006, when fewer than half the number of host plants surveyed in 2003 were visible owing to the uncontrolled fires (Figure 5). A recovery in the numbers of I. woodii var. laxa surveyed occurred between 2007 and 2010, when the number exceeded the number in 2003, owing to the presence of the firebreak. Another decline in visible host plant numbers was recorded in 2011 due to the fires that year, after which the number of visible host plants increased. The percentage of host plants with pods was low from 2006 to 2008, but increased markedly from 2009 to 2011 owing to the protective effect of the firebreak, which allowed the host plants and their re-growth to mature. The rejuvenating effect of fire is seen in the decline in the percentage of host plants showing signs of senescence between 2004 and 2007, and the subsequent increase from 2010 to 2011 in the absence of fire. Signs of senescence declined to near zero and the percentage of host plants with pods declined in 2012 after the site was burned in 2011.
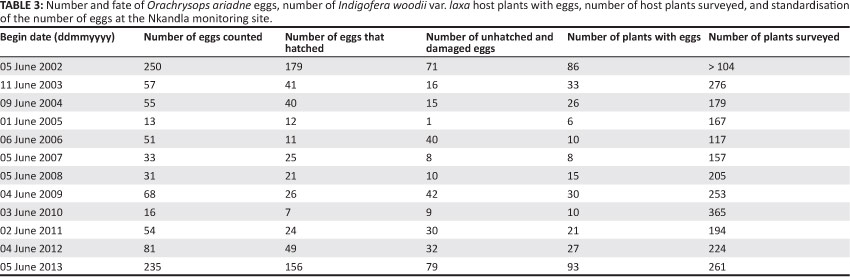
The mean height of the host plants varied according to the frequency of uncontrolled fires, decreasing after each burn and generally increasing over the years without fire, as seen between 2008 and 2011 (Figure 5). In 2005, after a series of annual burns and with extensive browsing, many of the host plants were overgrown by surrounding vegetation (Figure 5). The percentage of host plants overgrown by surrounding vegetation gradually increased from 2009 to 2011, after control over fire and browsing was established. This percentage dropped practically to zero in 2012 after the entire monitoring site was burned in 2011. The number of I. woodii var. laxa with O. ariadne eggs declined overall by 93% between 2002 and 2005, fluctuated somewhat thereafter until 2010, and then increased until 2013 (Table 3).
Egg numbers
The results of the O. ariadne egg monitoring at the Nkandla colony during the study period are presented in Table 3.
The number of eggs recorded decreased by 77% between 2002 and 2003, and by 95% between 2002 and 2005. After the management interventions were effectively implemented in 2008, the recorded number of eggs fluctuated and then increased year on year from 2011 to 2013, when the number of eggs recorded was nearly at target. No significant correlation was found between the number of eggs and the number of host plants between 2003 and 2013 (r = 0.167, n = 11,p > 0.05). The management of threats appears to have contributed to reversing the population trajectory of both O. ariadne and I. woodii var. laxa at the Nkandla monitoring site.
Pilot study to determine a monitoring method for the three colony sites near Howick Host-plant distribution boundaries
The host-plant distribution at The Start was 1.35 ha in extent, consisting of two patches divided by a narrow strip of riparian vegetation. This was the estimated extent of the O. ariadne breeding area here. The area of the host-plant patch at Wahroonga farm was calculated to be 8.15 ha. The boundaries of the host-plant patches (1.57 ha in total area) and the location of outlier I. woodii var. laxa at Stirling farm are indicated in Figure 6. Host plants were not uniformly distributed over the site, but were concentrated on southeast-facing slopes.
Comparison of the plot and the 'mature host plants sample' monitoring methods
The 50 mature host plants sample method was much more efficient than the plot method (Table 4). In the sample, a significantly higher proportion of host plants had pods (χ2 = 70.49; df= 1; p < 0.0001) and eggs (χ2 = 124.80; df= 1; p < 0.0001), and a significantly lower proportion were covered by other plants (χ2 = 186.97; df = 1;p < 0.0001) compared with the plot (the other comparisons of proportions were not significant). Even in the plot, which had host plants of a variety of ages, the majority of the host plants selected for oviposition by female O. ariadne were reproductively mature. The density of host plants and eggs in each plot quadrant varied between 31 and 106 and 0 and 7 per 100 m2, respectively. From the density of eggs and host plants recorded in the plot, an estimated 22% (71 out of 320) of the eggs at the colony were in the egg sample from the 50 mature host plants. A total of 38 P. acerosum plants, one Maesa lanceolata Forssk. (false assegai) shrub and one R. cuneifolius were recorded from the entire plot. From these figures, a total of 8384 I. woodii var. laxa, 320 eggs (107 hatched), 1283 P. acerosum plants, 34 M. lanceolata and 34 R. cuneifolius plants were estimated for the breeding area of the O. ariadne colony.
Mapping the host-ant distribution at the Stirling colony breeding area
The distribution of C. natalensis in relation to the I. woodii var. laxa distribution at Stirling farm is shown in Figure 6. The host ant was present in all the host-plant patches, but absent from some of the areas where host plants were absent. There were significantly more C. natalensis occurrences in the host-plant patches than expected and no absences of the host ant there (χ2 = 21.88; df = 1; p < 0.01). C. natalensis was recorded near many of the outlier host plants. From these results, only the distribution of the host plant needs to be taken into account in the monitoring method.
Implementation of the 'mature host plants index' method at The Start
Using equation (1) and the estimates from the pilot study, the appropriate egg sample size was estimated at 88 eggs, with N estimated at 320 eggs and P estimated at 0.4 (approximately 40% of the 50 mature host plants sampled when testing the index method did not have O. ariadne eggs, suggesting that the population could increase by a maximum of 40%). Searching of the 50 mature host plants during the pilot study returned 71 eggs. Therefore, to have recorded an adequate sample of 88 eggs using the index method during the pilot study, the estimated number of mature host plants that would have had to be searched was 88/71*50 = 61.97, (i.e. 62) mature host plants.
Following from this calculation, 62 reproductively mature host plants were searched for eggs at The Start colony in May 2012. Most of the 31 eggs observed had hatched; 29% did not hatch because they were damaged (6) or assumed parasitised (3). Three of the host plants with eggs showed signs of being browsed. The total area covered by the 547-m transect length walked was 2188 m2, representing 16.22% of the total colony breeding area.
Discussion
Monitoring and habitat management at the Nkandla colony
The Nkandla O. ariadne population was relatively large before and at the start of the monitoring programme (Lu & Samways 2001; this study). This population is a priority for conservation as it is separated from the other three known colonies by at least 120 km (Figure 1). The precarious situation of the monitored population of O. ariadne at the Nkandla site between 2003 and 2008 was the result of a lack of appropriate management of the colony area. The management guidelines for this colony included the necessity for maintaining a fire return frequency of 2-4 years (Lu & Samways 2002b) and for domestic livestock to be excluded from the breeding areas. The acceptance of, and involvement with, the management programme at Nkandla by local residents was a very important factor in eventually achieving appropriate management of the site and one that should assist in eventually achieving the target number of eggs and the overall success of the project (Louw & Armstrong 2010). Such cooperative management should become a feature of future invertebrate conservation programmes in Africa to increase the probability that the management goals will be achieved.
The large decline in the number of eggs at the monitored site between 2002 and 2003 was apparently partly caused by the browsing of the I. woodii var. laxa by domestic stock and perhaps also indigenous antelope. The eggs are usually laid within 3 cm of the tips of the buds of the I. woodii var. laxa and therefore the eggs are very vulnerable to accidental ingestion when the shoots are eaten by mammalian browsers (Lu & Samways 2002a). Fencing of the monitoring site was finally achieved in 2008 (Louw & Armstrong 2010), which prevents cattle and goats from accessing the host plants at the site. Cattle are known to have detrimental impacts on threatened butterfly host-plant abundance and habitat structure elsewhere (e.g. Schtickzelle, Turlure & Baguette 2007).
The decline in the number of visible I. woodii var. laxa through the agency of fire between the 2003 and 2004 monitoring events indicated the need for careful management of the habitat through fire. The firebreak around the Nkandla monitoring site has protected the monitoring site from uncontrolled fires since 2008, allowing appropriate fire management of the site. Fire is the best means of defoliation on the site as the vegetation and invertebrates such as O. ariadne are adapted to fire and many are dormant or underground during the fire season. The host plants are sensitive to fire before they are 2 years old (Lu & Samways 2001), but resprout as adults and also reproduce via seed. The vigour and health of the host plant community and the habitat as a whole are maintained by fire, including by enabling the recruitment of I. woodii var. laxa to offset the number that senesce.
Very few mature I. woodii var. laxa were visible in the monitoring area in 2005 owing to the prior uncontrolled fires (one in August 2004 and two in 2003). Most of the remaining host plants were young and many were covered by other vegetation. Female O. ariadne generally lay their eggs on the larger, mature I. woodii var. laxa and not on host plants covered by other vegetation (this study; Lu & Samways 2002a). Browsing of I. woodii var. laxa also continued. As a result, few eggs were recorded in 2005, 2006 and 2007. The exclusion of fire in 2008, 2009 and 2010 caused an increase in the percentage of host plants showing signs of senescence from 0.6% in 2007 to 59.8% in 2011. This indicates the need for rejuvenation of the host plant population by fire. The controlled fire in 2011 (and apparently also the unauthorised burn that year) achieved this purpose, and in 2013 the egg numbers (235) had nearly reached the target of 250, the number recorded in 2002. The trend in egg numbers is positive and a potential extirpation of the species at the site has been avoided. Although inappropriate burning of habitat affects some threatened butterfly species or subspecies (Powell, Busby & Kindscher 2007; Swengel et al. 2011), other threatened butterfly species require regular but not very frequent (and sometimes small-scale) burning of their habitats at the appropriate time of year and day to maintain habitat heterogeneity and the availability of required resources without killing vulnerable stages of the life cycles of the butterflies (see, e.g. New et al. 2010; Pickens & Root 2009).
The management interventions appear to have contributed to reversing the population trajectory of both O. ariadne and I. woodii var. laxa and need to be extended to the adjacent grassland areas, including the south-facing seasonal stream valleys where O. ariadne is known to occur. The areas that would require fencing are relatively small and the loss of browse for domestic stock would not be extensive. The total activity area for an O. ariadne colony is larger than the area in which the host plant is distributed, as the adult nectar plants may be more widely distributed than the host plant (Dennis, Shreeve & Van Dyck 2006; Samways & Lu 2007). The monitoring of nectar resources for butterflies of conservation concern, such as O. ariadne, is important because such butterflies can be flower-feeding specialists that have short flight periods, low mobility and biotope specialisation (Lu & Samways 2001; Samways & Lu 2007; Schultz & Dlugosch 1999; Tudor et al. 2004). Monitoring of nectar plants should be added to the present monitoring programme.
Monitoring and habitat management at the three colony sites near Howick
The results obtained using the index method at The Start suggested that sampling a certain number of mature I. woodii var. laxa for eggs is an efficient monitoring method with regard to time and resources and appears to be suitable for monitoring the status of other O. ariadne populations. Furthermore, it avoids the greater disturbance of the vegetation around the host plants associated with the plot counts. Congruence in the distribution of the host plant and the host ant, as determined at the Stirling colony site, indicates that transects can be placed either systematically or randomly over the breeding area. Assuming similar mean densities of host plants at all colony sites, and considering the ratio between the size of the breeding area at The Start and the breeding areas at the two other colonies near Howick, we recommend that at least 80 and 411 mature host plants are sampled for eggs on each monitoring occasion at Stirling and Wahroonga, respectively. At least four monitors would be required at Wahroonga to ensure that the sampling is finished within 2 days. The index method will have to be put into practice for some years and tested to see whether it is sensitive enough to detect negative unidirectional trends in population size rapidly enough to enable effective management interventions to be implemented to reverse the declines (Field et al. 2007). An increase in the robustness of estimates of the number of mature host plants to sample at each of the three colonies near Howick is desirable in the future. To this end, sampling a sufficient number of randomly placed plots or transects at each site would allow measures of variability within and between populations to be calculated. The results of the monitoring at the Nkandla colony suggest that monitoring should eventually be carried out at least every other year so that any major decline in population size is noticed quickly enough to effect a reversal.
The percentage of I. woodii var. laxa in the plot (24.2%) and in the index method sample (16%) at The Start that had signs of being browsed suggests that monitoring the eggs around the time of hatching of the last eggs is most appropriate as monitoring later may result in greater egg loss and monitoring earlier may under-estimate the number of eggs that hatched. Browsing by antelope could lead to an underestimation of the number of eggs laid and the host plants, being legumes, are suitable browse in the relatively dry and cold winter of in the mistbelt region of KwaZulu-Natal. Lu and Samways (2001) found that all the eggs monitored at The Start colony between 1997 and 1999 had hatched by 24 May. Monitoring of eggs at the colonies near Howick should therefore be carried out around this time of year. Rotational burning of about a third of the grassland at each site each year may be useful for promoting seedling growth and survival in the burnt area and in rejuvenating the foliage of mature host plants that resprout after fire. How fire might affect the C. natalensis is unknown, but the butterfly's habitat should be burnt during the day as the host ant is nocturnal and is underground during the day (Lu & Samways 2002a). Burning should also be performed only after the butterfly eggs have hatched and when the larvae are underground in ant nests.
The main threat to the habitat of O. ariadne at the three colonies near Howick appears to be encroachment by indigenous plants and invasion by alien plants into the breeding areas. P. acerosum, M. lanceolata and R. cuneifolius should be monitored as these species might out-compete the host plants and other plants for resources such as light, water, soil nutrients and space. The number of P. acerosum at The Start colony was estimated at several thousand, and although only one R. cuneifolius was recorded in the plot, many were found along the edges of the watercourses and stream. Edge et al. (2008) found that the encroacher Pteridium aquilinum (L.) Kuhn (bracken fern) regrows rapidly after fire and preoccupies space from the slower-growing Indigofera erecta Thunb. (erect indigo plant), the host plant of O. niobe, resulting in a low recruitment success for the host plant. A high biomass of the relatively large P. acerosum could lead to intense fires, which could kill the younger I. woodii var. laxa. A programme to actively control these species is in place, but the efficacy of the programme in reducing the number of invasive plants needs to be monitored.
Future priorities
Apart from monitoring the O. ariadne colonies, a priority is to continue the search for other colonies of these species and potential re-introduction sites, guided by the predicted distribution of the species (Armstrong 2010). If possible, grassland linkages of adequate width and quality (Pryke & Samways 2001, 2003) between colonies will need to be negotiated with landowners on the assumption that O. ariadne can disperse between colonies. Although individuals observed away from the known colony areas may lend support to the latter assumption (Pennington 1946), the assumption needs to be tested through genetic studies. The results of Samways and Lu (2007) indicate that male O. ariadne tend to remain in the colony area but that they are powerful fliers. The genetic distance between the Nkandla population and the populations near Howick needs to be established so that the sources of any re-introduced butterflies can be wisely chosen. The overall vision is to have thriving O. ariadne populations in various parts of the probable former distribution range of the species one day.
Conclusion
This study indicated that adaptive monitoring and management of threatened and endemic invertebrate species, with the cooperation of the owners, occupiers or managers of the land on which the species occur, may be crucial for the successful conservation of such species. Examples such as that of the Erikssonia edgei Gardiner & Terblanche 2010 (Waterberg copper butterfly) exist, where the local extinction of the species may have been prevented with the implementation of landowner-assisted adaptive monitoring and management (Henning et al. 2009). As land transformation continues in South Africa (e.g. Driver et al. 2012; Jewitt 2012) and elsewhere in Africa, such inclusive conservation efforts will be required for more invertebrate species. The need for more species-specific management protocols to ensure the long-term persistence of threatened habitat-specialist butterflies has been highlighted elsewhere (New et al. 2010; Swengel et al. 2011). In order to determine the most appropriate habitat management protocols for threatened South African grassland-specialist butterfly species, studies of the life history and ecology of each (e.g. see Lu & Samways [2001, 2002a, 2002b] for O. ariadne and Thomas [2005] for grassland species elsewhere), should be prioritised. The long-term conservation of these species will be more assured if adaptive management of their habitats in response to the information obtained through monitoring is effectively implemented, given the other requirements that may pertain for each species, such as the adequate spatial arrangement of sufficiently large colonies or populations within dispersal distance.
Acknowledgements
We gratefully thank the Midlands Meander Association, and especially Nikki Brighton, for their continued sponsorship and encouragement of our research into the conservation of the Karkloof blue butterfly, and especially for the bursaries that the Association awarded to Keenan Steers and Nonku Mkhize to assist them with their university studies and the fieldwork reported on here. We thank Hylton Adie for sampling ants and for his report, Dino Santoro for permission to work on Stirling farm, Rose Kunhardt for permission to work on Wahroonga farm, and John Craigie, Sihle Madonsela, Callum Beattie, George Pizzey, Liandra Bertolli and Caitlin Thorpe for assistance in the field. All the monitors are thanked for their participation in the study. Douglas Macfarlane and the Managers of The Start property from SAPPI are thanked for inviting and allowing us to undertake the work there. Several anonymous reviewers are thanked for their comments on previous versions of this manuscript.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
A.J.A. (Ezemvelo KwaZulu-Natal Wildlife) was the project leader, designed the study, carried out the fieldwork together with S.L.L. (Ezemvelo KwaZulu-Natal Wildlife), analysed the data and wrote the manuscript. S.L.L. organised and carried out the fieldwork at the Nkandla monitoring site and helped capture the data, liaised with the amaChube Traditional Authority, provided suggestions for the methodology, organised and guided the management of the Nkandla monitoring site and commented on drafts of the article.
References
Armstrong, A.J., Benn, G., Bowland, A.E., Goodman, P.S., Johnson, D.N., Maddock, A.H. et al., 1998, 'Plantation forestry in South Africa and its impact on biodiversity', South African Forestry Journal 182, 59-65. http://dx.doi.org/10.1080/10295925.1998.9631191 [ Links ]
Armstrong, A.J., 2010, 'Box 9.1 The use of cartographic modelling to map potential habitat for threatened endemic species', in M.J. Samways, M.A. McGeoch & T.R. New (eds.), Insect conservation: A handbook of approaches and methods, pp. 290-292, Oxford University Press, Oxford. PMid:19967358 [ Links ]
Armstrong, A.J. & Edge, D.A., 2013, 'Orachrysops ariadne (Butler, 1898a)', in S. Mecenero, J.B. Ball, D.A. Edge, M.L. Hamer, G.A. Henning, M. Kruger et al. (eds.), Conservation assessment of butterflies of South Africa, Lesotho and Swaziland: Red List and atlas, p. 542, Saftronics (Pty) Ltd., Johannesburg & Animal Demography Unit, Cape Town. [ Links ]
Campbell, S.P., Clark, J.A., Crampton, L.H., Guerry, A.D., Hatch, L.T., Hosseini, P.R. et al., 2002, 'An assessment of monitoring efforts in endangered species recoveryplans', Ecological Applications 12, 674-681. http://dx.doi.org/10.1890/1051-0761(2002)012[0674:AAOMEI]2.0.CO;2 [ Links ]
Dennis, R.L.H., Shreeve, T.G. & Van Dyck, H., 2006, 'Habitats and resources: The need for a resource-based definition to conserve butterflies', Biodiversity and Conservation 15, 1943-1966. http://dx.doi.org/10.1007/s10531-005-4314-3 [ Links ]
Driver, A., Sink, K.J., Nel, J.N., Holness, S., Van Niekerk, L., Daniels, F. et al., 2012, 'National Biodiversity Assessment 2011: An assessment of South Africa's biodiversity and ecosystems', synthesis report, South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria. [ Links ]
Edge, D.A., Cilliers, S.S. & Terblanche, R.F., 2008, 'Vegetation associated with the occurrence of the Brenton blue butterfly', South African Journal of Science 104, 505-510. http://dx.doi.org/10.1590/S0038-23532008000600028 [ Links ]
Edge, D.A., Woodhall, S.E., Ball, J.B., Henning, G.A., Armstrong, A.J. & Mecenero, S., 2013, 'Future priorities for butterfly conservation and research', in S. Mecenero, J.B. Ball, D.A. Edge, M.L. Hamer, G.A. Henning, M. Kruger et al. (eds.), Conservation assessment of butterflies of South Africa, Lesotho and Swaziland: Red List and atlas, pp. 35-40, Saftronics (Pty) Ltd., Johannesburg & Animal Demography Unit, Cape Town. [ Links ]
Field, S.A., O'Connor, P.J., Tyre, A.J. & Possingham, H.P., 2007, 'Making monitoring meaningful', Austral Ecology 32, 485-491. http://dx.doi.org/10.1111/j.1442-9993.2007.01715.x [ Links ]
Greenwood, J.J.D. & Robinson, R.A., 2006, 'Principles of sampling', in W.J. Sutherland (ed.), Ecological census techniques, pp. 11-86, Cambridge University Press, Cambridge. http://dx.doi.org/10.1017/CBO9780511790508.003 [ Links ]
Hamer, M., 2005, 'Report on the monitoring of Karkloof blue butterfly populations in 2005', Inland Invertebrate Initiative, University of KwaZulu-Natal, Pietermaritzburg. [ Links ]
Henning, G.A., Terblanche, R.F. & Ball, J.B. (eds.), 2009, South African red data book: Butterflies, SANBI Biodiversity Series 13, South African National Biodiversity Institute, Pretoria. [ Links ]
Hula, V., Konvicka, M., Pavlicko, A. & Fric, Z., 2004, 'Marsh fritillary (Euphydryas arinia) in the Czech Republic: Monitoring, metapopulation structure, and conservation of an endangered butterfly', Entomologica Fennica 15, 231-241. [ Links ]
Jewitt, D., 2012, 'Land cover change in KwaZulu-Natal', Environment 10, 12-13. [ Links ]
Lindenmayer, D.B. & Likens, G.E., 2009, 'Adaptive monitoring: A new paradigm for long-term research and monitoring', Trends in Ecology and Evolution 24, 482-486. http://dx.doi.org/10.1016/j.tree.2009.03.005, PMid:19409648 [ Links ]
Lindenmayer, D.B., Likens, G.E., Haywood, A. & Miezis, L., 2011, 'Adaptive monitoring in the real world: Proof of concept', Trends in Ecology and Evolution 26, 641-646. http://dx.doi.org/10.1016/j.tree.2011.08.002, PMid:21880394 [ Links ]
Louw, S. & Armstrong, A.J., 2010, 'Box 11.5 Community involvement in the conservation of threatened endemic invertebrate species', in M.J. Samways, M.A. McGeoch & T.R. New (eds.), Insect conservation: A handbook of approaches and methods, pp. 368-370, Oxford University Press, Oxford. [ Links ]
Lu, S-S. & Samways, M.J., 2001, 'Life history of the threatened Karkloof blue butterfly, Orachrysops ariadne (Lepidoptera: Lycaenidae)', African Entomology 9, 137-151. [ Links ]
Lu, S-S. & Samways, M.J., 2002a, 'Behavioural ecology of the Karkloof blue butterfly Orachrysops ariadne (Lepidoptera: Lycaenidae) relevant to its conservation', African Entomology 10, 137-147. [ Links ]
Lu, S-S. & Samways, M.J., 2002b, 'Conservation management recommendations for the Karkloof blue butterfly, Orachrysops ariadne (Lepidoptera: Lycaenidae)', African Entomology 10, 149-159. [ Links ]
McGeoch, M.A., Sithole, H., Samways, M.J., Simaika, J.P., Pryke, J.S., Picker, M. et al., 2011, 'Conservation and monitoring of invertebrates in terrestrial protected areas', Koedoe 53, Art. #1000, 13 pages. http://dx.doi.org/10.4102/koedoe.v53i2.1000 [ Links ]
Mucina, L. & Rutherford, M.C. (eds.), 2006, The vegetation of South Africa, Lesotho and Swaziland, Strelitzia 19, South African National Biodiversity Institute, Pretoria. [ Links ]
Murphy, D.D. & Weiss, S.B., 1988, 'A long-term monitoring plan for a threatened butterfly', Conservation Biology 2, 367-374. http://dx.doi.org/10.1111/j.1523-1739.1988.tb00201.x [ Links ]
New, T.R., Yen, A.L., Sands, D.P.A., Greenslade, P., Neville, P.J., York, A. et al., 2010, 'Planned fires and invertebrate conservation in south east Australia', Journal of Insect Conservation 14, 567-574. http://dx.doi.org/10.1007/s10841-010-9284-4 [ Links ]
Pennington, K.M., 1946, 'Notes on some rare South African butterflies', Journal of the Entomological Society of South Africa 9, 20-27. [ Links ]
Pfab, M.F., Victor, J.E. & Armstrong, A.J., 2011, 'Application of the IUCN Red Listing system to setting species targets for conservation planning purposes', Biodiversity and Conservation 20, 1001-1012. http://dx.doi.org/10.1007/s10531-011-0009-0 [ Links ]
Pickens, B.A. & Root, K.V., 2009, 'Behavior as a tool for assessing a managed landscape: A case study of the Karner blue butterfly', Landscape Ecology 24, 243-251. http://dx.doi.org/10.1007/s10980-008-9302-z [ Links ]
Pierce, N.E., Braby, M.F., Heath, A., Lohman, D.J., Mathew, J., Rand, D.B. et al., 2002, 'The ecology and evolution of ant association in the Lycaenidae (Lepidoptera)', Annual Review of Entomology 47, 733-771. http://dx.doi.org/10.1146/annurev.ento.47.091201.145257, PMid:11729090 [ Links ]
Powell, A.F.L.A., Busby, W.H. & Kindscher, K., 2007, 'Status of the regal fritillary (Speyeria idalia) and effects of fire management on its abundance in northeastern Kansas, USA', Journal of Insect Conservation 11, 299-308. http://dx.doi.org/10.1007/s10841-006-9045-6 [ Links ]
Pringle, E.L.L., Henning, G.A. & Ball, J.B. (eds.), 1994, Pennington's Butterflies of Southern Africa, Struik, Cape Town. [ Links ]
Pryke, S.R. & Samways, M.J., 2001, 'Width of grassland linkages for the conservation of butterflies in South African afforested areas', Biological Conservation 101, 8596. http://dx.doi.org/10.1016/S0006-3207(01)00042-8 [ Links ]
Pryke, S.R. & Samways, M.J., 2003, 'Quality of remnant indigenous grassland linkages for adult butterflies (Lepidoptera) in an afforested African landscape', Biodiversity and Conservation 12, 1985-2004. http://dx.doi.org/10.1023/A:1024103527611 [ Links ]
Samways, M.J. & Lu, S-S., 2007, 'Key traits in a threatened butterfly and its common sibling: Implications for conservation', Biodiversity and Conservation 16, 40954107. http://dx.doi.org/10.1007/s10531-007-9209-z [ Links ]
Samways, M.J., McGeoch, M.A. & New, T.R. (eds.), 2010, Insect conservation: A handbook of approaches and methods, Techniques in Ecology & Conservation Series, Oxford University Press, Oxford. PMCid:PMC2847917 [ Links ]
Schtickzelle, N., Turlure, C. & Baguette, M., 2007, 'Grazing management impacts on the viability of the threatened bog fritillary butterfly Proclossiana eunomia', Biological Conservation 136, 651-660. http://dx.doi.org/10.1016/j.biocon.2007.01.012 [ Links ]
Schultz, C.B. & Dlugosch, K.M., 1999, 'Nectar and hostplant scarcity limit populations of an endangered Oregon butterfly', Oecologia 119, 231-238. http://dx.doi.org/10.1007/s004420050781 [ Links ]
Smee, M., Smyth, W., Tunmore, M., Ffrench-Constant, R. & Hodgson, D., 2011, 'Butterflies on the brink: Habitat requirements for declining populations of the marsh fritillary (Euphydryas aurinia) in SW England', Journal of Insect Conservation 15, 153-163. http://dx.doi.org/10.1007/s10841-010-9334-y [ Links ]
Spatial data, 2013, Biodiversity Spatial Planning and Information Division, Ezemvelo KwaZulu-Natal Wildlife, South Africa. [ Links ]
Swengel, S.R., Schlicht, D., Olsen, F. & Swengel, A.B., 2011, 'Declines of prairie butterflies in the midwestern USA', Journal of Insect Conservation 15, 327-339. http://dx.doi.org/10.1007/s10841-010-9323-1 [ Links ]
Thomas, J.A., 2005, 'Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups', Philosophical Transactions of the Royal Society B 360, 339-357. http://dx.doi.org/10.1098/rstb.2004.1585, PMid:15814349, PMCid:PMC1569450 [ Links ]
Tudor, O., Dennis, R.L.H., Greatorex-Davis, J.N. & Sparks, T.H., 2004, 'Flower preferences of woodland butterflies in the UK: Nectaring specialists are species of conservation concern', Biological Conservation 119, 397-403. http://dx.doi.org/10.1016/j.biocon.2004.01.002 [ Links ]
Watson, J., 2001, 'How to Determine a Sample Size: Tipsheet #60', Penn State Cooperative Extension, University Park, PA, viewed 14 May 2012, from http://www.extension.psu.edu/evaluation/pdf/TS60.pdf [ Links ]
 Correspondence:
Correspondence:
Adrian Armstrong
Postal address: PO Box 13053
Cascades 3202, South Africa
Email: aarmstrong@kznwildlife.com
Received: 05 Mar. 2013
Accepted: 17 Aug. 2013
Published: 10 Oct. 2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}