










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.55 n.1 Pretoria Jan. 2013
ORIGINAL RESEARCH
Long-term stability of grazing lawns in a small protected area, the Mountain Zebra National Park
Peter Novellie; Angela Gaylard
Conservation Services Division, South African National Parks, South Africa
ABSTRACT
We examined a heavily grazed plant community dominated by creeping grass species with the aim of, (1) determining its response to the exclusion of grazing and (2) its long-term persistence. This plant community was particularly favoured by wild ungulate species that prefer short grasses - blesbok (Damaliscus dorcas phillipsi), springbok (Antidorcas marsupialis) and black wildebeest (Connochaetes gnou). Exclusion of grazing by large herbivores by means of fencing resulted in the virtual disappearance of the creeping grasses and their replacement by tall tufted species. On plots that remained unfenced, the plant species composition was found to be little changed after an interval of more than 20 years. The number large stock unit equivalents (LSU) per ha carried by the plant community was used as a proxy for grazing intensity. Monitored for approximately 2 years at the start of the study, LSU per ha was found to greatly exceed levels recommended for commercial livestock production. This plant community conforms to a recently published definition of a grazing lawn, in that intense grazing promotes palatable, grazing-tolerant grass species.
Conservation implications: The positive association between grazers and grazing-tolerant grass species evidently persisted for more than 20 years and there was no evidence of an increase in abundance of unpalatable plant species. Despite the small size of the park, which limited the extent of large herbivore movements, localised heavy grazing did not lead to range degradation.
Introduction
The term 'grazing lawn' has been widely used to describe patches that, as a result of frequent, intense grazing, become dominated by short, grazing-tolerant grass species (Archibald 2008; Archibald et al. 2005; Cromsigt & Olff 2008; McNaughton 1984; Stock, Bond & Van de Vijver 2009; Waldram, Bond & Stock 2007). In a detailed review of this phenomenon, Cromsigt and Kuijper (2011:212) defined a grazing lawn as 'a grazed or browsed patch in which intense foraging leads to increased resource availability and a proportional increase in palatable species in the patch'. Thus, grazing lawns are characterised by a positive association between grazers and grazing-tolerant grasses; intensive grazing has the effect of increasing the productivity and quality of the food resource (Cromsigt & Kuijper 2011). It has been demonstrated (Archibald 2008; Cromsigt & Olff 2008; Stock et al. 2009; Waldram et al. 2007) that grazing lawns do not derive solely from existing matrices of abiotic factors, such as soil heterogeneity (although their distribution may be influenced by such factors), but are animal driven. Consequently, they can increase or decrease depending on fire, rainfall and grazing regime.
The formation of a shifting mosaic of grazing lawns together with areas of taller grass appears to be a key determinant of spatial heterogeneity of food resources that is necessary to maintain a diversity herbivore species (Cromsigt & Olff 2008; Fuhlendorf & Engle 2001; Pickett, Cadenasso & Benning 2003) and is important for maintaining biological diversity and stability in grasslands (Tilman & Downing 1994). From this perspective, patch-selective grazing and grazing lawn formation should be regarded as a desirable phenomenon in grassland systems that are managed with the objective of maintaining biodiversity and natural ecological processes.
However, concern has been expressed about possible negative effects of patch-selective grazing (Fuls 1992; Kellner & Bosch 1992). It is conceivable that, under certain conditions, protracted use of individual patches may reach a point where the positive association between grazers and palatable grasses breaks down and the lawn transforms to a state that can no longer support grazing. This may be a particular risk in small protected areas, where fences restrict the spatial scale over which grazing dynamics can play out (Owen-Smith 1983). Under these conditions, patch-selective grazing may not result in a stable (or at least resilient) mosaic of grazing lawns, but may instead cause progressive depletion of the grazing resource.
To avoid range deterioration in small protected areas, it has been proposed that stocking rates of wild ungulate species should be maintained in accordance with norms set for commercial livestock production (Meissner 1982). However, if grazing lawns are prone to transform to a different state, then the setting of stocking rates for the area as a whole may be ineffective in preventing such change (Kellner & Bosch 1992).
It is therefore important to understand the drivers of grazing lawn formation in small protected areas and to monitor the stability of the grazing lawn mosaic. The key question is whether the lawns retain their attractiveness to grazing ungulates over the long term, or whether they eventually transform to a state, either through soil loss or encroachment by unpalatable species, which no longer provides grazing.
The Mountain Zebra National Park (MZNP) is a relatively small protected area, which carries localised high densities of grazing ungulates, some of which strongly favour grazing lawns (Novellie 1990). This study was conducted on some of the most heavily used grazing lawns in the MZNP, examining (1) their long-term persistence and (2) response to the exclusion of grazing by means of fenced exclosures.
Research method and design
Setting
The MZNP is situated 24 km west of the town of Cradock in the Eastern Cape Province, South Africa. At the time when the observations commenced (1987), the park was only 6536 ha. In the late 1990s it was greatly expanded in size, extending over 28 412 ha at the conclusion of the study. Monthly minimum and maximum temperatures vary from 6 °C to 28 °C in summer (from September to March) and from 0 °C - 20 °C in winter (from April to August) (Brown & Bezuidenhout 2000). Rainfall averages about 400 mm, with most (70%) falling in the summer months. Periodic light snow occurs during the winter months and frost is common between May and October.
The park lies in a transition zone, incorporating elements of three biomes, the Nama-Karoo, Grassland and Albany Thicket. It includes three of the vegetation types recognised by Mucina and Rutherford (2006): the Eastern Upper Karoo, Karoo Escarpment Grassland and Eastern Cape Escarpment Thicket. This study was conducted in Karoo Escarpment Grassland in a valley near the western boundary of the park, near two small impoundments called the Oudamme. Although fires occur periodically in the higher Karoo Escarpment Grassland of the park, the areas of grazing lawn seldom, if ever, burn. No fires occurred in the Oudamme Valley between 1987 and 2010.
Data on habitat use by grazing ungulates (Novellie 1990, 1991) revealed that the Oudamme Valley was one of the most heavily grazed areas of the park during the late 1980s. This area was therefore selected for a more detailed assessment of the long-term stability of the grazing lawns.
Procedure
In March and April of 1987, four matched pairs of plots, each roughly 30 m by 30 m in size, were laid out in the heavily grazed areas of the Oudamme Valley. In each plot a 200-point survey for canopy spread cover was conducted (see Novellie & Strydom 1987 for further details). The points were arranged in eight rows of 25 each, with 1-m spacing between points and 1-m or 2-m spacing between rows. A canopy spread strike was recorded if a point fell within an imaginary line drawn around the perimeter of the canopy of a plant. For each strike the species of plant was recorded. One member of each pair of plots was then selected at random and a fence approximately 1.5 m high was erected around it to exclude grazing by large herbivores. The other member of each pair remained exposed to grazing. Using the same method, the surveys of two of the pairs of plots were repeated in June 2009. The surveys of the remaining two pairs were completed in June 2010. The plant species names reported in this article follow Klopper et al. (2006). Canonical correspondence analysis was used to investigate changes from 1987 to 2009-2010 in plant species composition in the fenced and unfenced plots (Ter Braak 1987).
Between 1987 and 2009-2010 no formal surveys were conducted, but the plots were visited every second or third year (by the first author) to get a general impression of the state of the vegetation.
An indication of the intensity of use of the habitat by large herbivores was obtained by conducting five to ten counts per month from 1987 to 1989. Ideally, the density of animals should have been sampled exactly within the four 30 m by 30 m plots that were left unfenced, but at this small scale a very large number of counts would have been needed to get a reliable indication of average animal density. It was therefore necessary to conduct the animal counts in larger-sized plots. Two plots, each 100 m by 100 m (1 ha) were identified in similar habitat, situated about 200 m apart from one another and adjacent to the four matched pairs of plots used for the exclosure study. The corners were marked with 1.8 m high steel fencing standards which could be seen through binoculars by an observer on the ridge overlooking the Oudamme Valley. Thus, the animals within the boundaries of the plots could be counted without disturbing them.
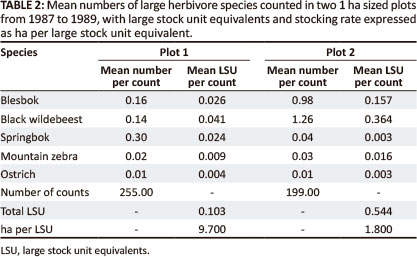
The canopy spread cover of plant species within the two 1 ha plots was determined by 200-point surveys, as described above. The 200-point surveys were replicated at randomly chosen locations within each 1 ha plot, ten surveys in one plot and eight in the other. (The results of these surveys are also reported by Novellie 1990). In total, one of the 1 ha plots was checked 199 times and the other 255 times. For comparison with Department of Agriculture stocking rate standards as applied during the 1980s, the average numbers of ungulates in each plot were converted to large stock unit equivalents (LSU), according to the method of Meissner (1982), as published in the Government Gazette (Republic of South Africa 1985).
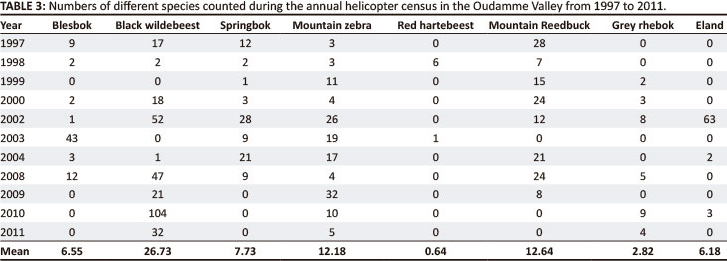
Animal counts in the 1 ha plots were not continued after 1989, but the locations of animals plotted during the annual helicopter census (Gaylard 2011) provided a rough indication of use of the Oudamme Valley by large herbivores between 1997 and 2011.
Results
Table 1 shows the mean canopy spread cover of grass species in the four matched pairs of plots and each of the two 1 ha plots at the start of observations in 1987. The species composition was broadly similar across all plots confirming that they were representative of the same plant community. The most abundant grasses in terms of canopy spread cover were the creeping species Cynodon incompletus and Tragus koelerioides. The former grew in dense lawns around accumulations of dung but elsewhere occurred generally as sparse runners. In contrast, T. koelerioides was not closely associated with dung patches and occurred as sparse runners rather than lawns. The bunch grasses Merxmuellera disticha, Digitaria eriantha and Eragrostis curvula were next most abundant in terms of cover. Merxmuellera disticha was very patchy in distribution and the tall, dense tufts showed relatively little signs of grazing. In contrast, D. eriantha and E. curvula were widely distributed and virtually all tufts of these species were cropped to a height of around 5 cm. The annual grass Aristida congesta was widespread but its cover was generally low. The taller bunch grasses: Themeda triandra, Cymbopogon pospischilii and Heteropogon contortus were rare throughout the Oudamme Valley, the only specimens seen at that time were growing within the canopy of shrubs where they were protected against grazing.
Figure 1 and Figure 2 compare the fenced and matched unfenced plots in the years 1987 and 2009-2010. Only the most abundant and widespread grass (Figure 1) and dwarf shrub (Figure 2) species are shown, as the others were either too uncommon or too localised to draw conclusions as to their response to the exclusion of grazing.

The canonical correspondence analysis showed that species composition was significantly associated with treatments, with the strongest association for the fenced plots in 2009-2010 (F = 9.77; p = 0.002). The ordination diagram (Figure 3) shows how the species were clustered with reference to the treatments. Of the grass species, T. triandra, C. pospischilii and H. contortus formed a group that was very strongly associated with the exclosures as surveyed during 2009-2010. In clear contrast to this group, the exclosure plots were characterised by a lack of the annual grass A. congesta and the creeping grasses C. incompletus and T. koelerioides. The bunch grasses E. curvula and D. eriantha were in an intermediate position in that they were associated with all plots, although they both showed a relative increase in cover in the fenced plots in 2009-2010 relative to the other plots (Figure 1).

In summary, the fenced plots in 2009-2010 showed a complete transformation in grass species composition; the creeping and annual species all but disappeared and taller bunch grass species were dominant (Figure 1). On the other hand, the unfenced plots showed relatively minor changes in grass species composition from 1987 to 2009-2010 (Figure 1).
In contrast to the grasses, none of the common dwarf shrub species showed a consistent response to fencing, instead differences are apparent between the survey years (Figure 2 and Figure 3). Chrysocoma ciliata is associated with both the fenced and unfenced plots in 1987, but in 2009-2010 it declined in all plots. Helichrysum dregeanum is associated with unfenced and fenced plots, but only in 2009-2010. Felicia fascicularis increased in the unfenced plots, but there was considerable between-plot variation in the case of the fenced plots. The reason for the changes in abundance in these dwarf shrub species from 1987 to 2009-2010 is unknown.
Over the period 1987-1989, both of the 1 ha plots were utilised mainly by short grass-grazers, blesbok (Damaliscus dorcas phillipsi), black wildebeest (Connochaetes gnou) and springbok (Antidorcas marsupialis). Mountain zebra (Equus zebra zebra) and ostrich (Struthio camelus) were observed there comparatively infrequently (Table 2). Red hartebeest (Acelaphus buselaphus), also common in the park at the time, were never observed in these plots. The stocking rate carried by Plot 1 was estimated at 9.7 ha per LSU, whilst that of Plot 2 was 1.8 ha per LSU. The difference in animal density between the two may have been caused by the fact that there was more surface rock in Plot 1; hence, the cover of short grass favoured by grazers was lower in Plot 1 (mean ± standard error: 16.0% ± 2.2%) than in Plot 2 (28.0% ± 2.7%). For both plots, the stocking rates are considerably higher than the 14 ha per LSU recommended for the area by the Grootfontein Agricultural Development Institute (1986).
The census results (Table 3) indicated that the Oudamme Valley continued to support fair numbers of large herbivores from 1997 to 2011. It was, however, not possible to be sure from the census results of the exact plant community being utilised, as the animals are likely to have moved around in response to the helicopter. This is probably the reason why Table 3 indicates a relatively greater abundance of mountain zebra and mountain reedbuck, species which tend to favour the ridges around the valley rather than the valley bottom habitat in the 1 ha plots. For this reason, we did not attempt to estimate animal densities from the census results.
Discussion
The results described above are in accordance with evidence in the literature (Archibald 2008; Cromsigt & Olff 2008; Stock et al. 2009; Waldram et al. 2007) that grazing lawns are animal driven; if grazing is removed the creeping grasses give way to taller bunch grasses.
The change in species composition in the exclosures follows the successional sequence predicted by Vorster's (1982) veld condition index for the Karoo Region, which classifies T. triandra, C. pospischilii, H. contortus and D. eriantha as 'decreaser' species, which decrease in abundance if subjected to heavy grazing, whereas E. curvula, C. incompletus and T. koelerioides are 'increaser' species which are typically promoted by grazing. In terms of Vorster's index, which is designed as a guide to commercial pastoralists, the proliferation of the increaser species at the expense of decreaser species is regarded as a sign of deterioration of veld condition, something to be rectified by reducing stocking densities. This is obviously not true for natural systems, where grazing lawns of increaser species are the product of a positive association between consumers and the resource. This is evident from the fact that blesbok, springbok and black wildebeest showed a strong preference for communities dominated by increaser species (Novellie 1990) and occupied these habitat patches at relatively high densities. Moreover, the increaser species are highly palatable to the short-grass grazers, as is evident from the extent to which they were defoliated in the study area.
It is noteworthy that, although all the fenced plots were dominated by one or other bunch grass species, there were differences between the plots in the particular grass species that eventually came to dominate; in some it was T. triandra, in others C. pospischilii or D. eriantha. Given that the tufted grasses were extremely rare in the area before fencing, it seems likely that this was caused by chance variation between plots in the nearest sources of colonising seeds.
For this plant community (described as Lower Slope Degraded Dwarf Shrubland by Van der Walt 1980), the term 'grazing lawn' is perhaps a misnomer, because it is only C. incompletus that forms actual lawns and these are generally no more than a few square metres in area immediately surrounding dense patches of ungulate dung. Nevertheless, we regard it as appropriate to refer to the C. incompletus -T. koelerioides community as a whole - rather than specifically the Cynodon lawns - as a grazing lawn, as it conforms to Cromsigt and Kuijper's (2011:212) definition of a lawn in that 'intense foraging leads to increased resource availability and a proportional increase in palatable species in the patch'.
The most common dwarf shrub species associated with the grazing lawns, F. fascicularis and H. dregeanum, are regarded as being palatable to domestic stock by Vorster (1982) and, as noted, frequently showed signs of browsing in the Oudamme Valley. It is interesting to consider whether this community may qualify as a browsing as well as a grazing lawn (as defined by Cromsigt & Kuijper 2011). However, these dwarf shrubs persisted in the exclosures (Figure 2) so, unlike the creeping grasses, their presence does not appear to be dependent on grazing.
We did not quantitatively measure the rate at which the transformation in plant species composition within the exclosures took place, but the periodic inspections during the study period suggested that it occurred very slowly. Little change in species composition could be discerned during the first 5 years after construction of the exclosures (P. Novellie, pers. obs.). As noted, the tall grass species that came to dominate the exclosures were initially very uncommon in the area, so the slow transition may have been due to scarcity of seed.
In the MZNP, it appears that the mechanism driving the formation of these grazing lawns is the patch-selective grazing of blesbok, springbok and black wildebeest. The habit of the territorial males of these species of occupying on 'stamping grounds' and of defecating and urinating at specific localities (Du Plessis 1972; Lynch 1971; Novellie 1975, 1990; Von Richter 1972), results in accumulations of dung and associated Cynodon lawns. Termite mounds are present in the Oudamme Valley and may also play a facilitating role in influencing the formation of grazing lawns (see Cromsigt & Olff 2008). Du Plessis (1972) describes how a blesbok male, introduced into a paddock, initially focused his grazing around the margins of a termite mound, eventually creating a patch of closely grazed grass. Megaherbivores, such as white rhino, have been demonstrated to play a key role in creating grazing lawns in Hluhluwe-iMfolozi Park (Cromsigt & Olff 2008; Waldram et al. 2007), but megaherbivores are absent from the MZNP.
Conclusion
Although there has been concern around the possibility that the grazing lawns may transform into an ungrazable state, it appears that the plant species composition on the grazed plots showed little change 20 years after the initial surveys, despite continued use of the area by the short-grass grazers. Not having conducted interim descending point surveys we are uncertain as to fluctuations in species composition that may have occurred during the 20 year interval. However, no sign of transformation could be detected during the visits to the study area made about every 2-3 years; in general, the appearance the plant species composition appeared to be very stable. There was never any indication of dominance by the species of annual grasses and unpalatable dwarf shrubs (e.g. A. congesta and C. ciliata) that were identified by Vorster (1982) as characterising the worst state of veld degradation in the Karoo Region.
Thus, no evidence could be detected of a breakdown of the positive association between grazers and the creeping grasses. However, this is not to say that it could not occur under certain circumstances. In small protected areas it is highly desirable to develop a predictive understanding of the relationship between the overall stocking rate maintained, on the one hand, and grazing lawn stability and/or resilience on the other.
Acknowledgements
We are grateful to Park Manager, Ettienne Fourie, for constructing and maintaining the exclosures in good condition over many years, to Section Ranger, Robyn Wood, and Student Ranger, Nonkazimlo Baca, for able assistance with the vegetation surveys and to Animal Ecologist, Guy Castley, for historical air census data. An earlier draft of the article was greatly improved by the comments of two anonymous reviewers.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
P.N. (South African National Parks) conducted the on-the-ground field surveys and wrote the manuscript, whilst A.G. (South African National Parks) collected air census data, conducted the statistical analysis and made conceptual contributions.
References
Archibald, S., 2008, 'African grazing lawns - How fire, rainfall, and grazer numbers interact to affect grass community states', Journal of Wildlife Management 72(2), 492-501. http://dx.doi.org/10.2193/2007-045 [ Links ]
Archibald, S., Bond, W.J., Stock, W.D. & Fairbanks, D.H.K., 2005, 'Shaping the landscape: Fire-grazer interactions in an African savanna', Ecological Applications 15, 96-109. http://dx.doi.org/10.1890/03-5210 [ Links ]
Brown, L.R. & Bezuidenhout, H., 2000, 'The phytosociology of the farm De Rust in the Mountain Zebra National Park, Eastern Cape', Koedoe 43(1), 1-18. http://dx.doi.org/10.4102/koedoe.v43i1.204 [ Links ]
Cromsigt, J.P.G.M. & Kuijper, D.P.J., 2011, 'Revisiting the browsing lawn concept: Evolutionary Interactions or pruning herbivores?', Perspectives in Plant Ecology, Evolution & Systematics 13, 207-215. http://dx.doi.org/10.1016/j.ppees.2011.04.004 [ Links ]
Cromsigt, J.P.G.M. & Olff, H., 2008, 'Dynamics of grazing lawn formation: An experimental test of the role of scale-dependent processes', Oikos 117, 1444-1452. http://dx.doi.org/10.1111/j.0030-1299.2008.16651.x [ Links ]
Du Plessis, S.S., 1972, 'Ecology of blesbok with special reference to productivity', Wildlife Monographs 30, Wildlife Society, Inc., Bethesda. [ Links ]
Fuhlendorf, S.D. & Engle, D.M., 2001, 'Restoring heterogeneity on rangelands: Ecosystem management based on evolutionary grazing patterns', BioScience 51(8), 625-632. http://dx.doi.org/10.1641/0006-3568(2001)051[0625:RHOREM]2.0.CO;2 [ Links ]
Fuls, E.R., 1992, 'Semi-arid and arid rangelands: A resource under siege due to patch-selective grazing', Journal of Arid Environments 22, 191-193. [ Links ]
Gaylard, A., 2011, 'Aerial survey report for the Mountain Zebra National Park', unpublished report, South African National Parks. [ Links ]
Grootfontein Agricultural Development Institute, 1986, 'Grazing capacity zones for the Karoo Region', Grootfontein Agricultural Development Institute, Middelburg. [ Links ]
Kellner, K. & Bosch, O.J.H., 1992, 'Influence of patch formation in determining the stocking rate for southern African grasslands', Journal of Arid Environments 22, 99-105. [ Links ]
Klopper, R.R., Chatelain, C., Banninger, V., Habashi, C., Steyn, H.M., De Wet, B.C. et al., 2006, 'Checklist of the flowering plants of Sub-Saharan Africa. An index of accepted names and synonyms', South African Botanical Diversity Network Report 42, SABONET, Pretoria. [ Links ]
Lynch, C.D., 1971, 'A behavioural study of the blesbok, Damaliscus dorcas phillipsi, with special reference to territoriality', Unpublished MSc. thesis, Department of Zoology, University of Pretoria. [ Links ]
McNaughton, S.J., 1984, 'Grazing lawns: Animals in herds, plant form, and coevolution', The American Naturalist 124(6), 863-886. http://dx.doi.org/10.1086/284321 [ Links ]
Meissner, H.H., 1982, 'Theory and application of a method to calculate forage intake of wild southern African ungulates for purposes of estimating carrying capacity', South African Journal of Wildlife Research 12, 41-47. [ Links ]
Mucina, L. & Rutherford, M.C. (eds.) 2006, 'The vegetation of South Africa, Lesotho and Swaziland', Strelitzia 19, South African National Biodiversity Institute, Pretoria. [ Links ]
Novellie, P.A., 1975, 'Comparative social behaviour of springbok and blesbok on the Jack Scott Nature Reserve, Transvaal', unpublished MSc. thesis, Department of Zoology, University of Pretoria. [ Links ]
Novellie, P.A., 1990, 'Habitat use by indigenous grazing ungulates in relation to sward structure and veld condition', Journal of the Grassland Society of Southern Africa 7, 16-23. [ Links ]
Novellie, P.A., 1991, 'Seasonal movements and habitat use by African grazing ungulates in a small conservation area', in Proceedings of the IVth International Rangeland Congress, vol. 2, pp. 709-713, Association Frangaise de Pastoralisme, Montpellier, France. [ Links ]
Novellie, P.A. & Strydom, G., 1987, 'Monitoring the response of vegetation to use by large herbivores: An assessment of some techniques', South African Journal of Wildlife Research 17, 109-117. [ Links ]
Owen-Smith, R.N., 1983, 'Dispersal and the dynamics of large herbivores in enclosed areas: Implications for management', in R.N. Owen-Smith (ed.), Management of large mammals in African conservation areas, pp. 127-143, HAUM Educational Publishers, Pretoria. [ Links ]
Pickett, S.T.A., Cadenasso, M.L. & Benning, T., 2003, 'Biotic and abiotic variability as key determinants of savanna heterogeneity at multiple temporal scales', in J.T. du Toit, K.H. Rogers & H.C. Biggs (eds.), The Kruger experience: Ecology and management of savanna heterogeneity, pp. 23-40, Island Press, Washington, DC. [ Links ]
Republic of South Africa, 1985, 'Government Notice R. 2687', Government Gazette 10029 (06 December)', Government printers, Pretoria. [ Links ]
Stock, W.D., Bond, W.J. & Van de Vijver, C.A.D.M., 2009, 'Herbivore and nutrient control of lawn and bunch grass distributions in a southern African savanna', Plant Ecology 206(1), 15-27. http://dx.doi.org/10.1007/s11258-009-9621-4 [ Links ]
Ter Braak, C.J.F., 1987, 'The analysis of vegetation-environment relationships by canonical correspondence analysis', Plant Ecology 69, 69-77. http://dx.doi.org/10.1007/BF00038688 [ Links ]
Tilman, D. & Downing, J.A., 1994, 'Biodiversity and stability in grasslands', Nature 367, 363-365. http://dx.doi.org/10.1038/367363a0 [ Links ]
Van der Walt, P.T., 1980, 'A phyto-sociological reconnaissance of the Mountain Zebra National Park', Koedoe 23, 1-32. http://dx.doi.org/10.4102/koedoe.v23i1.632 [ Links ]
Von Richter, W., 1972, 'Territorial behaviour of the black wildebeest Connochaetes gnou', Zoologica Africana 7, 207-231. [ Links ]
Vorster, M., 1982, 'The development of the ecological index method for assessing veld condition in the Karoo', Proceedings of the Grassland Society of southern Africa 17, 84-89. [ Links ]
Waldram, M., Bond., W. & Stock, W., 2007, 'Ecological engineering by a mega-grazer: White rhino impacts on a South African savanna', Ecosystems 11, 101-112. http://dx.doi.org/10.1007/s10021-007-9109-9 [ Links ]
 Correspondence:
Correspondence:
Peter Novellie
PO Box 787
Pretoria 0001, South Africa
peter.novellie@sanparks.org
Received: 15 Aug. 2013
Accepted: 07 June 2013
Published: 13 Aug. 2013


{kind=link}
{kind=link}
{kind=link}