










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkKoedoe
versión On-line ISSN 2071-0771
versión impresa ISSN 0075-6458
Koedoe vol.55 no.1 Pretoria ene. 2013
ORIGINAL RESEARCH
Virgilia divaricata may facilitate forest expansion in the afrotemperate forests of the southern Cape, South Africa
Corli Coetsee; Benjamin J. Wigley
School of Natural Resource Management, Nelson Mandela Metropolitan University, George campus, South Africa
ABSTRACT
Virgilia divaricata is a fast-growing nitrogen-fixing tree species often found on the margins of forest in the southern Cape of South Africa and is particularly abundant after fire. However, V. divaricata may invade fynbos even in the absence of fire and it has been described as a forest precursor. We investigated whether V. divaricata enriches soil fertility after its invasion into fynbos areas adjacent to forests. We measured soil organic carbon and soil nutrients at four sites. At each site, three vegetation types (forest, V. divaricata and fynbos) were examined on the same soil type and at the same elevation. Our results showed that, on average, soils taken from V. divaricata stands had higher nitrogen and phosphorus values than the adjacent fynbos soils, with either lower or similar values to the adjacent forest soils. Higher soil fertility under V. divaricata, together with their shading effect, may create conditions favourable for shade-loving forest species dependent on an efficient nutrient cycle in the topsoil layers, and less favourable for shade-hating fynbos species, which are generally adapted to low soil fertility. We suggest that the restoration of the nutrient cycle found in association with forest may be accelerated under V. divaricata compared with other forest precursor species, which has important consequences for the use of V. divaricata in ecosystem restoration.
Conservation implications: Alien plantations in the Outeniqua Mountains are being phased out and the areas are being incorporated into the Garden Route National Park. Fynbos areas are increasingly being invaded by forest and thicket species owing to fire suppression in lower-lying areas. An improved understanding of the fynbos-forest boundary dynamics will aid in efficient management and restoration of these ecosystems.
Introduction
In South Africa, natural forest covers about half a million hectares, which equates to only 0.5% of the land surface. Apart from being the smallest biome, forest is also the most fragmented and most vulnerable biome within the country (Van der Merwe 2011). The largest forest complex is the afrotemperate Knysna forest, which covers 65 000 ha of the southern Cape. Heathlike vegetation that is adapted to frequent fire, known locally as fynbos, forms a matrix with these forests (Manders, Richardson & Masson 1992). Fynbos is evergreen and sclerophyllous. Geldenhuys (1994) has shown that although environmental factors such as rainfall and substrate determine the potential limits of forest distribution, actual forest distribution is determined by the interaction between prevailing winds during dry periods, terrain physiography and the effect of this interaction on fire pattern. In our study area, forests are usually found in the topographic shadow areas of warm and desiccating berg winds, which are common in autumn and winter (Geldenhuys 1994). However, extensive plantation forestry, which has protected fynbos against fire, has led to forest expansion in the southern Cape in recent times (C. Geldenhuys, pers. comm., January 2011). In the prolonged absence of fire, forest species start to invade the surrounding fynbos and the dominant vegetation may switch to closed-canopy forest (Kruger 1984; Masson & Moll 1987).
Forest trees face certain inhibitory factors when expanding into fynbos. These include the lack of perches for birds, which inhibits seed dispersal, the inability of seedlings to survive summer droughts, lack of protection from wind and radiation and low levels of soil nutrients (Campbell & Killick 1985; Manders 1990; Manders & Richardson 1992; White 1978). Manders and Richardson (1992) propose a model of forest development in fynbos where the mechanism of succession involves facilitation. Forest precursor species act as 'nurse plants' by increasing the attractiveness of the area to avian dispersers, ameliorating summer water deficits owing to a mulching effect of litter, protecting forest trees from radiation and wind, and decreasing competition for water owing to decreased cover beneath these plants. The concept of pioneer plants acting as nurse plants and 'nucleation' sites for subsequent woody vegetation development has been reported in many ecosystems (e.g. Barnes & Archer 1999; Yarranton & Morrison 1974). Such modification of the environment by an organism has been referred to as 'niche construction' or 'ecosystem engineering' (Crain & Bertness 2006; Laland, Odling-Smee & Feldman 1999; Odling-Smee, Laland & Feldman 1996). Examples include changing the chemical nature of soils, the pattern of nutrient cycling, the temperature, humidity, fertility, acidity and salinity of soils, and the patterns of shade in the habitat (Laland et al. 1999).
Virgilia divaricata Adamson and the closely related Virgilia oroboides (P.J. Bergius) Salter (considered as one species by some authors and locally known as keurboom) are commonly observed growing on forest margins in the southern Cape (Phillips 1926). Virgilia spp. seeds can be viable for at least 230 years, enabling the plant to recover or regenerate rapidly after fire (Geldenhuys 1994). This fast-growing tree can establish without shade. Seedlings and saplings of forest canopy species can usually be found 5-10 years after the development of Virgilia spp. stands and become established after the natural suppression, mortality and consequent thinning of the pioneer stand (Geldenhuys 1994). Phillips (1926) has shown that the growth of seedlings and saplings of forest species co-dominant with Virgilia spp. is considerably more rapid under Virgilia spp. stands than in normal climax forest. The mechanism of this enhanced growth was not investigated; however, Phillips (1926) suggests that it may be related to light conditions under Virgilia spp. being more conducive to sapling growth than the low light conditions beneath forest.
We suggest that an alternative mechanism for facilitation of forest sapling growth may be that V. divaricata increases soil fertility, as both Virgilia spp. are members of Fabaceae, which have root nodules containing nitrogen-fixing bacteria (Palgrave 1981). Nitrogen-fixing alien acacias, which regularly invade fynbos, have been shown to increase soil nutrient mineral content (Musil 1993; Stock, Wienand & Baker 1995; Witkowski 1991a; Yelenik, Stock & Richardson 2004). However, little prior research has considered the effects of V. divaricata on soil fertility and the current study therefore aimed to investigate whether the establishment of V. divaricata in fynbos patches adjacent to forests enhances soil fertility. We hypothesised that soils underneath the legume V. divaricata will become nitrogen enriched as a result of the addition of nitrogen-rich litter and that the higher nitrogen levels will be perpetuated by forest development. We also hypothesised that levels of other nutrients, including carbon, will increase as forest establishment progresses.
Methods
Study site
The study sites were situated in the Garden Route National Park in the Western Cape: two on the northern edges of the Groeneweide Forest, 5 km east of George (Groenkop 1: 33.939436° S 22.560760° E; Groenkop 2: 33.943334° S 22.562730° E) and two in the Tsitsikamma Bloukrans forest area near Nature's Valley (Plateau: 33.565690° S 23.45856° E; Covie: 33.943334° S 22.562730° E). The average rainfall varies between 500 mm and 1200 mm, with peaks in autumn and early summer and a minimum in December. Temperatures are mild, with mean minima and maxima of 7 °C and 19 °C, respectively, in June and 15 °C and 26 °C, respectively, in January (Bond 1981). Soils are derived from quartzitic sandstone of the Table Mountain Group. The topsoil in the Groenkop sites is between 40 cm and 50 cm deep and overlays poorly drained subsoil with indications of a fluctuating water level, more so at Groenkop 1 than Groenkop 2. The two Tsitsikamma sites were situated on old dunes. (See Geldenhuys [1982, 1991] for a thorough environmental description of the area.)
The forests studied fall within the Southern Afrotemperate forest vegetation type (Mucina & Rutherford 2006). The surrounding vegetation is mostly fynbos, with South Outeniqua Sandstone Fynbos bordering the Groeneweide forest and Tsitsikamma Sandstone Fynbos in the Tsitsikamma sites (Mucina & Rutherford 2006). The study sites at Groeneweide (Groenkop 1 and 2) burnt in 1996 and well-established adult V. divaricata trees now form mostly consocies stands, approximately 10 m - 15 m high at the ecotone between forest and fynbos. The fynbos at Covie is dominated by young Leucadendron saplings, indicative of a recent fire, with a narrow band of V. divaricata between forest and fynbos areas. The fynbos at Plateau is mostly lacking in Proteaceae, probably because the area now conserved as a nature reserve has been used for (potato) farming until quite recently. The V. divaricata bordering the forest at Plateau established after a fire in 2005. Finding sites where the fynbos was in pristine condition was a typical problem during site selection. In the past, fynbos adjacent to forest was either farmed or planted with exotic plantations. Currently, fire suppression, especially in the lower-lying fynbos patches, has led to encroachment by thicket species, forest precursors or invasive alien trees.
Sample collection and analyses
At each of the four sites, soils were collected from within the forest, adjacent fynbos vegetation and V. divaricata stands. Care was taken to select sites where forest, fynbos and V. divaricata occurred at the same elevation and on the same soil type. At each site, the three treatments (forest, fynbos and V. divaricata) were close to one another. Adult stands of V. divaricata formed narrow strips (~20 m - 80 m) of consocies with an understorey of sparsely distributed forest saplings on the margin between forest and fynbos. We tried to avoid areas where Pteridium aquilinum (bracken) occurred as we were uncertain how the strong allelopathic substances exuded by bracken would affect soils (Chapman & Reiss 1992); however, bracken occurred in quite dense patches in some areas adjacent to V. divaricata, especially at Plateau. Soil was collected on transects spanning across the V. divaricata stands and on transects running parallel to either side of the V. divaricata stands in forest and fynbos. Soil samples were collected about 20 m - 30 m apart, with a soil auger at three depths: 0 cm - 10 cm, 10 cm - 20 cm and 20 cm - 50 cm (4 sites χ 3 vegetation types χ 3 depths χ 5 repetitions = 180 samples). After collection, soils were sieved through a 2-mm sieve to remove organic material and roots, and air-dried for several days.
All samples were analysed for organic carbon, nitrogen, phosphorus, calcium, magnesium, sodium and potassium at the Institute for Plant Production, Stellenbosch, South Africa. Carbon content was determined by a rapid dichromate oxidation method using the Walkley-Black procedure (Walkley 1947). Samples were not pre-treated for carbonates as the pH was well below 7.4, which suggested that carbonates were not present in significant amounts in our samples (Nelson & Sommers 1996). Repeatedly run samples analysed for carbon had a standard deviation of 0.03% and coefficient of variation (CV) of 0.07. Extractable phosphorus, calcium, magnesium, sodium and potassium were extracted with 1% citric acid and analysed by using a Thermo ICP iCAP 6000 Series Spectrometer (Thermofisher Scientific, Surrey, UK). Total soil nitrogen was analysed by total combustion at Bemlab (Pty) Ltd., Somerset West, South Africa, using a Leco Nitrogen Analyser FP 528 (LECO Corporation, St. Joseph, USA).
To determine bulk density, a known volume was removed from each soil depth using a core sampler. Known volumes were dried, weighed and sieved to remove all roots and stones, and the weight and volume of rocks were measured. The bulk density of each sample was calculated by dividing the dry mass of soil by the volume of soil, with the mass of the rocks subtracted in each case. Bulk density was used to convert concentration data to content data (i.e. aerial data). We underestimated bulk density for samples with high root biomass by not subtracting root volume from soil volume. As forests typically have high root biomass, forest bulk densities were consistently underestimated. We therefore considered both the concentration and content data in our analyses. At the two Tsitsikamma sites, we dried and weighed the roots before the samples were sent for analysis. Litter was also collected from beneath the canopy of the three treatments (forest, fynbos and V. divaricata) and both root and litter samples were analysed for nitrogen and carbon using a Leco TruSpec CN Analyser (LECO Corporation, St. Joseph, USA).
Statistical analyses
All statistical analyses in this study were performed using R (R Development Core Team 2011). Principle component analyses (PCA) were used to explore the relationships between the three vegetation types and various elemental soil variables. The broken-stick distribution (PCA signifance criterion in R) was used as the most reliable criterion to test which of the PCA axes contributed significantly to the ordination. PCA axes with larger percentages of accumulated variance than the broken-stick variance were considered significant (Kindt & Coe 2005; Legendre & Legendre 1998). A three-way analysis of variance (ANOVA) was fitted to explore the effects of site, vegetation type, depth and the respective interactions on soil factors. Where necessary, variables were transformed to attain approximate normality and homogeneity of variance. Post hoc Tukey tests were used to test for differences between levels of each factor in the ANOVAs.
Results
PC1 of the PCA explained 59% of the variance in soils (eigenvalue 4.19; x-axis in Figure 1), with high loads for nitrogen (-0.39), carbon (-0.39), potassium (-0.41), magnesium, sodium and phosphorus (all -0.38). The three vegetation types were arranged along a gradient of soil fertility, according to which forests were found on the most fertile soils, V. divaricata was associated with intermediate soil nutrients, and fynbos was found on low-nutrient soils. PC2 (y-axis in Figure 1) explained a further 16% of the variance and separated sites with high nitrogen (+0.42) and carbon (+0.23) from those with high calcium (-0.71) and magnesium (-0.44); however, this axis was not deemed significant. This axis reflected site differences to a great extent. Covie soils, for example, had very high levels of calcium and magnesium.
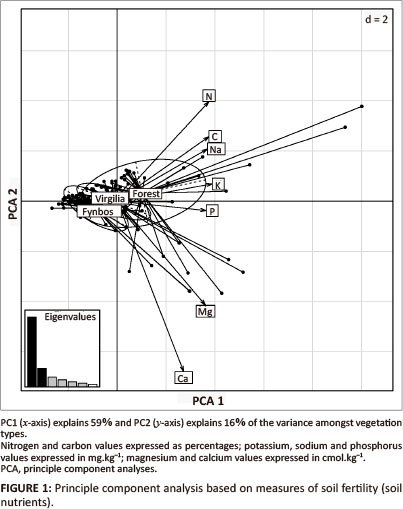
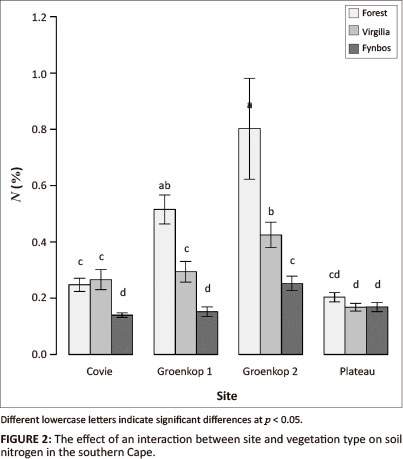
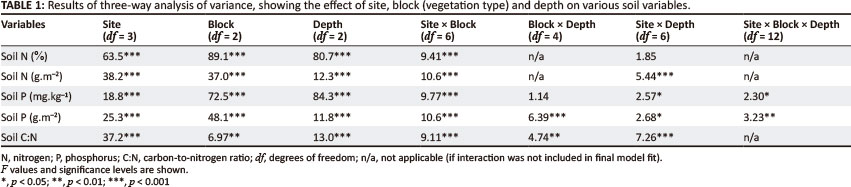
ANOVA results showed that the interactive effect of site and vegetation type on nitrogen concentration was statistically significant but not the interaction amongst the three factors or any of the other two-way interactions (Table 1). Although depth had less of an effect on nitrogen content, it was an important determinant of soil concentrations (Table 1). When nitrogen concentration was converted to nitrogen content, both the interactions between site and vegetation type and site and depth became important in determining nitrogen (Table 1).
Across sites, forest soils had the highest nitrogen concentration (0.44% ± 0.05% and 34.2 g.m-2 ± 3.18 g.m-2), followed by V. divaricata soils (0.29% ± 0.02% and 25.4 g.m-2 ± 1.55 g.m-2) and fynbos soils (0.18% ± 0.01% and 19.2 g.m-2 ± 0.86 g.m-2). For individual sites, Groenkop 1 and Groenkop 2 reflected the general pattern for both nitrogen concentration (Figure 1a) and nitrogen content (Table 2), with the highest values associated with forest and the lowest values associated with fynbos. At Covie, forest and V. divaricata soils had similar nitrogen concentrations (0.24% ± 0.02% and 0.26% ± 0.04%, respectively), but these were higher than for fynbos (0.14% ± 0.01%; Figure 1a). Nitrogen content was similar for forest and V. divaricata, but V. divaricata had higher nitrogen levels than fynbos (Table 2). At Plateau, all three vegetation types had similar nitrogen concentrations (Figure 2) and nitrogen content (Table 2). As expected, the shallowest soils generally had the highest nitrogen concentration (0.44% ± 0.56%), followed by the 10-cm - 20-cm depths (0.19% ± 0.01%) and then the deeper soils (0.27% ± 0.19%).

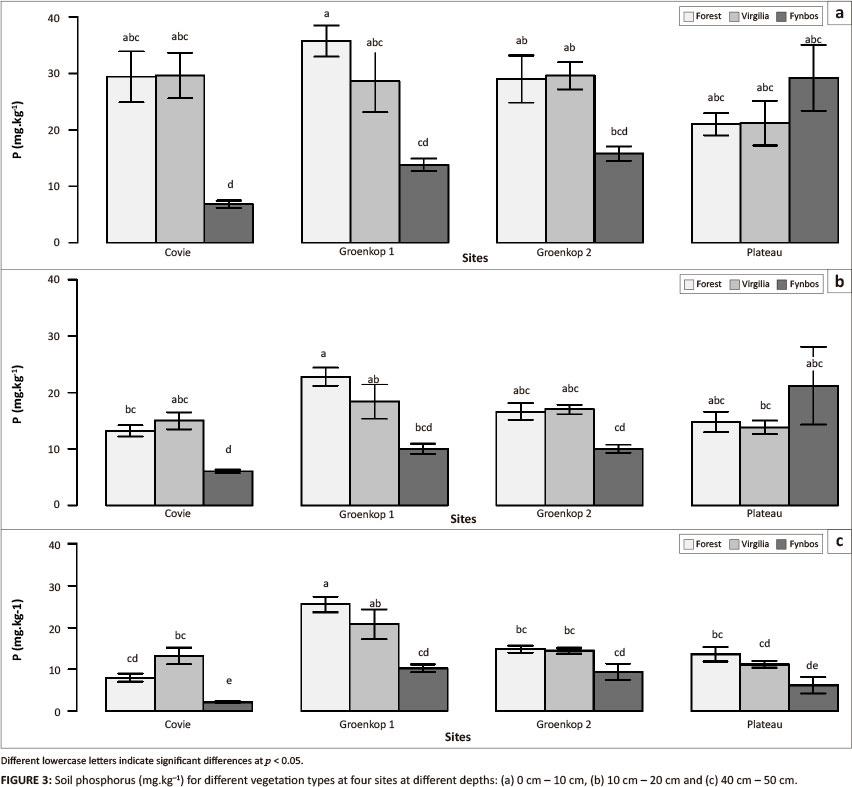
Significant interactions affecting phosphorus concentration are shown in Table 1. When phosphorus concentrations were converted to phosphorus content, the interaction between vegetation type and depth, in addition to the other factors and interactions, was also statistically significant (Table 1). Although the three-way interaction was significant, its importance was relatively small compared to site χ vegetation type and vegetation type χ depth (Table 1). However, to confirm that the three-way interaction was significant, we compared the interaction between site and vegetation type for each depth (Figure 3). At 0 cm - 10 cm and 10 cm - 20 cm, Covie forest and V. divaricata soils had higher phosphorus concentrations than fynbos; Groenkop 1 had higher phosphorus concentrations associated with forest than fynbos (Figure 3). No differences were found for phosphorus concentration between vegetation types at Plateau. At 40 cm - 50 cm, fynbos still had a lower phosphorus concentration than forest and V. divaricata at Covie, but phosphorus concentration for fynbos was lower than for both forest and V. divaricata at Groenkop 1 and Plateau (Figure 3). When phosphorus concentrations were converted to content data, no differences were found between vegetation types at Groenkop 2. Virgilia divaricata had higher phosphorus content than fynbos at Covie and Groenkop 1, whilst fynbos soils had higher phosphorus content than the other two vegetation types at Plateau (Table 2).
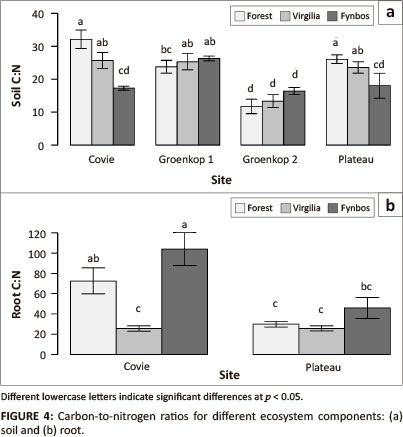
We measured soil carbon-to-nitrogen ratios (C:N) at all four sites, as well as C:N for litter and root material at Covie and Plateau. Site had a very pronounced influence on soil C:N and all interactions with site were significant (Table 1). Apart from the interactions with site, the interaction between vegetation type and depth was also statistically significant. The three-way interaction was not significant. When comparing individual sites, soils from Groenkop 1 and Groenkop 2 showed no differences in C:N between the three vegetation types (Figure 4). At Covie, soils beneath forest and V. divaricata showed the highest C:N, both of which were higher than for soils beneath fynbos (Figure 4). At Plateau, forest and V. divaricata had similar values, and V. divaricata and fynbos had similar values, but forest had higher soil C:N than fynbos. The site χ vegetation type interaction was statistically significant for root C:N (p = 0.002). At Covie, V. divaricata soils had lower root C:N than fynbos soils, but no differences were found at Plateau (Figure 4). Unfortunately, we did not have enough repetitions to compare C:N for litter statistically and can therefore comment only on the general trend. Litter C:N showed the same pattern as root C:N, with V. divaricata samples having the lowest litter C:N (22.8 ± 1.75), followed by forest (35.7 ± 3.16) and fynbos samples (51.8 ± 6.41).
Discussion
In this study, we compared differences in soil fertility between stands dominated by forest, V. divaricata and fynbos vegetation, respectively, to establish the effect of V. divaricata on soil fertility. Firstly, we used PCA to explore relationships amongst soil variables and found that the different vegetation types separated in ordinational space, with forest, V. divaricata and fynbos on a gradient of high to low soil nutrients and carbon. We also compared in more detail the differences in nitrogen and phosphorus amongst the three vegetation types. Although the results were site depended, both nitrogen and phosphorus concentrations appeared to be low in fynbos soils, with higher values found in forest and V. divaricata soils. It was only at Plateau where phosphorus concentration was higher in fynbos soils compared with the other vegetation types. The lower nutrient status of fynbos soils is not unexpected, as previous work has shown that fynbos soils are nutrient poor, with low concentrations and availability of nitrogen and phosphorus (Richards, Stock & Cowling 1997; Stock & Lewis 1986; Stock, Wienand & Baker 1995; Witkowski & Mitchell 1987). Fynbos vegetation has several adaptations to low nutrient availability, including cluster (e.g. proteoid, capillaroid and dauciform) roots and mycorrhizal associations that confer efficient nutrient uptake (Allsopp & Stock 1993; Lamont 1982) and high nutrient-use efficiency (Stock & Lewis 1984; Van Daalen 1981). The higher phosphorus concentration and content of fynbos soils at Plateau was an unexpected result. However, the fynbos area at this site was farmed until recently and we cannot rule out that phosphorus-rich fertilisers were used at the site. As phosphorus is bound quite strongly by soils, it is not easily leached from soils. This could explain the higher phosphorus values found for the top 20 cm, despite the lower phosphorus values in deeper soils compared with the other vegetation types.
The establishment of forest on fynbos soils is not an uncommon phenomenon and forest invasion into fynbos during prolonged fire-free periods has been reported for many sites (Geldenhuys 2004; Kruger 1984; Manders 1990). Although fynbos soils are generally low in nitrogen (Stock & Lewis 1986) and phosphorous (Witkowski & Mitchell 1987), Van Daalen (1984) has shown that in the southern Cape forest can establish on most soils. Fynbos and forest soils seldom appeared to differ and therefore Van Daalen (1984) described any existing differences as an artefact of the vegetation. Van Daalen (1981) rejected the hypothesis that the poorer nutrient status of fynbos soils precludes the colonisation of fynbos by forest species. Bond (2010) supported this when he showed that most fynbos soils have enough micronutrients to build forest when deeper soil layers are included. However, Van Daalen (1981) found no regeneration of forest in fynbos at some sites in our study area and attributed this to the dependence of forest species on the 'closed' nutrient cycling of the forest ecosystem, where nutrients are tightly and efficiently cycled within the root zone.
Increasing research findings point to vegetation feedbacks having significant effects on soil fertility to explain establishment of trees on nutrient-poor fynbos soils (Vitousek 1990; Vitousek & Walker 1989; Wedin & Tilman 1990). For example, invasive Australian Acacia species invade fynbos over vast areas in South Africa, with profound changes in soil as a result (Higgins et al. 1999; Van Wilgen et al. 2001). These alien nitrogen-fixing Acacia species regenerate rapidly after fire (Milton & Hall 1981) and the subsequent large inputs of high-nitrogen leaf litter elevate total soil nitrogen and organic matter (Stock et al. 1995; Witkowski 1991a; Yelenik et al. 2004). Alien acacias have been found to have higher leaf nitrogen concentrations than indigenous vegetation and no differences in resorption were found between fynbos species and alien acacias during a study in another fynbos area, which meant that more nitrogen was returned to soil in the acacia litter (Witkowski 1991b). A combination of high tolerance to low shade conditions and fast modification of the nutrient cycle thus results in successful invasion of alien acacias into fynbos.
In contrast with alien acacias, forest trees typical of afrotemperate forests do not tolerate regeneration in sunny micro-climates well and are, as a rule, quite sclerophyllous (Van Daalen 1981). This leads to low decomposition rates (Witkowski 1991b). Van Daalen (1984) suggests that for forest to re-establish in a fynbos habitat or after disturbance, forest precursor species that can tolerate low nutrient levels and summer water deficits establish first. Decomposition and nutrient return were found to be higher in a study comparing strandveld (sclerophyllous vegetation with a higher tree component on the west coast of South Africa) with fynbos (Witkowski 1991b). Although nitrogen-fixing trees increase soil nitrogen over time owing to nitrogen-enriched litter being returned to the soil, soil nutrients may also be enhanced in other ways. Trees may increase soil nutrients by:
• drawing nutrients from deep soil layers and areas beyond the canopy and depositing them in the form of root exude, litter and/or canopy leaching (Belsky 1994; Jobbágy & Jackson 2004; Kellman 1979; Scholes & Hall 1996)
• acting as an atmospheric dust trap and increasing nutrients from canopy leaching (Bernhard-Reversat 1988; Escudero et al. 1985)
• attracting mammals and roosting birds by providing shade and refuge, who then add nutrients by means of their droppings (Belsky 1994; Giorgiadis 1989).
When forest precursor trees establish in the current study area, litter builds up over time and eventually a dense mat of decomposing root material and organic material is formed (Geldenhuys 2011; Geldenhuys & Theron 1994). Shallow feeder roots depend on the nutrients released from decomposing material and take up these nutrients before they are leached; in this way nutrients are increased and cycled quite tightly within the rooting zone. Small additions of nitrogen to nutrient-poor fynbos soils resulted in significant changes in the growth-form composition of the vegetation and led to increases in graminoid forms of fynbos elsewhere (Witkowski 1991b). Some fynbos species show decreased growth rates with increased levels of nitrogen or phosphorus in the soil (Witkowski 1989). Some restios have been shown to decrease allocation to reproduction when treated with nitrogen- and phosphorus-rich fertilisers (Witkowski 1990). Apart from decreasing the competitiveness of fynbos vegetation and restoring the nutrient cycle, forest precursor species provide suitable shade, with accompanying higher dispersal opportunities and ameliorated soil water conditions. Once these conditions re-establish, forest trees reestablish (Geldenhuys 2004).
If nitrogen-fixing V. divaricata enriches soils in a similar manner to the invasive acacias, then the nutrient cycle should be re-established much faster than under sclerophyllous forest precursors. Invasive acacias show faster decomposition rates and nitrogen return compared with the local forest precursor tree Pterocelastrus tricuspidatus as a result of nitrogen-rich litter (Witkowski 1991b). Similarly, we found that V. divaricata litter (and roots) tends to have higher nitrogen concentrations than either forest or fynbos litter (with accompanying low C:N). In some ways V. divaricata has a very similar ecology to that of Australian acacias in that it recruits rapidly after fire and has the potential to fix nitrogen. However, some major differences between invasive acacias and V. divaricata are that V. divaricata is not as persistent and long lived as the acacias. This allows other forest species to establish over time, as Virgilia spp. stands usually die off naturally after approximately 20 years owing to caterpillars of the Leto venus moth ringbarking trees (C. Geldenhuys, pers. comm., January 2011). Virgilia divaricata stands are typically also less dense than invasive acacia stands, thereby allowing gaps for indigenous shade-tolerant forest species to establish.
Conclusion
We compared soil fertility between three co-occurring vegetation types: V. divaricata-dominated stands with forest and fynbos on either side. Our findings suggested that V. divaricata enhances soil fertility. However, owing to the nature of our work, the link between V. divaricata and higher soil nutrients is only tentative and further experimental work should be undertaken to confirm that V. divaricata enhances soil nutrients. Nonetheless, owing to its ability to grow fast and establish without shade, we suggest that V. divaricata can be gainfully used to restore the 'closed' nutrient cycle found beneath forest and in this way aid in efficient forest regeneration. In contrast, maintaining 'healthy' fynbos (i.e. with a competitive advantage over trees) requires regular fires, both to maintain the ideal micro-climate for fynbos vegetation and to disrupt the nutrient cycle associated with tree-dominated vegetation.
Acknowledgements
The authors want to thank the following people and organisations: BTech students Oiva Akudhenga, Iikela Sioni, Muzuma Uakendisa, Don de Swart, Andrew Jackson and Roxane Moulet for performing fieldwork, SANParks for allowing us to work in the Garden Route National Park, the Haupt family for allowing us to work at Plateau and Mike Cameron for comments on an earlier draft of the manuscript.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
C.C. (Nelson Mandela Metropolitan University) and B.J.W. (Nelson Mandela Metropolitan University) designed the research questions and organised fieldwork. C.C. wrote the text and B.J.W. assisted in creating artwork.
References
Allsopp, N. & Stock, W.D., 1993, 'Mycorrhizal status of plants growing in the Cape Floristic Region, South Africa', Bothalia 23, 91-104. [ Links ]
Barnes, P.W. & Archer, S., 1999, 'Tree-shrub interactions in a subtropical savanna parkland: Competition or facilitation?', Journal of Vegetation Science 10, 523-536. http://dx.doi.org/10.2307/3237187 [ Links ]
Belsky, A.J., 1994, 'Influences of trees on savanna productivity: Tests of shade, nutrients, and tree-grass competition', Ecology 75, 922-934. http://dx.doi.org/10.2307/1939416 [ Links ]
Bernhard-Reversat, F., 1988, 'Soil nitrogen mineralization under a Eucalyptus plantation and a natural Acacia forest in Senegal', Forest Ecological Management 23, 233-244. http://dx.doi.org/10.1016/0378-1127(88)90054-0 [ Links ]
Bond, W.J., 1981, 'Vegetation gradients in the southern Cape mountains', MSc thesis, University of Cape Town. [ Links ]
Bond, W.J., 2010, 'Do nutrient-poor soils inhibit development of forests? A nutrient stock analysis', Plant and Soil 334, 47-60. http://dx.doi.org/10.1007/s11104-010-0440-0 [ Links ]
Campbell, B.M. & Killick, D.J.B., 1985, A classification of the mountain vegetation of the fynbos biome, Memoirs of the Botanical Survey of South Africa, vol. 50, Botanical Research Institute, Department of Agriculture, South Africa. [ Links ]
Chapman, J.L. & Reiss, M.J., 1992, Ecology principles and applications, Cambridge University Press, Cambridge, UK. [ Links ]
Crain, C.M. & Bertness, M.D., 2006, 'Ecosystem engineering across environmental gradients: Implications for conservation and management', Bioscience 56, 211218. http://dx.doi.org/10.1641/0006-3568(2006)056[0211:EEAEGI]2.0.CO;2 [ Links ]
Escudero, A., Garcia, B., Gomex, J.M. & Luis, E., 1985, 'The nutrient cycling in Quercus rotundifolia and Q. pyrenaica ecosystems ('dehesas') of Spain', Acta Oecologica. Oecological Plantarum 6, 73-86. [ Links ]
Geldenhuys, C.J., 1982, 'The management of the Southern Cape forests', paper presented at the Eighth regular meeting, SARCCUS standing committee for forestry, Louis Trichardt, 09-13th October 1981. [ Links ]
Geldenhuys, C.J., 1991, 'Distribution, size and ownership of forests in the southern Cape', South African Forestry Journal 158, 51-68. http://dx.doi.org/10.1080/00382167.1991.9630375 [ Links ]
Geldenhuys, C.J., 1994, 'Bergwind fires and location pattern of forest patches in the Southern Cape landscape, South Africa', Journal of Biogeography 21, 49-62. http://dx.doi.org/10.2307/2845603 [ Links ]
Geldenhuys, C.J., 2004, 'Concepts and processes to control invader plants in and around natural evergreen forest in South Africa', Weed Technology 18, 13861391. http://dx.doi.org/10.1614/0890-037X(2004)018[1386:CAPTCI]2.0.CO;2 [ Links ]
Geldenhuys, C.J., 2011, 'How to live with trees and keep them healthy', South African Forestry Magazine, June, pp. 36-37. [ Links ]
Geldenhuys, C.J. & Theron, J.M., 1994, 'Litterfall and nutrient cycling in mixed evergreen forest near George, South Africa', report FOR-DEA 746, Division of Forest Science and Technology, CSIR, Pretoria. [ Links ]
Giorgiadis, N.J., 1989, 'Microhabitat variation in an African savanna: Effect of woody cover and herbivores in Kenya', Journal of Tropical Ecology 5, 93-108. http://dx.doi.org/10.1017/S0266467400003254 [ Links ]
Higgins, S.I., Richardson, D.M., Cowling, R.M. & Trinder-Smith, T.H., 1999, 'Predicting the landscape-scale distribution of alien plants and their threat to plant diversity', Conservation Biology 13, 303-313. http://dx.doi.org/10.1046/j.1523-1739.1999.013002303.x [ Links ]
Jobbágy, E.G. & Jackson, R.B., 2004, 'The uplift of soil nutrients by plants: Biogeochemical consequences across scales', Ecology 85, 2380-2389. http://dx.doi.org/10.1890/03-0245 [ Links ]
Kellman, M., 1979, 'Soil enrichment of Neotropical savanna trees', Journal of Ecology 67, 565-577. http://dx.doi.org/10.2307/2259112 [ Links ]
Kindt, R. & Coe, R., 2005, Tree diversity analysis: A manual and software for common statistical methods for ecological and biodiversity studies, World Agroforestry Centre (ICRAF), Nairobi. [ Links ]
Kruger, F.J., 1984, 'Effects of fire on vegetation structure and dynamics', in P. de V. Booysen & N.M. Tainton (eds.), Ecological effects of fire in South African ecosystems, pp. 219-244, Springer Verlag, Berlin. http://dx.doi.org/10.1007/978-3-642-69805-7_10 [ Links ]
Laland, K.N., Odling-Smee, F.J. & Feldman, M.W., 1999, 'Evolutionary consequences of niche construction and their implications for ecology', Proceedings of the National Academy of Sciences of the United States of America 96, 10242-10247. http://dx.doi.org/10.1073/pnas.96.18.10242 [ Links ]
Lamont, B., 1982, 'Mechanisms for enhancing nutrient uptake in plants, with particular reference to Mediterranean South Africa and Western Australia', The Botanical Review 48, 597-689. http://dx.doi.org/10.1007/BF02860714 [ Links ]
Legendre, P. & Legendre, L., 1998, Numerical ecology, 2nd edn., Elsevier, Amsterdam. [ Links ]
Manders, P.T., 1990, 'Fire and other variables as determinants of forest/fynbos boundaries in the Cape Province', Journal of Vegetation Science 1, 483-490. http://dx.doi.org/10.2307/3235782 [ Links ]
Manders, P.T. & Richardson, D.M., 1992, 'Colonization of Cape fynbos communities by forest species', Forest Ecology and Management 48, 277-293. http://dx.doi.org/10.1016/0378-1127(92)90150-8 [ Links ]
Manders, P.T., Richardson, D.M. & Masson, P.H., 1992, 'Is fynbos a stage in succession to forest? Analysis of the perceived ecological distinction between two communities', in B.W. van Wilgen, D.M. Richardson, F.J. Kruger & H.J. van Hensbergen (eds.), Fire in South African Mountain Fynbos: Ecosystem, community and species response at Swartboskloof, pp. 81-107, Springer Verlag, Berlin. [Ecological Studies vol. 93]. http://dx.doi.org/10.1007/978-3-642-76174-4_5 [ Links ]
Masson, P.H. & Moll, E.J., 1987, 'The factors affecting forest colonization of fynbos in the absence of recurrent fire at Orange Kloof, Cape Province, South Africa', South African Journal of Forestry 143, 5-10. http://dx.doi.org/10.1080/00382167.1987.9630294 [ Links ]
Milton, S.J. & Hall, A.V., 1981, 'Reproductive biology of Australian Acacias in the South-Western Cape province, South Africa', Transactions of the Royal Society of South Africa 44, 465-487. http://dx.doi.org/10.1080/00359198109520589 [ Links ]
Mucina, L. & Rutherford, M.C., 2006, The vegetation of South Africa, Lesotho and Swaziland, South African National Biodiversity Institute, Pretoria. [Strelitzia no. 19]. [ Links ]
Musil, C.F., 1993, 'Effect of invasive Australian Acacias on the regeneration, growth and nutrient chemistry of South African Lowland fynbos', Journal of Applied Ecology 30, 361-372. http://dx.doi.org/10.2307/2404637 [ Links ]
Nelson, D.W. & Sommers, L.E., 1996, 'Total carbon, organic carbon, and organic matter', in D.L. Sparks, A.L. Page, P.A. Helmke, R.H. Loeppert, P.N. Soluanpour, M.A. Tabatabai, C.T. Johnston & M.E. Sumner (eds.), Methods of soil analysis part 3: Chemical methods, pp. 961-1010, Soil Science Society of America, Inc. and American Society of Agronomy, Inc., Madison, Wisconsin, USA. [ Links ]
Odling-Smee, J., Laland, K.N. & Feldman, M.W., 1996, 'Niche construction', The American Naturalist 147, 641-648. http://dx.doi.org/10.1086/285870 [ Links ]
Palgrave, K.C., 1981, Trees of Southern Africa, Struik, Cape Town. [ Links ]
Phillips, J.F.V., 1926, 'Virgilia divaricata capensis Lam. ('Keurboom'): A contribution to its ecology and sylviculture', South African Journal of Science 23, 435-454. [ Links ]
R Development Core Team, 2011, R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna. [ Links ]
Richards, M.B., Stock, W.D. & Cowling, R.M., 1997, 'Soil nutrient dynamics and community boundaries in the fynbos vegetation of South Africa', Plant Ecology 130, 143-153. http://dx.doi.org/10.1023/A:1009742225383 [ Links ]
Scholes, R.J. & Hall, D.O., 1996, 'The carbon budget of tropical savannas, woodlands, and grasslands', in A. Breymeyer, D. Hall, J. Melillo & G. Agren (eds.), Modelling Terrestrial Ecosystems, pp. 69-100, Wiley, Chichester. [ Links ]
Stock, W.D. & Lewis, O.A.M., 1984, 'Uptake and assimilation of nitrate ammonium by an evergreen fynbos shrub species Protea repens L. (Proteaceae)', New Phytologist 97, 261-268. http://dx.doi.org/10.1111/j.1469-8137.1984.tb04130.x [ Links ]
Stock, W.D. & Lewis, O.A.M., 1986, 'Soil nitrogen and the role of fire as a mineralizing agent in a South African coastal fynbos ecosystem', Journal of Ecology 74, 317328. http://dx.doi.org/10.2307/2260257 [ Links ]
Stock, W.D., Wienand, K.T. & Baker, A.C., 1995, 'Impacts of invading N2-fixing Acacia species on patterns of nutrient cycling in two Cape ecosystems: Evidence from soil incubation studies and 15N natural abundances', Oecologia 101, 375-382. http://dx.doi.org/10.1007/BF00328825 [ Links ]
Van Daalen, J.C., 1981, 'The dynamics of the indigenous forest-fynbos ecotone in the southern Cape', South African Forestry Journal 119, 14-23. http://dx.doi.org/10.1080/00382167.1981.9630218 [ Links ]
Van Daalen, J.C., 1984, 'Distinguishing features of forest species on nutrient-poor soils of the southern Cape', Bothalia 1/2, 229-239. [ Links ]
Van der Merwe, I., 2011, Green heritage: Trees, forests and woodlands of South Africa, Department of Agriculture, Forestry and Fisheries, Pretoria. [ Links ]
Van Wilgen, B.W., Richardson, D.M., Le Maitre, D.C., Marais, C. & Magadlela, D., 2001, 'The economic consequences of alien plant invasions: Examples of impacts and approaches to sustainable management in South Africa', Environment, Development and Sustainability 3, 145-168. http://dx.doi.org/10.1023/A:1011668417953 [ Links ]
Walkley, A., 1947, 'A critical examination of a rapid method for determining organic carbon in soils: Effect of variations in digestion conditions and of inorganic soil constituents', Soil Science 63, 251-263. http://dx.doi.org/10.1097/00010694-194704000-00001 [ Links ]
Vitousek, P.M., 1990, 'Biological invasions and ecosystem processes: Towards an integration of population biology and ecosystem studies', Oikos 57, 7-13. http://dx.doi.org/10.2307/3565731 [ Links ]
Vitousek, P.M. & Walker, L.W., 1989, 'Biological invasions by Myrica faya in Hawai'i: Plant demography, nitrogen fixation, ecosystem effects', Ecological Monographs 59, 247-265. http://dx.doi.org/10.2307/1942601 [ Links ]
Wedin, D.A. & Tilman, D., 1990, 'Species effects on nitrogen cycling: A test with perennial grasses', Oecologia 84, 433-441. [ Links ]
White, F., 1978, 'The Afromontane Region', in M.J.A. Werger (ed.), Biogeography and Ecology of Southern Africa, pp. 463-513, Junk, The Hague. http://dx.doi.org/10.1007/978-94-009-9951-0_11 [ Links ]
Witkowski, E.T.F., 1989, 'Effects of nutrients on the distribution of dry mass, nitrogen and phosphorus in seedlings of Protea repens (L.) L. (Proteaceae)', New Phytologist 112, 481-487. http://dx.doi.org/10.1111/j.1469-8137.1989.tb00341.x [ Links ]
Witkowski, E.T.F., 1990, 'Effect of nutrient additions on the allocations of dry mass, phosphorus and nitrogen in the cape fynbos plant, Thamnochortus punctatus Pill. (Restionaceae)', South African Journal of Ecology 1, 38-44. [ Links ]
Witkowski, E.T.F., 1991a, 'Effects of invasive alien acacias on nutrient cycling in the coastal lowlands of the Cape fynbos', Journal of Applied Ecology 28, 1-15. http://dx.doi.org/10.2307/2404109 [ Links ]
Witkowski, E.T.F., 1991b, 'Growth and competition between seedlings of Protea repens (L.) L. and alien invasive, Acacia saligna (Labill.) Wendl., in relation to nutrient availability', Functional Ecology 4, 101-110. http://dx.doi.org/10.2307/2389560 [ Links ]
Witkowski, E.T.F. & Mitchell, D.T., 1987, 'Variations in soil phosphorus in the fynbos biome, South Africa', Journal of Ecology 75, 1159-1171. http://dx.doi.org/10.2307/2260320 [ Links ]
Yarranton, G.A. & Morrison, R.G., 1974, 'Spatial dynamics of a primary succession: Nucleation', Journal of Ecology 62, 417-428. http://dx.doi.org/10.2307/2258988 [ Links ]
Yelenik, S.G., Stock, W.D. & Richardson, D.M., 2004, 'Ecosystem level impacts of invasive Acacia saligna in the South African fynbos', Restoration Ecology 12, 4451. http://dx.doi.org/10.1111/j.1061-2971.2004.00289.x [ Links ]
 Correspondence:
Correspondence:
Corli Coetsee
Private Bag X6531
George 6530, South Africa
corli.coetsee@nmmu.ac.za
Received: 26 Nov. 2012
Accepted: 22 Apr. 2013
Published: 15 July 2013


{kind=link}
{kind=link}