










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkKoedoe
versión On-line ISSN 2071-0771
versión impresa ISSN 0075-6458
Koedoe vol.55 no.1 Pretoria ene. 2013
Ecology of the plant-dwelling spiders (Arachnida: Araneae) of the Erfenis Dam Nature Reserve, South Africa
René FourieI; Charles R. HaddadI; Anna S. Dippenaar-SchoemanII, III; Anel GroblerI
IDepartment of Zoology and Entomology, University of the Free State, South Africa
IIAgricultural Research Council - Plant Protection Research Institute, Queenswood, South Africa
IIIDepartment of Zoology and Entomology, University of Pretoria, South Africa
ABSTRACT
As part of the South African National Survey of Arachnida in the Grassland Biome, foliage-dwelling and grass-dwelling spiders (Arachnida: Araneae) were collected in the Erfenis Dam Nature Reserve in the central Free State Province from November 2005 to August 2007. Foliage-dwelling spiders were collected from three common tree or shrub species (Acacia karroo, Searsia ciliata and Searsia lancea) and grass-dwellers from four contrasting grasslands (uniform Themeda triandra, mixed, weedy and woodland grasslands). From the grass layer, 1649 spiders were collected, representing 15 families and 82 species, whilst 496 tree-dwelling spiders were collected that represented 17 families and 52 species. There was some overlap in the fauna of the two strata, resulting in a total of 108 species from 18 families being collected. The Araneidae, Philodromidae, Salticidae and Thomisidae were consistently the most abundant in all grassland types and tree species, although Salticidae were scarce on A. karroo. Assemblage analysis indicates high similarity and overlap in the fauna of the four grassland types, suggesting that the structural complexity of grasslands has a limited effect on species composition. In contrast, the foliage-dwelling assemblages were more distinct, with only some overlap between the faunas of S. ciliata and A. karroo, suggesting a stronger vegetation structural effect in shaping arboreal spider assemblages. The isolation of trees and shrubs within the extensive grassy habitat may contribute to the more unique fauna and lower species richness of the woody vegetation.
Conservation Implications: This study uncovered a rich diversity of plant-dwelling spiders from central South Africa. Grassland faunas show considerable temporal variation and some variability in microhabitat preferences, and sampling protocols should take this into account when spiders are considered in management plans and biodiversity surveys in reserves and private land.
Introduction
True grasslands were historically quite widespread throughout the southern parts of the African continent (Bredenkamp, Spada & Kazmierczak 2002), but, at present, the Grassland Biome is largely limited to the central plateau of South Africa, Lesotho and parts of Swaziland (Carbutt et al. 2011; Mucina & Rutherford 2006; Palmer & Ainslie 2005). The Grassland Biome covers approximately a quarter of South Africa's land surface (Palmer & Ainslie 2005) and is considered to have an extremely high plant biodiversity, second only to that of the Fynbos Biome (Low & Rebelo 1996). A large proportion (approximately 40%) of the biome has been irreversibly transformed through agriculture, mining, urbanisation and industrialisation (Mentis & Huntley 1982; O'Connor & Kuyler 2005) and, as such, the remaining grassland habitat is of great conservation value and can be considered highly irreplaceable (Egoh 2009). Only an estimated 2.04% - 2.80% of the grassland area is conserved (Carbutt et al. 2011; O'Connor & Kuyler 2005) and therefore effective management and conservation of private land is critical to protect the highly endemic fauna and flora of this biome (Wessels et al. 2003). Grasslands can be defined on the basis of a single-layered herbaceous plant community dominated by tufted graminoids (especially perennial Poaceae), with perennial forbs that appear annually, and the rarity of woody plants, which are usually restricted to specific habitats that act as fire refugia, including drainage lines and rocky hilltops (Carbutt et al. 2011).
Regarding spider biodiversity, data in the National Collection of Arachnida database at the Agricultural Research Council - Plant Protection Research Institute, Pretoria, indicates that nearly 910 point localities have been sampled in the Grassland Biome, but only 27 of these have more than 100 specimen records (Foord, Dippenaar-Schoeman & Haddad 2011). This shows that large parts of the biome remain very poorly studied. Only as recently as three decades ago were the first ecological studies on spiders undertaken in the Grassland Biome, focusing on the ground-dwelling (Jansen et al. in press; Lotz, Seaman & Kok 1991; Van den Berg & Dippenaar-Schoeman 1991), termitophilous (Haddad & Dippenaar-Schoeman 2002, 2006), grass- dwelling (Dippenaar-Schoeman, Hamer & Haddad 2011; Haddad 2005) and litter-dwelling assemblages (Butler & Haddad 2011). Consequently, there is considerable scope for research on spider biodiversity and ecological patterns in this unique vegetation type.
A project was initiated in September 2005 to determine the arachnofauna of the Erfenis Dam Nature Reserve, located near Theunissen in the Free State Province of South Africa. Initially, the primary focus of the study was to determine the impact of controlled burning on various faunal groups, which included arachnids, insects and small mammals. Whilst the project was focused on the ground-dwelling fauna, the potential was later recognised to expand the project to the sampling of foliage-dwelling and grass-dwelling spiders from unburned areas as well. Thus, the aim of this study was to determine the spider communities in four contrasting grassland types and three common tree species in the reserve over a 2-year period. This study also forms part of the South African National Survey of Arachnida (SANSA) in the Grassland Biome.
Materials and methods
Study area and period
The Erfenis Dam Nature Reserve is located in the central part of the Free State Province, about 18 km south-east of Theunissen. The dam that shares the same name as the reserve is part of the Vet River system and extends over 3800 ha when full, with the reserve comprising about 400 ha of grassland surrounding the dam (Stuart & Stuart 1997). Sampling was carried out during the last week of every third month from November 2005 until August 2007, coinciding with the end of each of the four seasons of the year.
Grass-dwelling spiders were collected from four contrasting types of grasslands (abbreviations and generalised coordinates given in parenthesis), which can be described as follows (plant classification follows Germishuizen et al. 2006). Uniform Themeda triandra Forssk. grassland (TG; 28°29.8'S, 26°47.5'E) that consisted primarily of homogenous T. triandra, with small scattered patches of Eragrostis and Cymbopogon grasses. Mixed grassland (MG; 28°29.9'S, 26°48.5'E), which consisted of various Eragrostis species, Aristida congesta Roem. & Schult., Enneapogon cenchroides (Roem. & Schult.) C.E.Hubb, Setaria pumila (Poir.) Roem. & Schult., Sporobolus fimbriatus (Trin.) Nees and Tragus berteronianus Schult., with very little T. triandra present. Weedy grassland (WeG; 28°29.7'S, 26°48.4'E) was structurally the most heterogenous, consisting of A. congesta, Cymbopogon pospischilii (K.Shumm.) C.E.Hubb., Eragrostis lehmanniana Nees, E. tef (Zucc.) Trotter, Setaria pumila and Sporobolus fimbriatus grasses, with a variety of weeds, including Nidorella resedifolia DC., and Berkeya spp. Lastly, woodland grassland (WoG; 28°30.2'S, 26°47.5'E), which was dominated by T. triandra and Eragrostis grasses, interspersed with Acacia karroo Hayne (Mimosaceae), Searsia lancea Barkley (Anacardiaceae) and Ziziphus mucronata Willd. (Rhamnaceae) trees.
Foliage-dwelling spiders associated with woody plants were collected from three different tree or shrub species: the trees A. karroo (AK; 28°30.2'S, 26°47.5'E) and S. lancea (SL; 28°29.6'S, 26°47.6'E) and the low-growing shrub Searsia ciliata Licht. ex Schult (SC; 28°29.9'S, 26°48.5'E). Acacia karroo is widespread throughout South Africa, except for the Fynbos and Succulent Karoo Biomes, and is found in a variety of habitats, from low-lying areas to the Highveld. This tree species usually has a rounded crown, branching fairly low down on the trunk. It is variable in shape and size, reaching a maximum height of about 12 m where there is good water nearby. The leaves are finely textured and dark green, with flowers that appear in early summer in a mass of yellow pompoms. The thorns are usually paired, with a greyish to white colour and are long and straight (Van Wyk & Van Wyk 1997; Venter & Joubert 1984). Searsia lancea is a small to medium sized evergreen tree that usually grows to a height of 7 m and a width of 7 m and is found throughout the Free State. It is usually a single-stemmed, low branching tree that has a dense, soft, round canopy with compound leaves, possessing three narrowly lanceolate (lance shaped) leaflets. The small, inconspicuous flowers are greenish-yellow in colour and are produced from June until September (i.e. winter to early spring). Searsia lancea occurs naturally in Acacia woodland and along drainage lines, rivers and streams (Van Wyk & Van Wyk 1997; Venter & Joubert 1984). Searsia ciliata forms an evergreen, thorny, thin and crooked-stemmed shrub that is 2 m - 4 m tall. The leaves are also trifoliolate, with minute flowers that are yellow-green in colour and insignificant. The fruits are drupes and are shiny light to dark brown in colour. It is widespread in the central drier parts of South Africa (Venter & Joubert 1984).
Sampling methods
Grass-dwelling spiders were collected using a standard sweep net with a diameter of 35 cm. A total of 250 sweeps, comprising five subsamples of 50 sweeps each, were completed per grassland site during each sampling period. Foliage-dwelling spiders were collected from the three woody plant species by holding a sweep net beneath vegetation and beating the same branch with a thick wooden stick three times per individual beat. A total of 100 beats, comprising two subsamples of 50 beats each, were taken from each tree species per sample date. The spiders were sorted in the field to avoid cannibalism and prevent specimens dying and drying out. All individuals were captured with a small glass vial and preserved in a larger bottle in 70% ethanol for each sample. The specimens were then sorted into morphospecies and then identified to family level and tallied. Morphospecies were identified further to genus and species level by the second and third authors. Owing to the poor taxonomic resolution of some families (e.g. Linyphiidae, Theridiidae and Araneidae), some morphospecies could not be identified beyond family or genus level.
Guilds
The spiders collected were grouped into guilds based on their foraging methods. Spiders can be generally separated into two main groups, namely wandering spiders that actively search for prey and web-builders that construct silk structures to capture prey. The wandering spiders can be separated into ground wanderers (GW) and plant wanderers (PW), based on the substrates on which they are primarily active. Web-builders can be separated based on the structure of their webs and the following groups can be recognised: funnel-web builders (FWB), gumfoot-web builders (GWB), hackle-web builders (HWB), orb-web builders (OWB), sheetweb builders (SWB) and space-web builders (SpWB).
Statistics
Differences in monthly abundance and species richness between grassland types and between tree species were compared using repeated measures analysis of variance (ANOVA) with a Tukey-Kramer multiple comparisons post-test. A linear regression analysis was performed to determine whether seasonal rainfall had an influence on the corresponding numbers of spiders present. All of these analyses were performed in GraphPad Instat version 3.00 (2008).
The observed species richness for the grassland and tree species was calculated and the proportion of species collected per site was indicated as a percentage of the total collected for each method. Chao estimated species richness (Magurran 2004) was calculated using the equation: SChao1 = Sobs + F12 / 2F2. Chao1 is based on the abundance data available, is a function of the ratio between the singletons and doubletons in the data and reaches an asymptote when each species is represented by at least two individuals. F1 equals the number of observed species represented by one individual and F2 equals the number of observed species represented by two individuals (Magurran 2004). Inventory completeness was analysed in PAleontological STatistics (PAST) version 2.07 (Hammer, Harper & Ryan 2001) using the sample rarefaction function, which implements the 'Mao tau' analytical procedure with standard errors indicated as 95% confidence intervals on the resulting plot. Samples were arranged chronologically for the analyses. This type of analysis is based on a Chao2-type estimator, as per Colwell, Mao and Chang (2004).
We conducted three separate analyses in PAST to assess levels of similarity in spider assemblages in the three tree species and four grassland types. Firstly, we conducted a one-way analysis of similarity (ANOSIM) procedure to determine similarity in spider assemblage structure based on the tree and grassland sample totals. ANOSIM can be considered analogous to one-way ANOVA and is performed on the underlying Bray-Curtis similarity matrix. Spider abundance was transformed to absence or presence data prior to analysis. The results were presented as a global R-value that is analogous to the F-value calculated using ANOVA. Pair-wise comparisons between the tree or grassland types were presented as R-values that are analogous to the post hoc comparisons of ANOVA (Clarke & Warwick 2001). A R-value close to 1 indicates high dissimilarity between the two compared assemblages, whilst a value close to 0 indicates high similarity in their composition. Secondly, we performed a non-metric multidimensional scaling (MDS) on the sample data from each grassland type or tree species based on a Bray-Curtis similarity matrix. The ordination diagrams produced group sites together based on similarities in their species composition, whilst also taking species abundance into account. Samples that were most similar were grouped close to one another. We added convex hulls to each ordination diagram to group samples from each grassland type and tree species. Lastly, we carried out a cluster analysis of the Bray-Curtis similarity matrix using the cluster function with a paired group algorithm. Taking both MDS and cluster analysis results into consideration allows for more accurate interpretation of the assemblage patterns (Clarke & Warwick 2001).
Results
Grassland
A total of 1649 spiders were collected from the four grassland types, representing 15 families and 82 species (see Online Appendix). The species richness of each of the grasslands varied between 38 and 56 and the number of unique species sampled from each (Table 1) can largely be attributed to the sampling of singletons and doubletons. The rarefaction curve of the grassland samples did not reach an asymptote, indicating that further sampling would be necessary to sample all of the species present in the grasslands (Figure 1a).
Of the species sampled, 54 were hunting spiders (65.85%) whilst 28 species were web-builders (34.15%). The numerically dominant species collected were Tibellus minor Lessert, 1919 (Philodromidae, 10.79%), Misumenops rubrodecoratus Millot, 1941 (Thomisidae, 10.49%), Thyene thyenioides (Peckham & Peckham, 1903) (Salticidae, 9.16%), Runcinia flavida (Thomisidae, 6.19%) and Tibellus hollidayi Lawrence, 1952 (6.06%) (see Online Appendix). All of these species are active hunting spiders and contributed largely to the dominance of Philodromidae, Thomisidae and Salticidae in the survey, whilst Araneidae, which were also relatively abundant, had several species contributing to the overall abundance of the family (Figure 2a). Other families were not universally abundant but were only prominent in one, two or three types of grasslands. Araneidae and Salticidae were clearly more abundant in the mixed grassland, Thomisidae were most prominent in the woodland grassland, whilst Philodromidae were considerably lower in abundance in the woodland grassland compared to the three other grassland types (Figure 2a).
Monthly spider abundance did not differ significantly between the four grassland types (ANOVA: -F37 = 0.6461, p = 0.5940). However, monthly species richness differed significantly between sites (ANOVA: F37 = 9.086, p = 0.0005).
Species richness was significantly higher in mixed grassland (p < 0.001) and Themeda grassland (p < 0.01) than in weedy grassland and also significantly higher in mixed than in woodland grassland (p < 0.05).
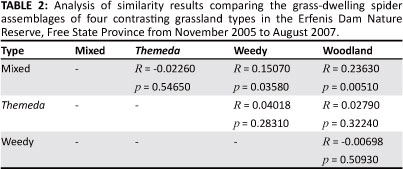
The ANOSIM comparisons of the grass-dwelling spider assemblages (Table 2) pointed to high levels of similarity between grassland spider assemblages (Global R = 0.0685, p = 0.0786). The pair-wise comparisons between sites showed significant dissimilarity between the assemblages of the mixed and woodland grasslands, and between mixed and weedy grasslands, but none of the other compared assemblages differed significantly from one another (Table 2). This was supported by the results of the MDS ordination (Figure 3a) and cluster analysis (Figure 3b), which showed a large degree of overlap between the assemblages of the four grasslands. It is interesting to note that on several occasions the assemblages of the four grassland types clustered together based on the month of sampling (e.g. November 2005, May 2006 and February 2007) or were in adjacent groups within the analysis (February 2006, August 2006 and May 2007), rather than grouping together based on grassland type (Figure 3b).
Regarding phenology, spider abundance appears not to be directly correlated to the rainfall of that particular season (R2 = 0.0231), but rather to preceding rainfall history, suggesting a lag effect (Figure 4a). This pattern may have been compounded in February 2006 by the very high rainfall that month, which may have restricted spider activity during this period, accounting for the drastic drop in spider abundance. However, in November 2005, high numbers of grass-dwelling spiders were collected, even though rainfall was low in the previous three months.
Foliage
A total of 496 spiders were collected by foliage-beating, representing 17 families and 52 species (Online Appendix). Of the three species, S. ciliata had much higher species richness and many more unique species; A. karroo and S. lancea had a similar number of unique species (Table 1). The rarefaction curve for the tree samples also did not reach an asymptote (Figure 1b) and the slope was noticeably steeper than in the case of the grassland samples (Figure 1a), indicating that a considerable number of species occurring on the woody plants were not sampled.
Compared to the grass-dwelling fauna, a slightly lower proportion of the foliage-dwellers were hunting spiders (32 spp., 61.54%); 20 species (38.46%) construct webs to capture prey. The most abundant foliage-dwelling species were Philodromus browningi Lawrence, 1952 (Philodromidae, 13.91%), Cheiracanthium furculatum Karsch, 1879 (Miturgidae, 9.88%), M. rubrodecoratus (8.47%), Baryphas ahenus Simon, 1902 (Salticidae, 6.65%) and Neoscona subfusca (C.L. Koch, 1837) (Araneidae, 5.47%). The abundance of these species, particularly P. browningi and M. rubrodecoratus, were largely responsible for shaping patterns of family dominance (Figure 2b). Philodromidae and Thomisidae were most common on A. karroo, whilst Salticidae, Theridiidae and Araneidae were more abundant on S. ciliata than the other woody species. Whilst all of the families showed low abundance on S. lancea, Theridiidae was totally absent from this species (Figure 2b).
Differences in spider abundance between tree species were very significant (ANOVA: F27 = 6.499, p = 0.0072). Both A. karroo (n = 206) and S. ciliata (n = 220) had significantly more spiders than the 65 individuals collected from S. lancea (p < 0.05), but differences between A. karroo and S. ciliata were not significant (p > 0.05). Monthly species richness also differed significantly between tree species (ANOVA: F27 = 7.842, p = 0.0052), although significant differences in the pairwise comparisons were only detected between S. ciliata and S. lancea (p < 0.01).
The ANOSIM results indicated considerable overlap between assemblages of the tree species (Global R = 0.3664, p < 0.0001). The pair-wise comparisons indicated the greatest dissimilarity between the assemblages of S. lancea and S. ciliata (R = 0.5145, p = 0.0003), with increasing similarity between the assemblages of A. karroo and S. lancea (R = 0.3078, p = 0.0036) and between A. karroo and S. ciliata (R = 0.2698, p = 0.0151). The MDS ordination of the arboreal spider assemblages (Figure 3c) supported these results and indicated that S. lancea has the most distinctive assemblage, whilst there was considerable overlap (and thus greater similarity) between the assemblages of A. karroo and S. ciliata. However, in the cluster analysis none of the trees' samples clustered together in a single group, although six of the eight S. lancea samples fell within adjoining groups (Figure 3d). This supported the MDS results that indicated this tree as having the most distinct spider assemblage.
As for the grass-dwelling fauna, rainfall in the season of sampling did not have a significant influence on the numbers of spiders in the tree foliage (R2 = 0.0976). Similar depressions in spider abundance were observed during the summer of 2006 and during the winter months, with spider abundance peaking during the spring of the 2005-2006 year of sampling, and during the spring or summer of the 2006-2007 year of sampling, depending on the tree species (Figure 4b).
Ethical considerations
This study was made possible through collecting permits from Free State Nature Conservation. The study follows the ethical guidelines proposed for the SANSA, approved by the SANSA steering committee.
Discussion
This study provided the first comprehensive dataset on the grass-dwelling and foliage-dwelling spiders of the Grassland Biome of South Africa within an ecological context. In part, the study aimed to assess the influence of vegetation diversity and structure on spider assemblages. As spiders are predators, they interact with the vegetation indirectly, which means that structural support available for webs and active hunting, prey availability and suitable microclimate may influence the composition of a spider community (e.g. Dippenaar-Schoeman & Wassenaar 2006; Gunnarsson 1990; Harwood, Sunderland & Symondson 2001; Haughton et al. 1999; Spears & MacMahon 2012; Uetz & Burgess 1979). Spider abundance and diversity may be correlated with the specific vegetation characteristics, suggesting that the availability of habitats is important for spider colonisation and establishment (jögar, Metspalu & Hiiesaar 2004).
Free State grasslands consist of a high diversity of grasses and shrubs that create greater habitat complexity, which means that there are many niches in which spiders may live (Haddad 2005). Yet, as a habitat, grasses appear quite uniform in structure, which may limit the availability of structurally different niches in grasslands, despite high overall grass species richness. Of the grasslands studied here, the mixed, woodland and weedy grasslands all had a more complex structure than the uniform Themeda grassland. Yet, the structurally complex weedy grassland has considerably lower spider species richness and projected species richness than the other habitats, despite quite similar abundance (Table 1). According to Mgobozi, Somers and Dippenaar-Schoeman (2008), the progressive invasions of the invasive alien weed Chromolaena odorata (L.) King & Robinson (Asteraceae) brings with it negative changes in native spider abundance, assemblage patterns, diversity and estimated species richness. It could be that the weeds present in the weedy grassland emitted allomones, which may have repelled many insects. This, coupled with a decrease in suitable plant species for phytophagous insects, may have caused a decrease in food resources for spiders, effectively contributing to the lower spider diversity.
The estimated species richness predicted that the Themeda grassland would have the highest species richness (Table 1). Yet, the mixed grassland was the most diverse habitat (structurally, as well as having the most plant species) and also had the most species rich spider assemblage. Although the Themeda grassland was very structurally uniform, it was very closely the second most species rich grassland. This is probably the result of the complex structure of the grass stems and inflorescences and the very dense tussocks that T. triandra form (less dense in the other grasses). In the present study, a similar number of species was collected in the Themeda grassland (54 spp.) compared to another study conducted in similarly homogenous Themeda grassland in the central Free State (Haddad 2005), in which 57 species were collected. The species richness in the woodland grassland was only slightly lower than the mixed and Themeda grasslands and may have been expected to be higher as a result of the presence of woody plants, which could have created a more structurally complex habitat with more niches available for occupation by spiders. Thus, expected migration between grasses and woody plants is apparently minimal and does not necessarily create more complex grass-dwelling assemblages.
The estimated species richness for the three woody plants sampled predicted that S. lancea would have the highest species richness and A. karroo the least, but the S. ciliata shrubs had greater observed species richness and also more unique species than the two trees (Table 1). This could be the result of the denser foliage of S. ciliata than that of the trees, which provides greater microhabitat complexity in the foliage of the plant. This can also be illustrated by the high abundance of web-building Theridiidae in these shrubs, but their absence or scarcity in the two tree species. The two tree species also have thick trunks that are covered in coarse bark, which contributes to the structural complexity of these trees, but because only peripheral foliage was sampled the contribution that trunk-dwelling spiders could make to the assemblage structure of these trees was minimal.
Whilst the grassland fauna showed a higher species richness than the foliage-dwelling fauna, this may be an artefact of collecting effort, which is reflected in the total abundance collected by the two methods (number of foliage spiders was approximately one-third of the grassland spiders, with 40% of the number of sample units taken from grassland), as well as the slopes of the respective rarefaction curves. The steeper slope of the curve for foliage-dwellers indicates that considerably more species could still be sampled than in the case of the grassland fauna. Considering the contrasting physical structures of the three woody species sampled, it was unusual to find that the two related species (S. lancea and S. ciliata) had the most different spider assemblages and that there was considerable overlap in the assemblages of S. ciliata with A. karroo. Although a slightly larger number of unique species was collected from the woody vegetation, and the assemblages harboured by each tree or shrub species were generally more distinctive than in the case of the grass-dwelling fauna, it is likely that further sampling from the woody plants (to collect comparable numbers of individuals from the two strata) would have considerably reduced the number of singleton and doubleton species and effectively reduced the number of unique species apparently occurring on the woody plants. This may also have increased the species richness of the foliage fauna considerably, which may have produced higher levels of similarity between tree species. As such, the sampling discrepancies may mask the true levels of overlap in the assemblages of the woody plants.
Both hunting and web-building spiders are sensitive to changes in habitat structure and rainfall can affect habitat structure in subtle ways (Langlands, Brennan & Pearson 2006). In their study, conducted in the Great Victoria Desert in Australia, sampling years with high spider abundance coincided with years of good rainfall that took place prior to sampling (Langlands et al. 2006). This was also evident in the present study, with spider numbers increasing following months with good rainfall and decreasing following months with low rainfall. A bottom-up effect from rainfall might be responsible for the increases in spider abundance; however, the linear regression test results showed that rainfall of a particular season had no significant influence on spider abundance of that season in both the foliage and grass layers. Our data would suggest that there is a lag between the times that rain falls and increased or decreased spider abundance; in the case of an increase this could be explained by the time needed for plants to grow and create more complex niches, which, in turn, attract a greater number of prey for spiders. At a finer temporal scale, rain falling immediately before or during the sampling day may also influence spider activity and hence capture rates (Romero & Vasconcellos-Neto 2003). This may explain the low number of spiders captured in February 2006, following several days of heavy rainfall.
According to Langlands et al. (2006), some spiders tend to have strong seasonality, so this could also influence the number of spiders present during a year (e.g. Van den Berg & Dippenaar-Schoeman 1991). Seasonal abundance of tree spiders sometimes differs from grass-dwelling spiders (Dippenaar-Schoeman, Van den Berg & Van den Berg 1989), although our study found a similar general pattern in both strata. Most studies show peaks in temporal patterns of spider numbers and species during early to mid-summers (Hatley & MacMahon 1980; Niemela et al. 1994). More spiders were captured in the summer months than in the winter months for both strata, which reflect the high availability of prey and emergence of immature spiders from egg sacs during spring. Lower prey availability during the winter is not problematic to spiders because they are very resistant to starvation and can lower their metabolic rates through inactivity, which enables them to survive harsh conditions (Dippenaar-Schoeman & Jocqué 1997). According to Aitchison (1987), there are winter-active spiders that utilise this season to gain an ecological advantage of little competition from other spiders or other predaceous invertebrates for food, or alternately are species that feed primarily on winter-active prey (e.g. Dippenaar-Schoeman, De Jager & Van den Berg 1996). A good example from the present study is the Archaeodictyna sp. (Dictynidae), of which most specimens were collected in autumn and winter. However, most of the species appear to be strongly affected by the onset of winter, during which spider numbers were at their lowest.
It is not surprising that the dominant species of both strata came from Araneidae, Salticidae and Thomisidae, as these are some of the most diverse families of spiders (Dippenaar-Schoeman & Jocqué 1997). These families have also dominated the plant-dwelling fauna in other South African studies in grassland (Haddad 2005), savannah (Dippenaar et al. 2008; Dippenaar-Schoeman, Van den Berg & Prendini 2009; Dippenaar-Schoeman et al. 1989;
Haddad et al. 2010; Muelelwa et al. 2010) and forest habitats (Dippenaar-Schoeman & Wassenaar 2006). Curiously, only a single species, M. rubrodecoratus, was abundant in both strata, indicating that this species is very flexible in its ecological requirements, whilst the other more abundant species are more particular in their microhabitat preferences.
Conclusion
The present study provided comprehensive data on the biodiversity of grass-dwelling and foliage-dwelling spiders in the Grassland Biome and tried to explain differences in assemblage structure in relation to vegetation structure and diversity. The dominant plant-dwelling families from both strata were the Araneidae, Philodromidae, Salticidae and Thomisidae, although only one species (M. rubrodecoratus) was abundant in both strata. Grass-dwelling assemblages were richer in species than foliage-dwellers and this was likely an artefact of sampling intensity. Sampling protocols should be adapted to have an equal number of sweeps and beats to collect more comparable numbers of specimens, which may better explain species richness and assemblage composition patterns. Grassland assemblages are generally similar and show considerable overlap in species, indicating that the grass fauna is probably more flexible in their ecological requirements and can occupy more structurally variable plant communities within the much more extensive grassy habitat. This suggests limited effects of plant diversity and structural complexity in shaping assemblages. The foliage-dwelling spider communities differed somewhat from one another, suggesting that either the sampling intensity was inadequate or that the vegetation structures were indeed markedly different, affecting community structure through niche specialisation. Estimations of total species richness (Chao1) for grassland habitats and tree species suggest that grasslands consistently maintain much higher species richness than the isolated woody plants. As such, trees may function as islands within the large grassland matrix, supporting very different assemblages than the surrounding grassy habitat, but with their isolation contributing to lower levels of species richness than the surrounding continuous grassland habitat.
Acknowledgements
We thank the Erfenis Dam Nature Reserve staff, particularly the conservation manager Robert Lotze, for support during the study and permission to collect in the reserve. We would also like to thank Robin Lyle, Dewald du Plessis and Joan Adendorff for occasional assistance with fieldwork. Andor Venter (University of the Free State) kindly identified some of the plant species. This study forms part of the first author's MSc at the University of the Free State and was funded through a National Research Foundation of South Africa (NRF) grant in the NRF Thuthuka programme to the second author (Grant number TTK2008050500003). The two anonymous referees provided useful comments and suggestions to improve the quality of the manuscript and their inputs are greatly appreciated.
Competing interests
The authors declare that they have no financial or personal relationships that may be inappropriately affected through the production of this article.
Authors' contributions
This study formed part of R.F.'s (University of the Free State) MSc on spider ecology in the Erfenis Dam Nature Reserve. R.F. conducted field work, sorted and tallied specimens, performed some statistical analyses and wrote part of the manuscript. C.R.H. (University of the Free State) supervised R.F., performed initial identifications, performed some statistical analyses and wrote part of the manuscript. A.S.D-S. (Agricultural Research Council - Plant Protection Research Institute) conducted final identifications and contributed to writing the manuscript. A.G. (University of the Free State) assisted with field work and sorting specimens.
References
Aitchison, C.W., 1987, 'Feeding ecology of winter-active spiders', in W. Nentwig (ed.), Ecophysiology of spiders, pp. 264-273, Springer-Verlag, Berlin. http://dx.doi.org/10.1007/978-3-642-71552-5_19 [ Links ]
Bredenkamp, G.J., Spada, F. & Kazmierczak, E., 2002, 'On the origin of northern and southern hemisphere grasslands', Plant Ecology 163, 209-229. http://dx.doi.org/10.1023/A:1020957807971 [ Links ]
Butler, V.P. & Haddad, C.R., 2011, 'Spider assemblages associated with leaf litter of three tree species in central South Africa (Arachnida: Araneae)', African Journal of Ecology 49, 301-310. http://dx.doi.org/10.1111/j.1365-2028.2011.01265.x [ Links ]
Carbutt, C., Tau, M., Stephens, A. & Escott, B., 2011, 'The conservation status of temperate grasslands in southern Africa', Grassroots 11, 17-23. [ Links ]
Clarke, K.R. & Warwick, R.M., 2001, Change in marine communities: An approach to statistical analysis and interpretation, 2nd edn., PRIMER-E, Plymouth. [ Links ]
Colwell, R.K., Mao, C.X. & Chang, J., 2004, 'Interpolating, extrapolating, and comparing incidence-based species accumulation curves', Ecology 85, 2717-2727. http://dx.doi.org/10.1890/03-0557 [ Links ]
Dippenaar-Schoeman, A.S. & Jocqué, R., 1997, African spiders: An identification manual, Plant Protection Research Institute handbook 9, Agricultural Research Institute, Pretoria. [ Links ]
Dippenaar-Schoeman, A.S. & Wassenaar, T.D., 2006, 'A checklist of spiders from the herbaceous layer of a coastal dune forest ecosystem at Richards Bay, KwaZulu-Natal, South Africa (Arachnida: Araneae)', African Invertebrates 47, 63-70. [ Links ]
Dippenaar-Schoeman, A.S., De Jager, M. & Van den Berg, A., 1996, 'Ammoxenus species (Araneae: Ammoxenidae) - Specialist predators of harvester termites in South Africa', African Plant Protection 2, 103-109. [ Links ]
Dippenaar-Schoeman, A.S., Hamer, M. & Haddad, C.R., 2011, 'An annotated checklist of the spiders (Arachnida: Araneae) of the Mkambathi Nature Reserve, Eastern Cape, South Africa', Koedoe 53, 1-10. [ Links ]
Dippenaar-Schoeman, A.S., Van den Berg, A. & Prendini, L., 2009, 'Spiders and scorpions (Arachnida: Araneae, Scorpiones) of the Nylsvley Nature Reserve, South Africa', Koedoe 51, 1-9. http://dx.doi.org/10.4102/koedoe.v51i1.161 [ Links ]
Dippenaar-Schoeman, A.S., Van den Berg, A.M. & Van den Berg, A., 1989, 'Species composition and relative seasonal abundance of spiders from the field and tree layers of the Roodeplaat Dam Nature Reserve', Koedoe 32, 25-38. http://dx.doi.org/10.4102/koedoe.v32i1.462 [ Links ]
Dippenaar, S.M., Modiba, M.A., Khoza, T.T. & Dippenaar-Schoeman, A.S., 2008, 'A checklist of the spiders (Arachnida, Araneae) of the Polokwane Nature Reserve, Limpopo Province, South Africa', Koedoe 50, 10-17. http://dx.doi.org/10.4102/koedoe.v50i1.128 [ Links ]
Egoh, B.N., 2009, Integrating ecosystem services into conservation planning in South Africa, Unpublished PhD thesis, Department of Botany and Zoology, Stellenbosch University. [ Links ]
Foord, S.H., Dippenaar-Schoeman, A.S. & Haddad, C.R., 2011, 'South African spider diversity: African perspectives on the conservation of a mega-diverse group', in O. Grillo & G. Venora (eds.), Changing diversity in changing environment, pp. 163-182, InTech Publishing, Rijeka. [ Links ]
Germishuizen, G., Meyer, N.L., Steenkamp, Y. & Keith, M. (eds.), 2006, A checklist of South African plants, South African Botanical Diversity Network report 41, SABONET, Pretoria. [ Links ]
GraphPad InStat version 3.00 for Windows 95, 2008, computer software, GraphPad Software, San Diego. [ Links ]
Gunnarsson, B., 1990, 'Vegetation structure and the abundance and size distribution of spruce-living spiders', Journal of Animal Ecology 59, 743-752. http://dx.doi.org/10.2307/4892 [ Links ]
Haddad, C.R., 2005, 'Ecology of spiders (Arachnida: Araneae) inhabiting Themeda triandra Forsskál grassland in semi-arid South Africa', Navorsinge van die Nasionale Museum Bloemfontein 21, 25-36. [ Links ]
Haddad, C.R. & Dippenaar-Schoeman, A.S., 2002, 'The influence of mound structure on the diversity of spiders (Araneae) inhabiting the abandoned mounds of the snouted harvester termite Trinervitermes trinervoides', Journal of Arachnology 30, 403-408. http://dx.doi.org/10.1636/0161-8202(2002)030[0403:TIOMSO]2.0 .CO;2 [ Links ]
Haddad, C.R. & Dippenaar-Schoeman, A.S., 2006, 'Spiders (Araneae) inhabiting abandoned mounds of the snouted harvester termite Trinervitermes trinervoides (Sjöstedt) (Isoptera: Termitidae: Nasutitermitinae) in the Free State, South Africa, with notes on their biology', Navorsinge van die Nasionale Museum Bloemfontein 22, 1-15. [ Links ]
Haddad, C.R., Honiball, A.S., Dippenaar-Schoeman, A.S., Slotow, R. & Van Rensburg, B.J., 2010, 'Spiders (Arachnida: Araneae) as indicators of elephant-induced habitat changes in the Maputaland Centre of Endemism, South Africa', African Journal of Ecology 48, 446-460. http://dx.doi.org/10.1111/j.1365-2028.2009.01133.x [ Links ]
Hammer, 0., Harper, D.A.T. & Ryan, P.D., 2001, 'PAST - PAlaeontological STatistics', Palaeo - Electronica 4, 1-33. [ Links ]
Harwood, J.D., Sunderland, K.D. & Symondson, W.O.C., 2001, 'Living where the food is: Web location by linyphiid spiders in relation to prey availability in winter wheat', Journal of Applied Ecology 38, 88-99. http://dx.doi.org/10.1046/j.1365-2664.2001.00572.x [ Links ]
Hatley, C.L. & MacMahon, J.A., 1980, 'Spider community organization: Seasonal variation and the role of vegetation architecture', Environmental Entomology 9, 632-639. [ Links ]
Haughton, A.L., Bell, J.R., Boatman, N.D. & Wilcox, A., 1999, 'The effects of different rates of the herbicide glyphosate on spiders in arable field margins', Journal of Arachnology 27, 249-254. [ Links ]
Jansen, R., Makaka, L., Little, I.T. & Dippenaar-Schoeman, A.S., in press, 'Response of ground-dwelling spider assemblages (Arachnida, Araneae) to montane grassland management practices in South Africa', Journal of Insect Conservation. [ Links ]
Jögar, K., Metspalu, L. & Hiiesaar, K., 2004, 'Abundance and dynamics of wolf spiders (Lycosidae) in different plant communities', Agronomy Research 2, 145-152. [ Links ]
Langlands, P.R., Brennan, K.E.C. & Pearson, D.J., 2006, 'Spiders, spinifex, rainfall and fire: Long-term changes in an arid spider assemblage', Journal of Arid Environments 67, 36-59. http://dx.doi.org/10.1016/j.jaridenv.2006.01.018 [ Links ]
Lotz, L.N., Seaman, M.T. & Kok, D.J., 1991, 'Surface-active spiders (Araneae) of a site in semi-arid central South Africa', Navorsinge van die Nasionale Museum Bloemfontein 7, 530-540. [ Links ]
Low, A.B. & Rebelo, A.G., 1996, Vegetation of South Africa, Lesotho and Swaziland, Department of Environmental Affairs and Tourism, Pretoria. [ Links ]
Magurran, A.E., 2004, Measuring biological diversity, Blackwell Publications, Bodmin. [ Links ]
Mentis, M.T. & Huntley, B.L., 1982, A description of the Grassland Biome project, South African National Scientific Programmes report 62, Graphic Arts Division of the Centre for Scientific and Industrial Research, Pretoria. [ Links ]
Mgobozi, M.P., Somers, M.J. & Dippenaar-Schoeman, A.S., 2008, 'Spider responses to alien plant invasion: The effect of short- and long-term Chromolaena odorata invasion and management', Journal of Applied Ecology 45, 1189-1197. [ Links ]
Mucina, L. & Rutherford, M.C., 2006, The vegetation of South Africa, Lesotho and Swaziland, Strelitzia 19, South African National Biodiversity Institute, Pretoria. [ Links ]
Muelelwa, M.I., Foord, S.H., Dippenaar-Schoeman, A.S. & Stam, E.M., 2010, 'Towards a standardized and optimized protocol for rapid assessments: Spider species richness and assemblage composition in two savannah vegetation types', African Zoology 45, 273-290. http://dx.doi.org/10.3377/004.045.0206 [ Links ]
Niemela, J., Pajunen, T., Haila, Y., Punttila, P. & Halme, E., 1994, 'Seasonal activity of boreal forest-floor spiders (Araneae)', Journal of Arachnology 22, 23-31. [ Links ]
O'Connor, T.G. & Kuyler, P., 2005, 'National Grasslands Initiative: Identification of compatible land uses for maintaining compatible biodiversity integrity', Unpublished report, South African National Biodiversity Institute, Pretoria. [ Links ]
Palmer, A.R. & Ainslie, A.M., 2005, 'Grasslands of South Africa', in J.M. Suttie, S.G. Reynolds & C. Batello (eds.), Grasslands of the world, pp. 77-120, Plant production and protection series 34, Food and Agriculture Organization of the United Nations, Rome. [ Links ]
Romero, G.O. & Vasconcellos-Neto, J., 2003, 'Natural history of Misumenops argenteus (Thomisidae): Seasonality and diet on Trichogoniopsis adenantha (Asteraceae)', Journal of Arachnology 31, 297-304. http://dx.doi.org/10.1636/02-19 [ Links ]
Spears, L.R. & MacMahon, J.A., 2012, 'An experimental study of spiders in a shrub-steppe ecosystem: The effects of prey availability and shrub architecture', Journal of Arachnology 40, 218-227. http://dx.doi.org/10.1636/P11-87. [ Links ]!
Stuart, C. & Stuart, T., 1997, Guide to southern African game and nature reserves, 4th edn., Struik Publishers, Cape Town. [ Links ]
Uetz, G.W. & Burgess, J., 1979, 'Habitat structure and colonial behaviour in Metepeira spinipes (Araneae: Araneidae), an orb-weaving spider from Mexico', Psyche 86, 79-90. http://dx.doi.org/10.1155/1979/76206 [ Links ]
Van den Berg, A. & Dippenaar-Schoeman, A.S., 1991, 'Ground-living spiders from an area where the harvester termite Hodotermes mossambicus occurs in South Africa', Phytophylactica 23, 247-253. [ Links ]
Van Wyk, B. & Van Wyk, P., 1997, Field guide to trees of southern Africa, Struik Publishers, Cape Town. [ Links ]
Venter, H.J.T. & Joubert, A.M., 1984, Climbers, tree and shrubs of the Orange Free State, P.J. de Villiers Publishers, Bloemfontein. [ Links ]
Wessels, K.J., Reyers, B., Van Jaarsveld, A.S. & Rutherford, M.C., 2003, 'Identification of potential conflict areas between land transformation and biodiversity conservation in north-eastern South Africa', Agriculture, Ecosystems and Environment 95, 157-178. http://dx.doi.org/10.1016/S0167-8809(02)00102-0 [ Links ]
 Correspondence:
Correspondence:
Charles Haddad
PO Box 339, Bloemfontein 9300, South Africa
haddadcr@ufs.ac.za
Received: 18 Sept. 2012
Accepted: 29 Jan. 2013
Published: 09 Apr. 2013

