










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.54 n.1 Pretoria Jan. 2012
ORIGINAL RESEARCH
Invertebrates on isolated peaks in the uKhahlamba-Drakensberg Park World Heritage Site, South Africa
Adrian J. ArmstrongI; Robert F. BrandII
IConservation Planning Division, Ezemvelo KZN Wildlife, South Africa
IIEnvironmental, Water and Earth Sciences, Tshwane University of Technology, South Africa
ABSTRACT
A survey to document and describe the alpine flora and various focal faunal taxa on six isolated inselberg-like peaks (total area of 31.9 ha), all 3000 m or higher, located in the uKhahlamba-Drakensberg Park World Heritage Site, South Africa, was undertaken in early summer in 2005. Study of the fauna of these peaks should be informative because the impacts of controllable anthropogenic threats on the invertebrate communities on them should be minimal or absent in comparison with those on the main massif. A total of 341 invertebrate individuals representing 61 species were recorded from the focal taxa (Oligochaeta, Gastropoda and certain groups of Insecta, i.e. focal taxa within the Blattoidea, Dermaptera, Orthoptera, Hemiptera, Diptera, Lepidoptera, Coleoptera and Hymenoptera). The 61 species recorded consisted of two species from the Oligochaeta, one species from the Gastropoda and 58 species from the Insecta. Eleven species (one from the Oligochaeta, ten from the Insecta) are endemic and 11 species (one from the Oligochaeta, ten from the Insecta) are probably endemic to the Drakensberg Alpine Centre, constituting 36.1% of the total species recorded. The results suggest that the Drakensberg Alpine Centre (DAC), as for plants, is a centre of endemism for invertebrates. Cluster analysis showed that the species composition of the two northern peaks, Sentinel and Eastern Buttress, clustered together, separate from a cluster formed by the Outer Horn, Inner Horn and Dragon's Back and from the cluster formed by the southernmost peak, Cathkin. Non-metric multi-dimensional scaling results indicated that distance from the Sentinel, the most northerly peak sampled, and mean minimum temperature for July had the strongest correlations with the species data, reflecting change over a straight-line distance of nearly 60 km in a south-easterly direction.
CONSERVATION IMPLICATIONS: Only a small proportion (ca. 5.5%) of the DAC is conserved, the majority of which lies in the uKhahlamba-Drakensberg Park World Heritage Site. Conservation of more of the DAC, including more of its latitudinal extent, is required to adequately conserve its unique plant and invertebrate communities.
Introduction
The invertebrate fauna of the Drakensberg Alpine Centre (DAC), including the alpine region of the uKhahlamba-Drakensberg Park World Heritage Site in KwaZulu-Natal with its escarpment and peaks over 3000 m, has not been as well studied as some areas at lower altitudes in the province, for example, the World Heritage Site foothills and some of the midlands, coastal and savanna regions (Bazelet & Samways 2011; Lovell et al. 2010; Pryke & Samways 2001, 2003, 2012; Uys, Hamer & Slotow 2006, 2009, 2010; see also Clark, Barker & Mucina 2011). Consequently little is known regarding invertebrate distributions and invertebrate community composition and structure on high-altitude inselbergs in the Drakensberg. The DAC is located in South Africa - in parts of KwaZulu-Natal (KZN), the Eastern Cape and Free State - and in Lesotho. It contains a unique assemblage of plants that is rich in endemics, and is an important centre of plant diversity in southern Africa (Killick 1978; Van Wyk & Smith 2001). Approximately 2520 angiosperm (higher plant) taxa, of which 334 (13%) are endemics, occur in the DAC (Carbutt & Edwards 2004, 2006; Van Wyk & Smith 2001). The DAC is defined by floristic criteria (Killick 1990), but was designated as 'alpine' based on climatological criteria (Van Wyk & Smith 2001). The Centre can be divided into a subalpine zone (c. from 1800 m a.s.l. to 2800 m a.s.l.) and an alpine zone (c. from 2800 m a.s.l. to 3482 m a.s.l.; Killick 1978; Van Wyk & Smith 2001).
The peaks were chosen for several reasons: their altitude (situated within the alpine zone), the assumed infrequent nature or exclusion of the two major disturbance factors along the Drakensberg escarpment, namely, fire and herbivory by domestic livestock, and the isolation of the peaks, or inselbergs, which can be regarded as an archipelago of 'islands', which theoretically could house unique, high-altitude endemics. Fire frequency, extent and intensity, as well as season of burn and time since last burn can influence the composition of invertebrate assemblages at lower altitudes in the uKhahlamba-Drakensberg Park World Heritage Site (Uys 2006; Uys et al. 2006, 2010). Invertebrate diversity is lower in grassland under communal grazing in comparison to grassland under nature conservation in a region south-east of the uKhahlamba-Drakensberg Park World Heritage Site (Robertson & Van Noort 2003; Wildy 2003).
The results of an invertebrate survey carried out between 28 November and 05 December 2005 are presented here. The goal of the invertebrate survey was to sample six out of a possible 11 isolated peaks at altitudes of 3000 m or more in the uKhahlamba-Drakensberg Park World Heritage Site in KwaZulu-Natal. The peaks were chosen to ensure that a sufficient latitudinal and longitudinal range was incorporated by the sampling and that the peaks were large enough to safely land a helicopter and to establish sample plots adequate for invertebrate and vegetation sampling.
Aims
The aims of this survey were to identify invertebrate species of focal groups that occur on the isolated peaks in the alpine zone of the KwaZulu-Natal section of the DAC and to give some indication of whether:
- The DAC is a centre of endemism for invertebrates, where a relatively high proportion of invertebrate species are endemic.
- Invertebrate communities change from isolated peak to isolated peak, and possible reasons for this change.
Research method and design
Invertebrate species targeted
The invertebrate groups targeted during sampling included those known to have numbers of species endemic to mountainous areas of South Africa (e.g. Armstrong, Van Hensbergen & Geertsema 1994), for some groups owing to a proportion or all of them being flightless or for which taxonomic expertise and/or aids to identification of species were available in South Africa. These focal animal groups were as follows (other reasons for their inclusion in the study are given in parentheses): earthworms (soil engineers), snails (herbivores), large cockroaches (plant detritivores), wingless grasshoppers (herbivorous on forbs), leafhoppers and other bugs (herbivores, predators), beetles (herbivorous on various plant parts, predators of invertebrates, detritivores, decomposers), crane flies and robber flies (decomposers, insect predators), satyrid butterflies (herbivorous on grasses) and bees (pollinators).
Only adults of each species were included in the analyses because juveniles do not necessarily have the specific diagnostic identification characters. Specimens were identified to species through: (1) reference to identified specimens and sometimes also to unidentified specimens housed in several collections, that is, the KwaZulu-Natal Museum in Pietermaritzburg (NMSA), and the South African National Collection of Insects (SANC) and the Ditsong National Museum of Natural History, formerly the Transvaal Museum (TMSA), in Pretoria, (2) identification keys, published taxonomic papers and a regional fauna guide (Alexander 1964; Brown 1962; Eardley 1988; Eardley, Kuhlmann & Pauly 2010; Fennah 1967; Iwan 2001; Londt 1981; Marshall 1904; Princis 1963; Pringle, Henning & Ball 1994; Slater 1964; Straneo 1958; Synave 1956; Theron 1986; Wittmer 1949) and (3) specialist taxonomists (see Acknowledgements). The distribution ranges and endemicity of the species were ascertained or estimated from specimen label data and distribution records contained in the publications listed above and in the Biodiversity Database of Ezemvelo KZN Wildlife, and from Armstrong et al. (1994), Armstrong and Van Hensbergen (1997a, 1997b, 1999), Arnott (2006), Hamer and Slotow (2007) and Uys et al. (2006, 2010).
Study sites
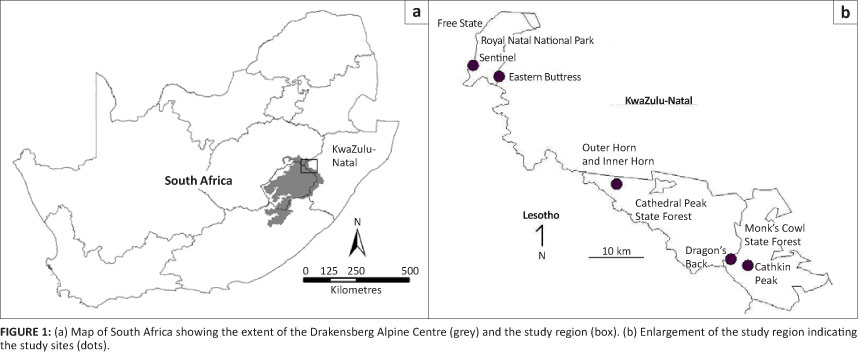
Six isolated peaks, all greater than 3000 m in elevation, were sampled during the survey, respectively occurring from north to south: the Sentinel, Eastern Buttress, Inner Horn, Outer Horn, Dragon's Back and Cathkin (Figures 1a and 1b). A 50 m by 50 m (0.25 ha) plot was demarcated using a tape-measure-calibrated Ranging TLR 75 rangefinder and four orange-dyed, 1.5 m high, wooden dowels on a northerly-facing slope (where possible, one side was aligned to an approximately 59° magnetic bearing) of similar topographic profile on each peak. The top of the profile was a rocky slope and the bottom of the profile was more or less flat, generally with small shallow pools of standing water. Topographic constraints (only precipitous northerly slopes) on the Outer Horn meant that the plot was predominantly flat with some of it on a southerly-facing slope. The co-ordinates of the centre of the plot at each site were recorded using a global positioning system (GPS) (except for the Sentinel plot, where the co-ordinates were obtained from a 1:50 000 scale topographic map). The surface area in hectares of each peak was calculated from the track log of a GPS used to record these areas, and from a 1:30 000 orthophoto for the Dragon's Back peak. A brief description of each plot and various parameters associated with each are given in Table 1.
Each plot was sampled by Adrian J. Armstrong for 15 minutes of search time (i.e. excluding time used to record or capture and store specimens), in three phases, by means of a slow meandering walk through the plot. One period of five minutes was concentrated on searching bare ground and rocks and also under rocks for endogaeic, epigaeic and other stationary invertebrates. The countdown timer was stopped when an area of suitable rocks for turning was encountered, and all turnable rocks within an area of approximately 4 m2 were turned over to reveal invertebrates hiding under the rocks. The rocks were then replaced in their original positions. The timer was started again when walking resumed. Another period of five minutes of search time was spent searching flowering plants for pollinators. Apart from bees, only insects that were feeding or sitting on the pollen-producing parts of the flowers were considered potential pollinators. A third period of five minutes search time was spent looking for low-flying insects, insects sitting on vegetation and insects disturbed from vegetation by the observer's feet and net. Specimens were collected for identification in the laboratory and in museums. Sampling was done between 06h25 and 11h45, to make the most of the generally clearer early morning weather. The plot data were used in the comparison of sites. The comparison is relative and indicates differences in species density; it is not a comparison of total species complement.
Statistical analysis
The true number of focal species on each plot was estimated from the sampling data in Estimates (Colwell 2005) using the Chao1 estimator with the classic setting if there were no singletons or doubletons, or with the bias-corrected setting otherwise, and with the default settings for the other parameters.
Analysis of the plots by species matrix was carried out using PC-Ord5 (McCune & Mefford 1999). McCune and Grace (2002) and the PC-Ord5 advisor wizard were used to select an appropriate data relativisation method and analysis type. The percentage of cells empty was 74.317%. The co-efficient of variation (CV) of plot totals was 88.48% and the CV of species totals was 240.30%. species abundances were relativised by maximum to counter the effect that a few species that were relatively abundant at the time of sampling would have on the analysis and because the sampling time was restricted to a short period, which led to most species being rare in the samples. After relativisation, no plots or species were recognised as outliers using the Sorensen (Bray-Curtis) distance measure and a cutoff of two standard deviations from the grand mean. A cluster analysis dendrogramme was produced using the Sorensen (Bray-Curtis) distance measure and the flexible beta (beta set to -0.25) group linkage method. Non-metric multidimensional scaling (NMDS) was carried out using the autopilot mode and the Sorensen (Bray-Curtis) distance measure (McCune & Grace 2002). A random starting configuration and 250 runs with real data and 250 runs with randomised data for comparison were utilised.
- The variables included in the environmental matrix in the NMDS are indicated in Table 1. The expectations were that:
- The further away the peaks from the Sentinel in straight-line distance, the greater the turnover of species and therefore the greater the difference in observed species complement.
- Species composition should be more similar between closer peaks than between peaks further apart.
- Peaks further away from the main massif were expected to have depauperate faunas compared to peaks closer to the main massif, owing to stochastic events that may have eliminated certain species that would not have recolonised the peak easily or rapidly from the main massif.
- The larger the area of the peak, the greater the number of species that would be recorded.
- Differences in extreme temperatures (represented by the means of daily minimum temperature for the coldest month, July, and the means of daily maximum temperature for the hottest month, January) or else in mean annual temperature would influence the distribution of species on the peaks (climate data obtained from Schulze 2007).
- Differences in mean annual precipitation between the peaks would cause differences in the faunal communities on the peaks.
- Differences in the altitude of the peaks are small and would not influence the composition of the faunal communities.
The six peaks were grouped into a northern pair (Sentinel and Eastern Buttress), a central pair (Inner and Outer Horns) and a southern pair (Dragon's Back and Cathkin).
Results
Invertebrates
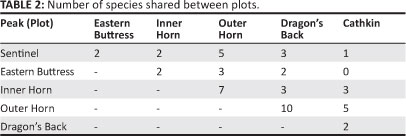
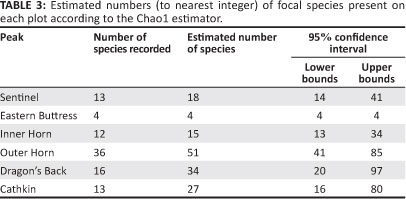
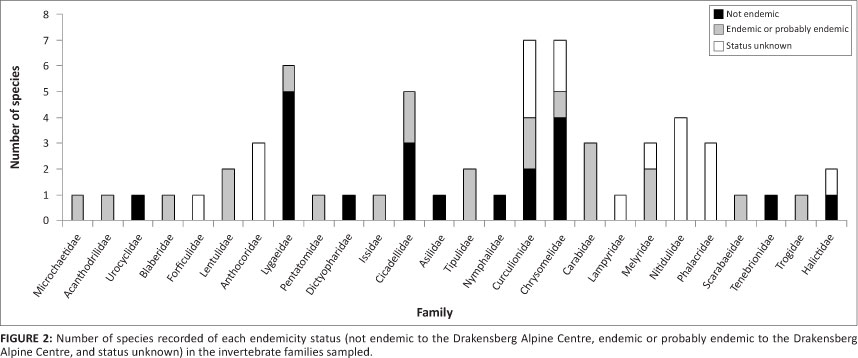
A total of 341 individuals of 61 species, comprising 26 families, of the focal taxa were recorded in all plots sampled (Appendix 1). The total of 61 species of invertebrates included two orders, two families and two species of Oligochaeta, a single order, single family and single species of Gastropoda and eight orders, 23 families and 58 species (95% of the total invertebrate species sampled) of Insecta. Within the focal groups sampled, between 18% and 36% of the total invertebrate species recorded are endemic to the DAC (known endemics and known + possible endemics, respectively; Appendix 1), although the proportion varied between families, ranging from 0% to 100% (Figure 2). Twenty-nine species of the total 61 were precisely identified (three of which were described after the survey), four were undescribed, and eight others were probably undescribed (Appendix 1). Many of the remaining species may also be undescribed. Eleven species are not known outside the DAC, with a further 11 possibly endemic to the DAC (Appendix 1; Figure 2). The Outer Horn had the most species (36) and the Eastern Buttress the fewest (4; Appendix 1). The Outer Horn and Dragon's Back peaks shared the most species (ten), followed by the Inner Horn and Outer Horn peaks with seven shared species (Table 2). Five species were shared between the Outer Horn and Cathkin and between the Outer Horn and Sentinel. The estimated true number of focal species for most plots was higher than that actually recorded (Table 3).
Vegetation and invertebrate association
The phytosociological survey done simultaneously with the invertebrate survey on the same inselbergs (Brand, Collins & Du Preez, in prep.) found the vegetation structured into four main communities consisting of shrubs, graminoids (grasses and sedges), forbs and geophytes. The first main vegetation community consists of shrubs and graminoids found on all peaks and consists of low shrubs (from 50 cm to 200 cm height) - Erica glyphra, Erica dominans, Helichrysum trilineatum and Anthospermum basuticum - plus tall bunch grass, Merxmullera drakensbergensis, and the sedge Scirpus falsus. The second main vegetation type consists mostly of high-altitude endemic grasses, Pentaschistis exerta, Pentaschistis basutorum and Festuca caprina. This community predominates on the Eastern Buttress and Cathkin peaks. The third community type is not well defined and consists of short, scattered forbs, geophytes and high-altitude, endemic grasses. The fourth main vegetation community is confined to wetlands or rocky seeps and consists of limited numbers of small, scattered, low geophytes, forbs and succulents most of which are obligate wetland plants, having a > 99% probability of occurring in wetlands (Tiner 1999). These habitats with their associated vegetation and invertebrates occur on all the inselbergs, portions of which were included in the invertebrate sampling plots (Figure 3a). As a result, no clear association between invertebrates and specific vegetation communities could be discerned.
Invertebrate data analysis
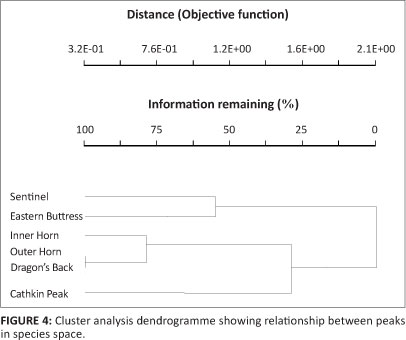
The cluster analysis indicated that the two northern peaks clustered separately from the Cathkin Peak cluster and the cluster formed by the other three peaks in terms of species composition, but that the closest peaks in straight-line distance did not necessarily cluster together the most closely (Figure 4, Table 1).
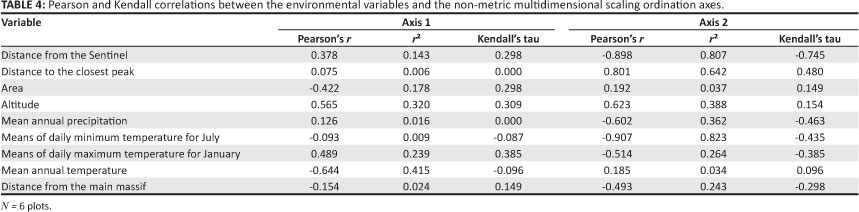
The preliminary runs of the NMDS ordination recommended a two-dimensional solution. The best two-dimensional solution had a final stress of 0.00 after 97 iterations. The Monte Carlo test result indicated that the final stress could not have been obtained by chance (p < 0.0478). The plot of stress versus iteration number for the final run was flat after iteration ten, indicating that a stable solution had been found. No good correlations between axis one of the NMDS and the environmental variables were obtained, whereas some good correlations between axis two and the environmental variables were evident (Table 4). The latter results indicated that the distance from the Sentinel and the mean daily minimum temperature for July had the highest correlations with the species data (Table 4), with distance to the closest peak having the next highest correlation. The correlations between the species data and the other environmental variables generally were low (Table 4).
Ethical considerations
All work was done under Ezemvelo KZN Wildlife Permit No. 2762/2004 (see the Acknowledgements section).
Discussion
The study set out to sample focal taxa on a number of inselbergs in the alpine region of KZN to give an indication of whether the DAC is a centre of endemism for invertebrates, whether invertebrate communities change from isolated peak to isolated peak, and possible reasons for any detected change. The inselbergs were chosen as study sites because of their relative isolation from the main Drakensberg massif, on the assumption that the plant and animal communities have been little changed over the past few millennia by anthropogenic pressures such as livestock grazing and altered fire regimes. Grazing by cattle and other domestic livestock is absent on the inselbergs and it is assumed that some uncontrolled fires started by humans do not reach the inselbergs because of their relative isolation. The main herbivores on the inselbergs are invertebrates and small mammals. Rodent faeces, burrows and cuttings in the grass were found on the Outer Horn and Cathkin peaks as well as small bones also suggestive of rodents (Brand 2006, submitted). However, this herbivory is likely to have a positive effect on the vegetation in contrast to that of medium-sized to large domestic livestock. Consequently, the vegetation can be regarded as being 'pristine' and not exhibiting the effects of over-grazing by cattle and sheep.
This study of the plant and invertebrate communities on the inselbergs of the alpine region of the uKhahlamba-Drakensberg Park World Heritage Site appears to be the first of its kind. The conservation context of the study is the planned increase in area under protection in the Maloti Drakensberg Transfrontier Project planning region (Maloti Drakensberg Transfrontier Project 2008).
Endemic invertebrates of the Drakensberg Alpine Centre
Between 18% and 36% of the invertebrate species recorded are endemic to the DAC, a higher percentage than that for angiosperms as a whole (Carbutt & Edwards 2006). This is suggestive that the DAC is also a centre of endemism for invertebrates, although much more field collecting and taxonomic work on the invertebrate fauna is required (Clark et al. 2011) before this suggestion can be substantiated. A firm conclusion on the exact percentage of the invertebrates recorded during this study that are endemic to the DAC is hampered by the fact that a large proportion of the species could not be identified. Only a proportion of the invertebrate taxa on the inselbergs (the focal taxa) could be sampled in the short time available. To compile for the DAC as comprehensive a list of insects and other focal invertebrates as that compiled for the angiosperms, so as to determine amongst other things the exact percentage of endemism for these invertebrates, will require much more intensive and extensive sampling in the various seasons throughout the DAC.
The number of undescribed species indicates that the invertebrate fauna of the DAC is still not well-described. Certain species, however, do appear to have restricted ranges even within the DAC (Appendix 1). The fact that some endemic microchaetid earthworms and wingless bugs have restricted distribution ranges is well known (Plisko 2003; Stiller 2009, 2010; Theron 1986). These species may only occur on certain peaks as a result of the spatial distribution of the environmental factors that influence the species' distributions and the relative immobility of the species. These environmental factors may include soil attributes such as pH, depth, and clay and water content, and vegetation attributes such as physiognomy and host-plant distributions (e.g. Edwards & Bohlen 1996 for earthworms). Species associated with grasses (because they or their larvae are grass-feeders or else they are associated with forbs growing in grassland) would be more likely to be found on the peaks with a greater proportion of the area under grasses, these being the Sentinel, Dragon's Back, and Cathkin (main plant communities one and two comprised of shrubs and grasses). Lepidopteran and hemipteran grass-feeders such as Pseudonympha gaika, Pravistylus interdiscus, and Drakensbergena bisulca (Pringle et al. 1994; Stiller 2009, 2010), and an orthopteran that feeds on leaves of bushes or forbs in grassland, Basutacris inflatifrons (Brown 1962; Figure 3d), tend to show this type of distribution (Appendix 1). Other possibilities are that specific plant species with which certain wingless species are associated may not occur on some inselbergs, or finally, some wingless species may not be present on certain peaks because they have become extinct on those peaks through the impact of environmental stochasticity.
Factors responsible for the diversity and composition of invertebrate communities on the inselbergs
Inselbergs are defined as isolated, freestanding topographic features, clearly visible in the landscape (Gröger & Barthlott 1996; Parmentier et al. 2006). They are composed either of igneous or sedimentary rock and have sheer, or near sheer sides (Groger & Barthlott 1996). They constitute sites of high biodiversity with large numbers of endemic taxa and represent relictual populations of previous episodes of climate change (Parmentier et al. 2006; Porembski et al. 1996). The six inselbergs surveyed are isolated from the escarpment by several hundred metres and form island-like archipelago strings of peaks with sheer basalt sides up to 1000 m high. They are all inaccessible to humans except by climbing or helicopter. The length of time the individual peaks have been separated could not be inferred from geological or geomorphological information, but King (1963) accounts for a retreat of the original Escarpment rim near the current day KZN coast of 161 km in 123 million years, which Killick (1990) reduces to an average rate of 1.3 mm per year.
The most important factors influencing invertebrate community composition on the inselbergs was distance from the Sentinel and mean minimum temperature for July, according to the NMDS analysis (Table 4). Distance from the Sentinel reflects how much further away in a south-easterly direction the next peak is from the Sentinel (Figure 1b). Turnover of species in this south-easterly direction is reflected. That the two northern peaks clustered together, the most southerly peak, Cathkin, formed its own cluster, and the other three peaks formed a cluster according to the cluster analysis (Figure 4) gives support to this contention. Mean annual rainfall tends to increase in this direction, as do the maximum and minimum temperature figures to a lesser extent (Table 1). Change in species identities and taxon richness occurs with increasing altitude in the Drakensberg mountains and, at least for some taxa, fewer species are encountered at higher altitudes (e.g. Armstrong & Van Hensbergen 1997a, 1997b; Samways 1989; Stuckenberg 1962). Pryke and Samways (2008, 2010) found that invertebrate species richness, abundance and composition changed with altitude on Table Mountain. Epigaeic and aerial invertebrates decreased in richness and abundance with increasing altitude but the converse was true for foliage invertebrates. Therefore, beta diversity might be expected to be lower at higher altitudes in the DAC. Only one species was recorded in all three clusters (Figure 4; Appendix 1). More intensive sampling may reveal that other species occur in all the clusters. Uys et al. (2009) determined high turnover values for flightless forest invertebrates at scales varying from a few hundred metres to tens of kilometres at lower altitudes in the same general region of the uKhahlamba-Drakensberg Park World Heritage Site as this study.
Mean minimum temperature for July (the coldest month) was found to influence invertebrate community composition. The temperature data were at a one minute by one minute scale that was possibly too large to give precise temperatures for each peak. The South African Weather Service has 17 stations in the Drakensberg, of which two, Sani Pass (2865 m) and Witsieshoek (2500 m, below the Sentinel), are at high altitude, yet still below the altitudes of the study sites (Nel 2007). The influence of low temperatures, frost, and low oxygen partial pressure on insects has been documented (Chapman 2002), and these play an appreciable part as limiting factors responsible for insect endemism and diversity at high altitude. Altitude and vegetation structure are two of the most important environmental variables that influence invertebrate diversity on Table Mountain (Pryke & Samways 2008, 2010). The texture and structure of Alpine vegetation, regardless of slope, aspect and soils, is dependent on the maximum and minimum temperatures (Körner 2003), which might thus influence the relationship between invertebrates and vegetation type.
Insects and other invertebrates, like lizards, are poikilotherms; their temperature depends largely on the amount of incident solar radiation and on the ambient temperature (Chapman 2002; Clusella-Trullas et al. 2008). Clusella-Trullas et al. (2008) found that skin reflectance was more strongly related to mean annual radiation than to mean annual temperature in their sample of 68 and 66 species of lizards, respectively. Dark beetles absorb more direct shortwave and visible radiation than light-coloured ones, which would enable them to use more limited amounts of sunlight to warm up and become active, providing wind is absent (e.g. Turner & Lombard 1990). Alpine and arctic insects are often dark, or have darker scales and wing undersides to take advantage of low ambient light and temperature levels (Chapman 2002). About 60% of the insect species sampled during the present study are black or very dark brown or grey, and may be darker than congeners at lower altitudes. The montane tail-wagger snail Sheldonia fuscicolor (Figure 3e), for example, is darker than its congeners at lower altitudes in KZN (Herbert & Kilburn 2004).
Insect taxa that have species known to be frost tolerant include Blattoidea, Hemiptera, Diptera, Lepidoptera, Coleoptera and Hymenoptera (Chapman 2002; cf. Appendix 1). Insects living in the DAC have to endure temperatures close to and below freezing at various times of the year, and especially in winter, and so are likely to be freeze tolerant (Sinclair & Chown 2005). Two species from the KZN part of the DAC, a cockroach (probably Perisphaeria guillarmodi; S. van Noort, pers. comm., 23 April 2012) and a ground beetle, were included in the study of Sinclair and Chown (2005). Both these species are moderately freeze-tolerant, with a lethal temperature lower than the temperature of crystallisation. The low temperature at which 50% mortality (LT50) of the cockroach was recorded was -9.7 °C and the LT50 for the ground beetle was -4.4 °C (Sinclair & Chown 2005). Therefore, many of the species recorded are likely to be adapted to the low solar radiation incidence and ambient temperatures experienced during the winter season and the frequently cloudy and wet summer period in the DAC (Killick 1978). The ability to endure low solar radiation and temperatures is likely a major factor that influences the composition of invertebrate communities in the DAC.
Expectations one, two, five and seven were the only ones supported or partially supported by the data (see Research methods and design: Statistical analysis; Figure 4; Tables 1, 2 & 4). Surface area of the inselbergs was not important in terms of structuring the invertebrate communities nor in terms of the number of species present (Tables 1 & 4). The Eastern Buttress yielded only four species from the focal groups, even though it was the second-largest peak, possibly as a result of the cool and damp conditions following the storm the evening before sampling, which may have inhibited some invertebrate activity and therefore detection of certain species. The Outer Horn and the Inner Horn were expected to have the most similar faunal community in terms of invertebrate species identities, because they were the pair of peaks closest together (Table 1). They shared seven species (as recorded during the plot counts), and this pair of peaks was ranked second in number of shared species.
One of the more important factors allowing for high biodiversity is terrain heterogeneity (Cowling & Lombard 2002), that is, mountains tend to be topographically complex with variations of soil, rocky slopes, and aspect and have more species than flat areas due to the greater number of ecological niches available. The relatively flat topography on the inselbergs (Figure 3b & 3c) does not allow for significant habitat differentiation and only four main vegetation communities (habitats) are found. Despite this, of the total 61 invertebrate taxa found, 11 are DAC endemics, with a further 11 possibly endemic to the DAC (Figure 2). This represents 36.1% endemic and possibly endemic species out of the total fauna sampled and is comparable with that found in a study of invertebrates on Table Mountain, Cape Town, where habitat heterogeneity was higher (total of 175 species, with 6 surface-endemic invertebrate species and 28 Cape Peninsula endemics, giving 16% endemicity; Pryke & Samways 2008, 2010). Endemicity varied from 0% to 100% amongst the various families recorded in the present study (Figure 2). Picker and Samways (1996) recorded percentage endemicity of between 5% and 67% for various faunal taxa on the Cape Peninsula in a far more comprehensive study.
Limitations of the study
The fact that a high percentage of invertebrate species were only represented by a single specimen (Appendix 1) and that the true numbers of focal species on the plots is likely to be higher than those recorded (Table 3) indicates that more time needs to be spent sampling on the peaks in all four of the main vegetation communities of the study plots to gain a fuller picture of the invertebrate communities and the associations between the invertebrates and the plant communities. This was not possible during the present study owing to logistical constraints, including the inaccessibility of the peaks, which could only be reached by helicopter and in good flying conditions, and the mountain weather conditions, which are very variable in early summer in the Drakensberg. The weather conditions, particularly with respect to the formation of clouds, can and did change rapidly at the sampling sites. In particular, the periodic overcast conditions that pertained during sampling may have suppressed insect activity to some extent and, therefore, detection of insects.
Further research
Further sampling to elucidate the impacts that unmanaged burning and livestock grazing have on the invertebrate fauna of the DAC would be valuable. A survey design that used paired plots, on the main Drakensberg massif that is subject to human activity, especially burning and livestock grazing, and on neighbouring isolated peaks that are not, or seldom, subject to these impacts, would be most appropriate. This may be difficult to achieve, as access to the peaks is conditional on sufficient budget for helicopter transport, and the prevailing weather conditions.
The results of the present study suggest that the focal invertebrate taxa for sampling should be groups with range-restricted DAC-endemic species, such as (other reasons in parentheses): earthworms (detritivores and soil engineers that are likely to be sensitive to soil water changes caused by trampling and excessive removal of vegetation and plant litter), wingless grasshoppers (herbivores that may be sensitive to changes in vegetation structure and composition), flightless bugs (herbivores that are likely to be sensitive to vegetation changes caused by excessive fire and livestock herbivory), crane flies (detritivores that are likely to be sensitive to changes in vegetation and surface water), ground beetles (predators of other invertebrates), darkling beetles (detritivores) and hair beetles (decomposers of vertebrate remains that may be sensitive to changes in abundance of mammals and birds caused by hunting pressure and changes in vegetation structure caused by altered fire and herbivory regimes). A relatively large proportion of insect species inhabiting high-altitude areas are wingless (e.g. Endrody-Younga 1989; Roff 1990). These insects are most likely to be vulnerable to large-scale changes in vegetation as their mobility is reduced compared to fully-winged species. The sampling methods used would have to be appropriate to these groups and the sampling would have to be intensive enough to discern changes in the abundance and identities of species occurring in the treatment area relative to the control.
The precise relationship between plant community structure and alpine invertebrate use of, dependence on, and insect pollination of plants is not clear from this survey and requires further investigation. A vegetation-based approach to insect conservation may be fruitful (e.g. Panzer & Schwartz 1998).
Biodiversity conservation
The diversity of invertebrates and plants found on the six inselbergs in the Drakensberg is exceptional, especially considering that 61 focal invertebrate species and 189 plant species were recorded on the 31.9 ha studied. The high number of endemic and possibly endemic invertebrate species (22), which constitutes about 36% of the invertebrate species sampled, coupled with the 112 endemic and near endemic plant species, adds further prominence to the unique faunal and floral value of this region of the DAC. The inselbergs represent refugia for plants and animals because these are sites relatively untouched by direct anthropogenic influence. Climate change is likely to start influencing the diversity of plant species, vegetation structure and invertebrate species on the inselbergs. However, this should not prevent these high-altitude, alpine areas from receiving statutory protection. Carbutt and Edwards (2006) calculated that only about 5.5% of the DAC is formally conserved, and about 97% of this conserved area occurs within the uKhahlamba-Drakensberg Park World Heritage Site in KwaZulu-Natal. The latter authors have already noted the need for the continued protection of the biota of the DAC. The present study provides evidence that it is not only the unique and threatened flora that needs continued protection and conservation, but also the relatively large proportion of invertebrate species that are endemic to the DAC. Conservation of more of the DAC should form the basis for inter-institution and international (Lesotho and South African) collaboration to conserve its unique plants and animals, and also to obtain a better understanding of this alpine ecosystem through research in various and distant areas of the DAC where anthropogenic impacts are minimal.
Conclusion
This study is the first to have sampled invertebrate taxa in a standardised way (timed counts on a 50 m by 50 m plot) on the top of each of six high inselbergs of the uKhahlamba-Drakensberg Park World Heritage Site, which are free of most anthropogenic impacts. The results suggest that, as for plants, the DAC is a centre of endemism for invertebrates, with a substantial number of undescribed species and species endemic to the DAC being recorded. More sampling both on the inselbergs and on the main massif is required to enable an accurate estimate of the proportion of invertebrate species that are endemic to the DAC to be made, as well as to give an indication of how various anthropogenic threats impact the invertebrate fauna. Invertebrate species turnover was shown to occur in a south-easterly direction. The results indicate that more of the latitudinal and longitudinal extent of the DAC should be given statutory protection, as there are likely to be invertebrate species endemic to the DAC that are not yet protected. This would be best achieved through the ongoing collaboration between South Africa and Lesotho through the Maloti-Drakensberg Transfrontier Park programme.
Acknowledgements
The National Geographic Committee for Research and Exploration supported the fieldwork (grant # 7920-05), as without these funds the study would not have been possible. We thank the following people for identifying specimens or for assisting with their identification: Danuta Plisko (earthworms), Michael Stiller (bugs), David Rider (Aegaleus), Dai Herbert (snails), Beth Grobbelaar (beetles), Riaan Stals (beetles), Ruth Muller (beetles) and James Harrison (beetles). We also thank Nunku Mkhize and Keenan Stears for setting specimens. Richard Lechmere-Oertel is thanked for providing the areas of the larger peaks calculated from the track logs of a GPS and for photographs, Debbie Jewitt for advice on using PC-Ord5 and comments on a previous version of the manuscript, and Heidi Snyman for assistance with the production of some of the figures. Specimens were collected under Ezemvelo KZN Wildlife Permit No. 2762/2004 and are deposited in the South African National Collection (SANC), the Ditsong National Museum of Natural History (TMSA), the Durban Natural Science Museum (DMSA) and the KwaZulu-Natal Museum (NMSA). The photographs in Figures 3a, 3b and 3c are courtesy of Richard Lechmere-Oertel of the Maloti-Drakensberg Transfrontier Park Project. Editorial comments by Ray Jansen of Tshwane University of Technology (TUT) and the comments of two anonymous reviewers were very useful and improved the manuscript.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this paper.
Authors' contributions
A.J.A. (Ezemvelo KZN Wildlife) was responsible for the sampling techniques, collection of field data, processing of specimens and deposition in museums, identification of specimens through collaboration with taxonomic experts, collation of ancillary data such as distribution ranges from specimens and the literature, analysis of the data, and writing up of the paper. R.F.B. (Tshwane University of Technology) was the project leader and developed the concept of sampling on the isolated peaks, secured the grant that enabled the fieldwork to be undertaken, contributed ideas as to how to further develop the paper, and contributed to the botanical and other sections of the paper.
References
Alexander, C.P., 1964, 'Diptera (Nematocera): Tanyderidae, Ptychopteridae, Tipulidae', in B. Hanstrom, P. Brink & G. Rudebeck (eds.), South African Animal Life, vol. 10, pp. 229-441, Swedish Natural Science Research Council, Stockholm. PMid:21540865 [ Links ]
Armstrong, A.J. & Van Hensbergen, H.J., 1997a, 'Evaluation of afforestable montane grasslands for wildlife conservation in the north-eastern Cape, South Africa', Biological Conservation 81, 179-190. PMid:17538161 [ Links ]
Armstrong, A.J. & Van Hensbergen, H.J., 1997b, 'A test of the applicability of a wildlife conservation evaluation method to an afforestable region of Mpumalanga', South African Journal of Wildlife Research 27, 73-77. PMid:17960012 [ Links ]
Armstrong, A.J. & Van Hensbergen, H.J., 1999, 'Identification of priority regions for animal conservation in afforestable montane grasslands of the northern Eastern Cape Province, South Africa', Biological Conservation 87, 93-103. PMid:18784101 [ Links ]
Armstrong, A.J., Van Hensbergen, H.J. & Geertsema, H., 1994, 'Evaluation of afforestable montane grasslands for wildlife conservation in the north-eastern Cape, South Africa. Part 1. Methods', South African Forestry Journal 171, 7-20. PMid:19197326 [ Links ]
Arnott, W.L., 2006, 'The effect of burning frequency on invertebrate and indigenous flowering forb diversity in a Drakensberg grassland ecosystem', MSc thesis, School of Biological and Conservation Sciences, University of KwaZulu-Natal, Pietermaritzburg. [ Links ]
Bazelet, C.S. & Samways, M.J., 2011, 'Relative importance of management vs. design for implementation of large-scale ecological networks', Landscape Ecology 26, 341-353. [ Links ]
Brand, R.F., 2006, 'The ecology and biogeography of pre and post Gondwana inselbergs, Drakensberg Alpine Centre, South Africa', Report to the National Geographic Society, Committee for Science and Exploration, Washington, DC. [ Links ]
Brand, R.F., submitted, 'A floristic, biogeographic and growth form assessment of Afroalpine inselberg vegetation in the Drakensberg Alpine Centre', South African Journal of Botany. PMid:8136946 [ Links ]
Brand, R.F., Collins, N. & Du Preez, P.J., in prep., 'A phytosociology survey and biogeographical description of inselberg vegetation in the Drakensberg Alpine Centre, South Africa'. [ Links ]
Brown, H.D., 1962, 'New and interesting grasshoppers from southern Africa - 3 (Othoptera: Acridoidea)', Journal of the Entomological Society of South Africa 25, 198-229. [ Links ]
Carbutt, C. & Edwards, T.J., 2004, 'The flora of the Drakensberg Alpine Centre', Edinburgh Journal of Botany 60, 581-607. [ Links ]
Carbutt, C. & Edwards, T.J., 2006, 'The endemic and near-endemic angiosperms of the Drakensberg Alpine Centre', South African Journal of Botany 72, 105-132. [ Links ]
Chapman, R.F., 2002, 'Thermal relations', in R.F. Chapman (ed.), The insects: Structure and function, pp. 509-530, Cambridge University Press, Cambridge. [ Links ]
Clark, V.R., Barker, N.P. & Mucina, L., 2011, 'The great escarpment of southern Africa: A new frontier for biodiversity exploration', Biodiversity and Conservation 20, 2543-2561. [ Links ]
Clusella-Trullas, S., Terblanche, J.S., Blackburn, T.M. & Chown, S.L., 2008, 'Testing the thermal melanism hypothesis: A macrophysiological approach', Functional Ecology 22, 232-238. [ Links ]
Colwell, R.K., 2005, EstimateS: Statistical estimation of species richness and shared species from samples, Version 7.5, User's Guide and application published, viewed 11 June 2012, from http://purl.oclc.org/estimates [ Links ]
Cowling, R.M. & Lombard, A.T., 2002, 'Heterogeneity, speciation/extinction history and climate: Explaining regional plant diversity patterns in the Cape Floristic Region', Diversity and Distributions 8, 163-179. [ Links ]
Eardley, C.D., 1988, 'A revision of the genus Lithurge Latreille (Hymenoptera: Megachilidae) of sub-Saharan Africa', Journal of the Entomological Society of South Africa 51, 251-263. [ Links ]
Eardley, C., Kuhlmann, M. & Pauly, A., 2010, 'The bee genera and subgenera of sub-Saharan Africa', Abc Taxa 7, 1-139. [ Links ]
Edwards, C.A. & Bohlen, P.J., 1996, Biology and ecology of earthworms, Chapman & Hall, London. [ Links ]
Endrödy-Younga, S., 1989, 'The evolution of alternative life styles in Coleoptera', in M.N. Bruton (ed.), Alternative life-history styles of animals, pp. 317-327, Kluwer Academic Publishers, Dordrecht. [ Links ]
Fennah, R.G., 1967, 'New and little-known Fulgoroidea from South Africa (Homoptera)', Annals of the Natal Museum 18, 655-714. [ Links ]
Gröger, A. & Barthlott, W., 1996, 'Biogeography and diversity of the inselbergs (Laja) vegetation of southern Venezuela', Biodiversity Letters 3, 165-179. [ Links ]
Hamer, M. & Slotow, R., 2007, 'An assessment and ecological interpretation of the terrestrial invertebrate fauna of the South African portion of the Maloti-Drakensberg bioregion', Unpublished Report to the Maloti-Drakensberg Transfrontier Park project, Inland Invertebrate Initiative, University of KwaZulu-Natal, Pietermaritzburg. [ Links ]
Herbert, D. & Kilburn, D., 2004, Field guide to the land snails and slugs of eastern South Africa, Natal Museum, Pietermaritzburg. [ Links ]
Iwan, D., 2001, 'Two new species of Bantodemus Koch, 1955 (Coleoptera: Tenebrionidae: Platynotini) from South Africa', Annales Zoologici (Warszawa) 51, 505-510. [ Links ]
Killick, D.J.B., 1978, 'The Afro-alpine Region', in M.J.A. Werger (ed.), Biogeography and ecology of southern Africa, pp. 515-560, Junk, The Hague. [ Links ]
Killick, D.J.B., 1990, A field guide to the flora of the Natal Drakensberg, Jonathan Ball and Ad. Donker, Johannesburg. [ Links ]
King, L.C., 1963, South African scenery: A textbook of geomorphology, Oliver and Boyd, Edinburgh. [ Links ]
Körner, C., 2003, Alpine plant life: Functional plant ecology of high mountain ecosystems, Springer-Verlag, Berlin. [ Links ]
Londt, J.G.H., 1981, 'Afrotropical Asilidae (Diptera) 5: The genus Dasophrys Loew, 1858 (= Hobbyus Bromley, 1952) (Asilinae: Asilini)', Annals of the Natal Museum 24, 635-699. [ Links ]
Lovell, S.J., Hamer, M.L., Slotow, R.H. & Herbert, D., 2010, 'Assessment of sampling approaches for a multi-taxa invertebrate survey in a South African savanna-mosaic ecosystem', Austral Ecology 35, 357-370. [ Links ]
Maloti Drakensberg Transfrontier Project, 2008, 'Conservation and development strategy (2008-2028) for the Maloti Drakensberg Transfrontier Conservation Area', Maloti Drakensberg Transfrontier Project South Africa, Howick. [ Links ]
Marshall, G.A.K., 1904, 'A monograph on the Coleoptera of the genus Hipporrhinus, Schh. (Curculionidae)', Proceedings of the Zoological Society of London, 6-141. [ Links ]
McCune, B. & Grace, J.B., 2002, Analysis of ecological communities, MjM Software Design, Gleneden Beach, Oregon. [ Links ]
McCune, B. & Mefford, M.J., 1999, PC-ORD: Multivariate analysis of ecological data, Version 4, MjM Software Design, Gleneden Beach, Oregon. [ Links ]
Nel, W., 2007, 'On the climate of the Drakensburg: Rainfall and surface-temperature attributes, and associated geomorphic effects', PhD dissertation, Department of Geography, Geo-Informatics and Meteorology, University of Pretoria, Pretoria. [ Links ]
Panzer, R. & Schwartz, M.W., 1998, 'Effectiveness of a vegetation-based approach to insect conservation', Conservation Biology 12, 693-702. [ Links ]
Parmentier, I., Oumorou, M., Porembski, S., Lejoly, J. & Decocq, G., 2006, 'Ecology, distribution, and classification of xeric monocotyledonous mats on inselbergs in West Africa and Atlantic central Africa', Phytocoenology 36, 547-564. [ Links ]
Picker, M.D. & Samways, M.J., 1996, 'Faunal diversity and endemicity of the Cape Peninsula, South Africa - A first assessment', Biodiversity and Conservation 5, 591-606. [ Links ]
Plisko, J.D., 1994, 'New and little-known species of Proandricus Plisko, 1992 from Lesotho and Transkei (southern Africa) (Oligochaeta: Microchaetidae)', Annals of the Natal Museum 35, 229-235. [ Links ]
Plisko, J.D., 2003, 'Eleven new South African earthworms (Oligochaeta: Microchaetidae) with new information on some known species, and an inventory of the microchaetids of KwaZulu-Natal', African Invertebrates 44, 279-325. [ Links ]
Porembski, S., Szarzynske, J., Mund, J-P. & Barthlott, W., 1996, 'Biodiversity and vegetation of small-size inselbergs in a West African rain forest (Taï, Ivory Coast)', Journal of Biogeography 23, 47-55. [ Links ]
Princis, K., 1963, 'Blattariae', in B. Hanstrom, P. Brinck & G. Rudebeck (eds.), South African animal life, vol. 9, pp. 9-318, Swedish Natural Science Research Council, Stockholm. [ Links ]
Pringle, E.L.L, Henning, G.A. & Ball, J.B. (eds.), 1994, Pennington's butterflies of southern Africa, Struik, Cape Town. [ Links ]
Pryke, J.S. & Samways, M.J., 2008, 'Conservation of invertebrate biodiversity on a mountain in a global biodiversity hotspot, Cape Floral Region', Biodiversity and Conservation 17, 3027-3043. [ Links ]
Pryke, J.S. & Samways, M.J., 2010, 'Significant variables for the conservation of mountain invertebrates', Journal of Insect Conservation 14, 247-256. [ Links ]
Pryke, J.S. & Samways, M.J., 2012, 'Ecological networks act as extensions of protected areas for arthropod biodiversity conservation', Journal of Applied Ecology 49, 591-600. [ Links ]
Pryke, S.R. & Samways, M.J., 2001, 'Width of grassland linkages for the conservation of butterflies in South African afforested areas', Biological Conservation 101, 85-96. [ Links ]
Pryke, S.R. & Samways, M.J., 2003, 'Quality of remnant indigenous grassland linkages for adult butterflies (Lepidoptera) in an afforested African landscape', Biodiversity and Conservation 12, 1985-2004. [ Links ]
Robertson, H.G. & Van Noort, S., 2003, 'Effects of agricultural practices on ants and other Hymenoptera in four farming areas of South Africa', Unpublished Conservation Farming Project Report to the National Botanical Institute, Pretoria. [ Links ]
Roff, D.A., 1990, 'The evolution of flightlessness in insects', Ecological Monographs 60, 389-421. [ Links ]
Samways, M.J., 1989, 'Taxon turnover in Odonata across a 3000 m altitudinal gradient in southern Africa', Odonatologica 18, 263-274. [ Links ]
Schulze, R.E. (ed.), 2007, 'South African atlas of climatology and agrohydrology', WRC Report 1489/1/06, Water Research Commission, Pretoria. [ Links ]
Sinclair, B.J. & Chown, S.L., 2005, 'Climatic variability and hemispheric differences in insect cold tolerance: Support from southern Africa', Functional Ecology 19, 214-221. [ Links ]
Slater, J.A., 1964, 'Hemiptera (Heteroptera). Lygaeidae', in B. Hanstrom, P. Brinck & G. Rudebeck (eds.), South African animal life, vol. 10, pp. 15-228, Swedish Natural Science Research Council, Stockholm. [ Links ]
Stiller, M., 2009, 'A revision of the South African leafhopper genus Drakensbergena Linnavuori (Hemiptera: Cicadellidae: Drakensbergeninae)', Zootaxa 2132, 1-39. [ Links ]
Stiller, M., 2010, 'Revision of the southern African leafhopper genus Pravistylus (Hemiptera: Cicadellidae: Deltocephalinae)', Zootaxa 2468, 1-81. [ Links ]
Straneo, S.L., 1958, 'Carabidae Pterostichinae', in B. Hanstrom, P. Brinck & G. Rudebeck (eds.), South African animal life, vol. 5, pp. 9-318, Swedish Natural Science Research Council, Stockholm. [ Links ]
Stuckenberg, B.R., 1962, 'The distribution of the montane palaeogenic element in the South African invertebrate fauna', Annals of the Cape Provincial Museums 2, 190-205. [ Links ]
Synave, H., 1956, 'Contribution a l'étude des Issidae Africains (Homoptera - Fulgoroidea)', Bulletin de Institute royal des Sciences naturelles de Belgique 32, 1-22. [ Links ]
Theron, J.G., 1986, 'New genera and species of southern African Coelidiinae (Homoptera: Cicadellidae), with description of the new Tribe Equeefini', Phytophylactica 18, 153-163. [ Links ]
Tiner, R.W., 1999, Wetland indicators: A guide to wetland identification, delineation, classification and mapping, Lewis Publishers, Washington, D.C. [ Links ]
Turner, J.S. & Lombard, A.T., 1990, 'Body color and body temperature in white and black Namib desert beetles', Journal of Arid Environments 19, 303-315. [ Links ]
Uys, R., 2006, 'Fire effects on the fauna and flora of the Maloti-Drakensberg bioregion: A review', Unpublished report, Maloti-Drakensberg Transfrontier Project, Ezemvelo KZN Wildlife, Pietermaritzburg. [ Links ]
Uys, C., Hamer, M. & Slotow, R., 2006, 'Effect of burn area on invertebrate recolonization in grasslands in the Drakensberg, South Africa', African Zoology 41, 51-65. [ Links ]
Uys, C., Hamer, M. & Slotow, R., 2009, 'Turnover in flightless invertebrate species composition over different spatial scales in Afrotemperate forest in the Drakensberg, South Africa', African Journal of Ecology 47, 341-351. [ Links ]
Uys, C., Hamer, M. & Slotow, R., 2010, 'Step process for selecting and testing surrogates and indicators of Afrotemperate forest invertebrate diversity', PLoS ONE 5(2), e9100. PMid:20161757 [ Links ]
Van Wyk, A.E. & Smith G.F., 2001, Regions of floristic endemism in southern Africa: A review with emphasis on succulents, Umdaus Press, Hatfield. [ Links ]
Wildy, E.J., 2003, 'Effect of different land use practices on invertebrate diversity in Underberg (KwaZulu-Natal, South Africa)', Unpublished Conservation Farming Project Report to the National Botanical Institute, Pretoria. [ Links ]
Wittmer, Von W., 1949, 'Neue Malacodermata aus der Sammlung des British Museum, London (2. Beitrag zur Kenntnis der Malacodermata Afrikas)', Annals and Magazine of Natural History Series 12 2, 746-764. [ Links ]
 Correspondence to:
Correspondence to:
Adrian Armstrong
PO Box 13053, Cascades 3202, South Africa
Email: aarmstrong@kznwildlife.com
Received: 30 Apr. 2012
Accepted: 09 July 2012
Published: 30 Oct. 2012
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}