










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.54 n.1 Pretoria Jan. 2012
ORIGINAL RESEARCH
Phytosociology of the farm Haribes in the Nama-Karoo biome of southern Namibia
Ben J. StrohbachI; Willem J. JankowitzII
INational Botanical Research Institute, Windhoek, Namibia
IISchool of Natural Resources and Tourism, Polytechnic of Namibia, Namibia
ABSTRACT
Limited historic vegetation data (prior to the 1980s) are available for Namibia. Finding such historic data at Haribes prompted a follow-up survey of the vegetation. We present a classification of the recent data in this paper as a first step towards comparing the two data sets. Six new associations (three with two subassociations each) are formally described. The landscape at Haribes is dominated by a pan with surrounding hummock dunes. The pan supports the Lycio cinereum - Salsoletum, whilst on the hummock dunes the Salsolo -Tetragonietum schenckii can be found. The surrounding plains and escarpment can be divided into three landforms: the torras with the Monsonio umbellatae - Boscietum foetidae, the ranteveld with the Acacio senegal - Catophractetum alexandri (and two subassociations) and calcrete ridges with the Zygophylo pubescentis - Leucosphaeretum bainesii. Dry river beds on the farm support two subassociations of Anthephoro pubescentis - Ziziphodetum mucronatae. The area covered by each dominant landform has been calculated after being mapped. The composition and diversity of the associations are briefly compared to other known vegetation descriptions within the Nama-Karoo. Since November 2011, Haribes has been used as a resettlement farm. This may result in the overutilisation of the limited grazing resources, to the extent that the present, fairly dense Acacio senegal - Catophractetum alexandri of the ranteveld is feared to become degraded to resemble the Monsonio umbellatae - Boscietum foetidae of the torras.
CONSERVATION IMPLICATIONS: This paper describes six plant associations of the Nama-Karoo biome in arid southern Namibia. The information presented forms a baseline description, which can be used for future monitoring of the vegetation under altered land use.
Introduction
Vegetation in Namibia has been described only on a few attempts in the past (Burke & Strohbach 2000; Strohbach & Jürgens 2010). To date, the only concise vegetation map of Namibia was produced in the 1970s, based on field observations rather than actual surveys (Giess 1971, 1998), and depicts broad categories at biome rather than landscape level. Later attempts to map the vegetation of Namibia had to rely on partial, often incomplete, regional updates used in conjunction with the Giess vegetation map to fill in gaps (Mendelsohn et al. 2002). Grazing capacity maps dating from the 1970s (Departement Landbou Tegniese Dienste 1979) are used for land-use planning, including the ongoing Land Reform Programme of the Government of Namibia. However, these maps are regarded as outdated and in urgent need of revision (Espach 2006; Espach, Lubbe & Ganzin 2006; Lubbe 2005). Current efforts to upgrade the grazing capacity estimation with the aid of satellite images require extensive ground surveys, including surveys of the structure and composition of vegetation (Espach et al. 2006; Espach, Lubbe & Ganzin 2010). With this in mind, but also as basis for a biodiversity inventory, the Vegetation Survey of Namibia project was initiated (Strohbach 2001).
Prof. O.H. Volk was one of only a few vegetation scientists active in Namibia before the 1980s. During 1956, he conducted extensive vegetation surveys on selected farms in the former South West Africa. Unfortunately, only one of these data sets had been analysed and formally published (Volk & Leippert 1971); the others that could be retrieved were all in the form of raw data sets with brief comments on the vegetation, generally held in farm archives. Nonetheless, these data sets have proved to be of immense value in reconstructing vegetation changes over the past five decades of land use, especially with regard to the effects of land use and/or global climate change. One of these data sets, surveyed in April 1956, was from the farm Haribes.
This study was initiated to provide data on the current vegetation, for later comparison with Volk's historic data. The aim of this paper, however, is to classify and describe the present vegetation of the farm Haribes as an attempt towards filling the immense gap in vegetation data in Namibia (Strohbach & Jürgens 2010). In a subsequent paper, we intend to compare the present vegetation with that documented in 1956.
Study area
Haribes, in the Mariental district of the Hardap region in southern Namibia (Figure 1), belonged to the Neue Haribes Company until 2010. In 1956 the farm covered 77 500 ha, but has been reduced to its present 50 100 ha through a process of selling off blocks of land. The present survey was conducted across blocks 019/1, 019/REM, 020, 672 and 673, which total 55 487 ha. Initially, Haribes was used for extensive commercial Karakul sheep farming. The most important land-use change took place in 1992, when all the sheep and goats were sold and replaced by cattle as the only domesticated commercial species. As a result of the selective utilisation of the grass layer, woody species, especially Catophractes alexandri, Rhigozum trichotomum and Acacia nebrownii, increased in abundance, resulting in a phenomenon commonly referred to as 'bush encroachment' (De Klerk 2004). A wide variety of game, often in large numbers, are found on the farm. Most numerous are springbok and oryx. There are also many bird species, including many ostriches. The farm was sold in May 2010 to the Government of Namibia as part of its land reform and resettlement scheme (Cloete 2010a; Pretorius 2010), with the first 13 families receiving their plots in November 2010 (Cloete 2010b; Kleinhans 2010).
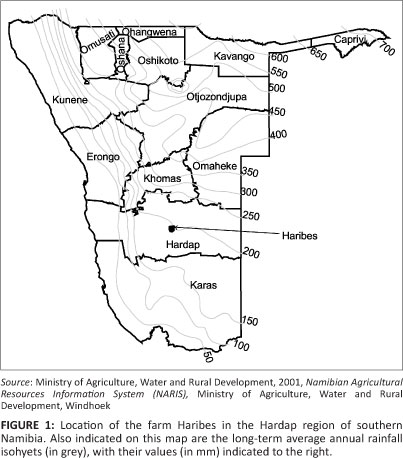
Haribes is situated approximately 30 km west of Mariental and southwest of the Hardap Dam Nature Reserve within the Central Plateau 4-8 agro-ecological zone (De Pauw et al. 1998). This zone is described as a strongly dissected plain on Karoo rocks. The average rainfall is about 200 mm per year, with a coefficient of variation between 50% and 60% (Botha 1996). Temperatures rise to above 36 °C in summer, with minimum temperatures of between 2 °C and 4 °C in winter (Mendelsohn et al. 2002).
The geomorphology can be described as a plateau broken up by many dry washes and some bigger ephemeral rivers that are responsible for the drainage of the area [a dissected plain according to the terminology of the Digital Soils and Terrain Database (SOTER) (FAO 1995; Oldeman & Van Engelen 1993]. The Riet River drains into a big pan surrounding the farmhouse complex. In and around the pan several well-developed dune hummocks have formed over time. The farm lies between 1120 m and 1320 m above sea level. The geological strata belong to the Fish River subgroup of the Nama sediments. The eastern part of the farm is situated on the Gross Aub Formation dominated by red shale and sandstone, whilst the larger, western part of the farm forms part of the older Nababis Formation with red sandstone and shales (Geological Survey 1980). The soils in the region (at a scale of 1:1 000 000) are classified as eutric leptosols, which means that the soils are fertile but generally very shallow (ICC, MAWRD & AECI 2000; Mendelsohn et al. 2002).
The farm falls within the Nama-Karoo biome (Irish 1994) and its vegetation is classified as Dwarf shrub savanna (Giess 1971, 1998). This can broadly be described as open, short-to-low shrublands dominated by C. alexandri, R. trichotomum, Boscia foetida and several dwarf shrubs. The grass layer, which includes species such as Stipagrostis uniplumis, Eragrostis nindensis, Enneapogon cenchroides and Schmidtia kalahariensis, is relatively sparse. Trees and larger shrubs, including species such as A. nebrownii, Acacia karroo, Euclea pseudebenus and Ziziphus mucronata, are largely confined to dry washes and rivers.
Research method
A total of 177 plots were sampled between March and May 2005. Below average rain (c. 150 mm, D. Poggenpoel, pers. comm., May 2005) had fallen that season, with the first substantial rains falling only in March.
To compare the present composition with that documented by Volk in 1956, we made exhaustive attempts to relocate his original sampling sites according to route descriptions, odometer distance readings (in miles) and old farm maps. Although this information greatly eased the task of relocation, the exact position of those plots could not be determined. Additional vegetation types were sampled using a stratified sampling approach to achieve a good average for the main vegetation types. For the purpose of stratification, a satellite map of the farm (based on Landsat 7 ETM scene path 177 row 077, 26 March 2001) was used as base map. Relatively homogenous areas were delineated as derived from segmentations calculated by Definiens 5 (Definiens 2006). Occasional azonal vegetation types (especially small vleis and related features) were also sampled.
Surveying followed the Braun-Blanquet concept as applied during the Vegetation Survey of Namibia project (Strohbach 2001). For each 20 m x 50 m plot, all species were recorded, as well as their typical growth form and average height. Abundance was estimated according to the modified Braun-Blanquet scale (Mueller-Dombois & Ellenberg 1974). In addition, the position of the plot and environmental parameters such as altitude, aspect, slope, topography, stone cover, erosion and disturbances were noted. Owing to time constraints, no soil samples were taken. Instead, available soil data (ICC et al. 2000; Mendelsohn et al. 2002) were interpolated for the various vegetation associations described. Unknown plant species were collected and identified at the National Herbarium of Namibia. The relevé data were captured in the TurboVeg database (Hennekens & Schaminée 2001).
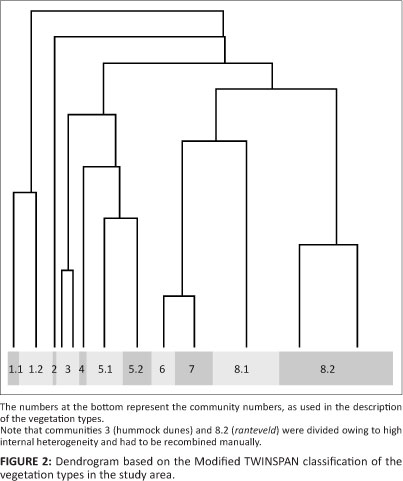
The data were classified with JUICE (Tichý 2002), using the Modified TWINSPAN classification procedure (Rolecek et al. 2009). The pseudospecies cut level was set to presence/ absence, and total inertia was selected as heterogeneity measure. The classification process concluded after 14 divisions. At this stage, two communities (hummock dunes and the ranteveld) were already subdivided owing to high internal heterogeneity and had to be recombined manually (Figure 2). The classification result was ordered as a table using the phi coefficient fidelity measure (Chytrý et al. 2002) as sorting criterion.
The classification results were confirmed with an ordination using Detrended Correspondence Analysis (DCA) (Hill & Gauch 1980), prepared with PC-Ord 5 (McCune, Grace & Urban 2002). Two iterations were performed, with the first excluding only two wetland relevés (as outliers) whilst the second also excluded the pans and associated hummock dunes. Although DCA has been criticised in recent years for 'detrending' the arch effect, this very effect proved useful in illustrating the relationships between the various vegetation associations, as Reciprocal Averaging (RA) depicted only a severe arch effect. The DCA ordination diagram was also used to depict habitat relations, which could often only be overlaid as categorical data.
Mapping was performed using a Landsat 7 ETM satellite image (path 177, row 077; 26 March 2001). In addition, a digital terrain model derived from Shuttle Radar Topography Mission (SRTM) data (available from http://srtm.csi.cgiar.org/) (Jarvis et al. 2008) was used to elucidate the topography. The satellite image was clipped to an area slightly larger than the study area before further processing. This clip was imported into the Definiens software package (Definiens 2006) and segmented into relatively homogenous areas. The segments were classified using the classified sample sites as ground truth data. In a final step, the resulting shape file was clipped according to the farm boundaries and areas for each landscape calculated using the IDRISI Andes software package (Eastman 2006).
Results
A total of 218 species, representing 42 families, were observed on the 177 relevés. Poaceae was the most abundant familiy, represented by the most species (54), followed by the Fabaceae (27 species), Asteraceae (16 species) and the Aizoaceae (13 species).
The classification of these relevés revealed eight communities, of which three were divided into subcommunities. The phytosociological table is presented in an online appendix 1, whereas online appendix 2 is a synoptic table. Figure 2 depicts a dendrogram of the classification result. The ordination diagrams of the DCA are shown in Figure 3a (all data except for two outlier relevés) and Figure 3b (also excluding the relevés representing pans and associated hummock dunes). In Figure 3c, the lithology (i.e. the rock types which gave rise to the soils) has been imposed onto the DCA ordination diagram of Figure 3b to illustrate the habitat relationships between vegetation communities.
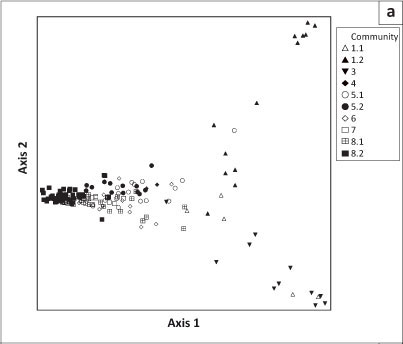
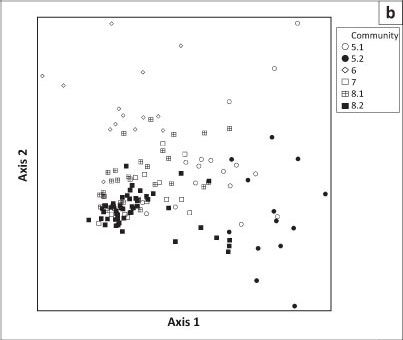
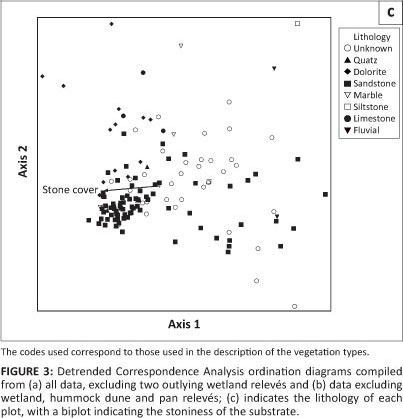
The pan and hummock dune communities form a well-defined continuum to the right of the ordination diagram in Figure 3a, indicating most likely a combined salinity and soil moisture gradient from the pan centre (community 1.2; upper right corner) to the surrounding hummock dunes (community 3; lower right corner). In contrast, the remaining communities (plains, ranteveld and rivers) are compressed on the left of the main axis. Splitting out the relevés representing the pan and hummock dune communities resulted in a continuum spread between the stony ranteveld, plain and torra communities as well as the rivers (Figure 3b). The limited habitat data available indicate that this is a gradient of increased stoniness towards the left of the main gradient (Figure 3c), representing the ranteveld. Together with the lithology (i.e. the rock type from which the substrate is derived), the stoniness forms the main ecological gradient in this environment.
Description of the plant associations
We formally describe six of the observed communities as associations, three with subassociations, following the International Code of Phytosociological Nomenclature (Weber, Moravec & Theurillat 2000). The two wetland communities, however, are described only informally, as the number of samples taken was too small for proper description.
1. Lycio cinereum - Salsoletum ass. nov.
The typicum for this association is relevé 2514, sampled at 24°35'36"S, 17°32'58"E on 09 March 2005 (Figure 4b). The association is represented by 21 relevés and characterised by the presence of Salsola spp. dwarf shrubs, which also dominate the vegetation. The structure is a short, open shrubland (sensu Edwards 1983), with an average shrub cover of 6%. Total vegetation cover barely reaches 10%. In total, 58 species have been observed on these pans, with an average species density of 12 per 1000 m2.
The Lycio cinereum - Salsoletum is typical of the large Haribes pan on the farm. The soils originate from weathered sandstones and some calcretes consisting of fine material which is very prone to capping and surface sealing (Figure 4c). The cation levels are probably elevated to form a halophitic environment, but not sufficiently so to render the pan a salt pan (M. Coetzee, pers.comm., March 2010).
Two subassociations can be distinguished, as described below.
1.1 The Zygophylo simplicis - Salsoletosum subass. nov.
The typicum for this subassociation is relevé 2518, sampled at 24°37'53"S, 17°32'31"E on 09 March 2005 (Figure 4a). Only five relevés represent this subassociation. The occurrence of Tetragonia schenkii, Trianthema sp. and Salsola spp. is characteristic and S. uniplumis var. uniplumis, S. kalahariensis, Boerhavia repens and A. nebrownii also occur constantly.
Along the outer parts of the pan and on the floodplains to the pan the vegetation is better developed, forming a tall, moderately closed shrubland (Figure 5). Few perennial herbaceous species occur here; this layer is dominated by ephemerals. The species density is on average 14 species per 1000 m2 and in total 39 species have been observed here.
1.2 The Lycio cinereum - Salsoletosum subass. nov.
This subassociation forms the typicum for the Lycio cinereum - Salsoletum. It is represented by 16 relevés and characterised by near-monospecific stands of Salsola shrubs. Other species that occur here are mostly of ephemeral nature. The shrubs are on average only 0.5 m - 1 m high and reach a cover of only about 6%, thus forming a short, open shrubland (Figure 5). On average, only five species occur per 1000 m2, with a total of 31 species being observed on these pans.
2. The Eragrostis rotifer - Leptochloa fusca community
This community was found once in relevé 2599 from a sample on a deeper vlei, which is regularly inundated. This is indicated by the presence of the geophytic hydrophyte Aponogeton cf. desertorum and Eragrostis rotifer, also generally associated with water-logged soils (Figure 4d). These uncommon vleis form in relatively small depressions, mostly on top of the plateau area in pans that range from 20 m to 50 m in diameter. The pans are surrounded by sandstone stones varying in size between 5 cm and 80 cm. The reddish colour indicates that the soil is most probably relatively high in nutrient content (M. Coetzee, pers. comm., March 2010).
3. Salsolo - Tetragonietum schenckii ass. nov.
The typicum for this association is relevé 2501, sampled at 24°37'00"S, 17°31'01"E on 08 March 2005 (Figure 4e). Eleven relevés were classified within this association. The association is characterised by the presence of the endemic shrub T. schenckii. Constant species include S. uniplumis var. uniplumis, Stipagrostis ciliata, Salsola spp. and R. trichotomum. The association is dominated by T. schenckii. The typical structure is a tall, moderately closed shrubland (Figure 5), with the shrubs reaching a height of between 1 m and 1.5 m. On average 11 species are found per 1000 m2, with a total diversity of 56 species.
The Salsolo - Tetragonietum schenckii is found on hummock dunes probably originating from a deflation process during the formation of the Haribes pan, similar to that described for the Koës pan and other depressions in the Kalahari (Bullard & Nash 2000; Lancaster 1986; Shaw 1988; Shaw & Thomas 1997). The dunes vary in length, generally between 30 m and 50 m. The slopes of the dunes are often quite steep. The occurence of Salsola spp. on these dunes indicate a high base status, which, in turn, supports the hypothesis that the dunes are the result of the deflation process of the pan.
4. The Cullen obtusifolia - Leptochloa fusca community
This community was identified in three relevés within the larger Haribes pan ecosystem. The presence of Leptochloa fusca and Indigofera alternans indicates water-logging at these vleis, whilst the absence of E. rotifer and Aponogeton sp. and the presence of the flat-growing therophytes Cullen obtusifolia and Platycarpha carlinoides indicate a drier environment than occurs in the deep vleis of the E. rotifer - Leptochloa fusca community. Typically for these vleis, A. nebrownii is dominant along the fringes (Figure 4f). These pans are small (50 m - 80 m in diameter) and very shallow. The high clay content of the soil allows the water to remain in the pan for approximately 2-3 weeks. In most cases, these pans are surrounded by reddish gravel, but no larger stones.
There are very few of these small pans that differ greatly from the big pans on the farm. Some are relatively small depressions, mostly on top of the plateau area, but there are also small pans below the plateau, especially near the old road along the tar road between Mariental and Maltahöhe. These depressions allow water to accumulate, resulting in microhabitats with a wetland character.
5. Anthephoro pubescentis -Ziziphodetum mucronatae ass. nov.
The typicum for this association is relevé 2649, sampled at 24°38'17"S, 17°28'12"E on 16 May 2005 (Figure 4h). A total of 30 relevés have been classified within this association. It is characterised by the diagnostic species Z. mucronata, Ocimum americanum var. americanum, Aristida meridionalis, Grewia flava, Anthephora pubescens, Monechma divaricatum and Melinis repens. These are constantly associated with the species S. uniplumis var. uniplumis, S. kalahariensis, Lycium cinereum, E. cenchroides, Gisekia africana, A. nebrownii, Eragrostis porosa, R. trichotomum and B. foetida. The structure is a low open woodland (sensu Edwards 1983), dominated by trees between 4 m and 6 m high that attain a cover of about 2%. Shrubs attain a cover of between 5% and 10%, at a height of about 1 m to 2 m. A total of 130 species have been observed in this association.
The Anthephoro pubescentis - Ziziphodetum mucronatae represents the riverine woodlands (sensu Giess 1998). There are two types of drainage system on the farm, with dry washes, which contain water only for very short periods during rain storms, being most common. Geomorphologically they can be classified as very young river systems. In many cases the slope is relatively steep and the run-off very fast. They occur mostly on the plateau or at the edge of the plateau. Towards the centre of the farm, more mature rivers, like the Riet, Holz and Lewer, are responsible for drainage. There is a big dam near the farmhouse, where the water of these rivers is captured. These two different habitats support two different vegetation subassociations (5.1 and 5.2).
Although the vegetation of the riverbed and the riverbank differ, they were sampled as one for the purposes of this study and are thus included in the same plant association.
5.1 Ehretrio albae - Ziziphodetosum mucronatae subass. nov.
The typicum of this subassociation is relevé 2574, sampled at 24°32'34"S, 17°32'45"E on 11 May 2005 (Figure 4g). A total of 17 relevés belong to this subassociation. There are no diagnostic species for this association, but the following are constantly present: S. uniplumis var. uniplumis, S. kalahariensis, L. cinereum, A. nebrownii, R. trichotomum, Limeum sulcatum, G. africana, B. foetida, E. cenchroides, G. flava, E. porosa and B. repens. The structure is a low, open woodland (Figure 5), with the trees only 4 m - 5 m tall and shrubs reaching height of 1 m - 2 m. In this subassociation, 99 species have been recorded; on average 21 species occur per 1000 m2.
The Ehretrio albae - Ziziphodetosum mucronatae occurs along the dry washes. Their slopes vary from steep to almost level (15°-20° and < 3°, respectively). Erosion is a common occurrence on the steeper slopes. Most of these dry washes start on top of the plateau and generally run eastward or southward to join the Riet, Holz and Lewer rivers. They are all typically ephemeral and are active only during and shortly after thunderstorms. Varied soil character is observed:
although most soils are very shallow, occasionally with big rocks, gravel and small stones also represent a common soil cover. At the base of the plateau some of the washes have deeper soils, which are also higher in nutrient content.
5.2 Heteropogo contorti - Ziziphodetosum mucronatae subass. nov.
This subassociation, with 13 relevés, is the typicum of the Anthephoro pubescentis - Ziziphodetum mucronatae (Figure 4h). It is characterised by the presence of the diagnostic species Heteropogon contortus, Cleome hirta, Kyphocarpa angustifolia and Senecio sp. in addition to the diagnostic species defining the Anthephoro pubescentis - Ziziphodetum mucronatae. These are also constantly accompanied by Tephrosia dregeana, Otoptera burchellii, G. flava, Tribulus terrestris, Mollugo cerviana, Kohautia caespitosa subsp. brachyloba and Cenchrus ciliaris (in addition to the constant species listed for the association). Owing to the higher moisture content in the soils, the trees grow up to 7 m tall, thus forming a short, open woodland with a tree cover of 2% (Figure 5). Species density is on average 28 species per 1000 m2, whilst a total of 109 species have been observed in this subassociation.
The Heteropogo contorti - Ziziphodetosum mucronatae occurs along the more mature rivers (Riet, Holz and Lewer). Their slopes are normally less than 3° and water remains standing in river pools for long periods. The soils in the riverbeds are normally rough gravels, which can be very stony with big rocks. The nutrient level in these gravels is low, but water content is usually relatively high. The soils of the river banks are sandy loams with a relatively high nutrient content, whilst the moisture content varies depending on the slope of the river.
6. Zygophylo pubescentis - Leucosphaeretum bainesii ass. nov.
The typicum for this association is relevé 2526, sampled at 24°32'34"S, 17°37'08"E on 10 March 2005 (Figure 4i). The association comprises 14 relevés and is characterised by the diagnostic species Leucosphaera bainesii, Commiphora glandulosa, Zygophyllum pubescens, Stipagrostis obtusa and Eriocephalus pubescens. These are constantly associated with B. foetida, S. uniplumis var. uniplumis, R. trichotomum, C. alexandri, E. cenchroides, O. burchellii, Enneapogon desvauxii, Dicoma capensis and Tribulus sp. The structure is a short, moderately closed shrubland (Figure 5), with shrubs standing approximately 0.5 m - 2 m tall and a canopy cover of about 10%, reaching up to 20%. Here, 65 species have been observed, with an average of 19 species per 1000 m2.
Zygophylo pubescentis - Leucosphaeretum bainesii is associated with shallow calcrete soils, either along the northern Haribes pan edge, as part of the flood plain system of larger rivers or on dolerite ridges. The soils are shallow (20 cm - 30 cm deep), whitish in colour and have a high clay content. The soils are expected to be alkaline with a pH of more than 7 (M. Coetzee, pers. comm., March 2010). Stone cover in the form of calcrete gravel and pebbles is often found mixed with medium to large dolerite blocks and can reach up to 80%.
Steep terraces, with slopes of between 5° and 10°, are formed along the pan edge.
7. Monsonio umbellatae - Boscietum foetidae ass. nov.
The typicum for this association is relevé 2632, sampled at 24°37'10"S, 17°37'03"E on 15 May 2005 (Figure 4j). This association was found on 17 plots in the study area. The occurrence of the low spreading forb Monsonia umbellata ('rabas') is characteristic of this association. In addition, the constant presence of S. uniplumis var. uniplumis, R. trichotomum, C. alexandri, B. foetida, S. kalahariensis, Tribulus cristatus, Ptychtolobium biflorum subsp. biflorum, G. africana, E. desvauxii, D. capensis and T. dregeana is typical for this association. The vegetation structure can be defined as a tall, open shrubland (Figure 5). The shrubs are between 0.5 m and 2 m high and represent less than 5% of the ground cover. Density is on average 17 species per 1000 m2, with a total of 67 species being observed in this association.
The Monsonio umbellatae - Boscietum foetidae typically occurs on the torras on the farm. Torras are defined as flat plains, with little or no internal relief, covered with reddish brown gravel on the surface (often more than 80%). The soils are loamy, with a crust, and with very low water infiltration capacity. This low water infiltration capacity results in patchy, near-banded vegetation, with run-off from the open patches feeding the vegetated patches (run-on). Similar vegetation patterns have been described as brousse tigree in the Sahel (Boaler & Hodge 1962; Bromley et al. 1997; Couteron et al. 2000; Thiéry, D'Herbès & Valentin 1995) and banded mulga in arid Australia (Dunkerley 1997a, 1997b).
8. Acacio senegal - Catophractetum alexandri ass. nov.
The typicum for this association is relevé 2606, sampled at 24°32'31"S, 17°28'59"E on 13 May 2005 (Figure 4l). The association comprises 80 relevés. It is characterised by Eragrostis nindensis as characteristic species, with the following constant species: S. uniplumis var. uniplumis, C. alexandri, B. foetida, L. cinereum, E. cenchroides, S. kalahariensis, Triraphis ramosissima, T. dregeana and Aristida adscensionis. The structure is a typical tall, open shrubland, with shrubs of 1 m - 2 m in height, reaching a cover of about 5%. The total cover varies between 20% and 50%. Species density is on average 19 species per 1000 m2, whilst a total of 131 species have been recorded for this association. Two subassociations have been identified within this association.
8.1 The Schmidtio kalahariensis - Boscietosum foetidae subass. nov.
The typicum of this subassociation is relevé 2546, sampled at 24°41'17"S, 17°34'54"on 14 March 2005 E (Figure 4k). This subassociation was represented by 28 relevés and is characterised by the following constant species: S. uniplumis var. uniplumis, B. foetida, L. cinereum, R. trichotomum, C. alexandri, S. kalahariensis, E. cenchroides, Tribulus sp., D. capensis, A. nebrownii, T. dregeana and Aptosimum albomarginatum. No characteristic species were identified. The structure is a tall, open shrubland (Figure 5), with a shrub cover of less than 5%. Shrub height ranges between 0.5 m and 1.8 m. The species density is on average 18 species per 1000 m2, whilst a total of 87 species have been found in this subassociation.
The Schmidtio kalahariensis - Boscietosum foetidae occurs on flat plains and footslopes as well as on ranteveld proper and forms a transition between the torras and the ranteveld. These plains are underlain by sandstones of the Nababis formation. Fragments of these form the stone cover on the soil surface, ranging from gravel to medium-sized stones and covering up to 80% of the soil surface. The low vegetative cover indicates a tendency to degrade to a similar vegetation as the Monsonio umbellatae - Boscietum foetidae on the present torras (Figure 4j).
8.2 The Acacio senegal - Catophractetosum alexandri subass. nov.
This subassociation forms the typicum of the Acacio senegal -Catophractetum alexandri (Figure 4l). A total of 52 relevés have been classified into this subassociation. It is characterised by E. nindensis as diagnostic species, with S. uniplumis var. uniplumis, C. alexandri, B. foetida, L. cinereum, E. cenchroides, T. ramosissima, G. africana, S. kalahariensis, A. adscensionis, T. dregeana and Eragrostis porosa as constant species. This subassociation forms a tall, open shrubland or occasionally a low bushveld (Figure 5), with shrubs of 0.5 m - 3 m tall. Occasional trees stand 4 m - 5 m tall. On average, 19 species have been found per 1000 m2, whilst the total species diversity is 105.
The Acacio senegal - Catophractetosum alexandri forms the ranteveld and occurs mainly in areas on top of the plateau. The plateau is dominated by very stony outcrops and small escarps (rante) of Nababis sandstone. The soils are leptosols, red in colour and very shallow, of moderately coarse to medium texture, with many coarse fragments and well drained. The surface layer is typically 10 cm - 20 cm thick. These soils have a moderate hydraulic conductivity and present a moderate infiltration rate with a low water holding capacity (ICC et al. 2000). The landscape is relatively flat on top of the plateau, but forms occasional low hills and an escarpment area to the west of the farm, with steep-sloped ravines in between.
Vegetation mapping
It was not possible to map all vegetation types, for several reasons. Firstly, many units are just too small to map with the 30-m ground resolution of Landsat 7 ETM satellite images. This holds true for the two vlei communities, E. rotifer -L. fusca and C. obtusifolia - L. fusca, as well as the Zygophylo pubescentis - Leucosphaeretum bainesii, which occurs on ridges with very limited spatial extent.
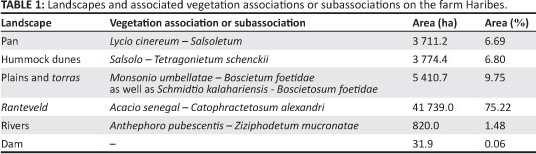
Differentiating between the Monsonio umbellatae - Boscietum foetidae and the Schmidtio kalahariensis - Boscietosum foetidae proved extremely difficult owing to their similar structures and especially similar low vegetative cover. For this reason, a landscape map was produced (Figure 6) to differentiate between plains and torras, and ranteveld. Table 1 lists the mapped landscapes, their spatial extent and their included vegetation associations or subassociations.
Discussion
Four distinct higher syntaxonomic groupings can be recognised, namely the saline pans and associated hummock dunes (as represented by the Lycio cinereum - Salsoletum and the Salsolo - Tetragonietum schenkii), the wetlands (as represented by the two vlei communities), the river systems (as represented by the Anthephoro pubescentis - Ziziphodetum mucronatae) and the surrounding plateaus (as represented by the Zygophylo pubescentis - Leucosphaeretum bainesii, the Monsonio umbellatae - Boscietum foetidae and the Acacio senegal - Catophractetum alexandri). Unfortunately the data set is too limited to define and describe such higher syntaxonomic levels at this stage. Volk and Leippert (1971) refer to the class Rhigozetea as representing the larger dwarf shrub savanna of southern Namibia; this encompasses all those syntaxa containing R. trichotomum (i.e. specifically the latter grouping).
Giess (1998) superficially described the Dwarf shrub savanna, as follows:
Characteristic for the greatest part is Rhigozum trichotomum (...). Parkinsonia africana, Acacia nebrownii, Boscia foetida subsp. foetida, B[oscia] abitrunca var. albitrunca and Catophractes alexandri as well as Karoo bushes such as Pentzia spp., Eriocephalus spp., and others are typical for this vegetation type. (Giess 1998)
This description is illustrated with three plates. Yet the Acacio senegal - Catophractetum alexandri has a too tall structure (2 m - 3 m) to be regarded as true dwarf shrub savanna. Within the broader Nama-Karoo (of which Giess' dwarf shrub savanna forms the northern extent), the Acacio senegal - Catophractetum alexandri can structurally be compared to the Acacia mellifera open shrubland of the Blouputs karroid thornveld as described for the Augrabies Falls National Park (Bezuidenhout 1996; Mucina & Rutherford 2006). However, the shrub component here is dominated by A. mellifera subsp. detinens, a species rarely found in the Acacio senegal - Catophractetum alexandri. In this respect, the Acacio senegal - Catophractetum alexandri is to be regarded as a xeric outlier of the highland savanna (sensu Giess 1998). The rocky habitat with shallow soils, and often steep slopes, is similar in both vegetation types.
The Salsolo - Tetragonietum schenckii resembles the Saline desert and dwarf shrub savanna fringe (sensu Giess 1998) described for the Etosha pan. Here the structure of the hummock dune vegetation can specifically be compared to the Okondeka duneveld (Le Roux 1980). However, the composition varies widely, with T. schenkii not occurring in northern Namibia. For the Lycio cinereum - Salsoletum no comparable vegetation type is known. It is likely that these, in a future synopsis of syntaxa, will form a class on their own.
The very close relationship between the Monsonio umbellatae - Boscietum foetidae and the two subassociations of the Acacio senegal - Catophractetum alexandri is well illustrated in the ordination diagram in Figure 3b. This leads to the assumption that these three units form degradation stages of one another:
Acacio senegal - Catophractetosum alexandri degrading to Schmidtio kalahariensis - Boscietosum foetidae, and in very extreme cases (possibly also owing to soil degradation processes such as erosion and capping) to Monsonio umbellatae With reference to biodiversity patterns, species density is - Boscietum foetidae. This hypothesis needs to be tested and comparable to other descriptions within the Nama-Karoo described in a paper in which dynamic trends are considered. biome in southern Namibia. The Nico observatories (Nico
North and Nico South) of the BIOTA Southern Africa project are the nearest to Haribes and are also situated on relatively stony soil, albeit of origin from the Dwyka formation (Geological Survey 1980). The Nico observatories display an average of 21 and 20 species per 1000 m2, respectively (Jürgens et al. 2010), compared with the 17-19 species per 1000 m2 for the Zygophylo pubescentis - Leucosphaeretum bainesii, Monsonio umbellatae - Boscietum foetidae and Acacio senegal - Catophractetum alexandri. The slightly lower values observed during the current study can be related to the survey being conducted during a single, comparatively dry year, whilst the Nico observatories had been resurveyed over a period of seven years (but unfortunately not in 2005). However, the overall number of species observed at Haribes (218) is higher than the total number of species observed at both Nico North (166) and Nico South (204). This can, in turn, be attributed to the fact that the area sampled at Haribes is far bigger and more diverse than the 1 km2 sample areas of the BIOTA observatories. Comparing the Zygophylo pubescentis - Leucosphaeretum bainesii to the Narias and Duruchaus observatories, which also have shallow soils on calcretes (thus a comparable edaphic habitat), a far lower species density was observed (19 vs 36-37 per 1000 m2). The climatic environment at the Narais and Duruchaus observatories is, however, far more moist, with a mean annual rainfall of 289 mm (Jürgens et al. 2010).
Conclusion
Haribes is unlikely ever to become a conservation area. On the contrary, the recent change in ownership also introduces an entirely different land use, with the farm being targeted for resettlement. Some 13 families have already been resettled on small parcels of the farm, each ranging between 1700 ha and 7500 ha (Cloete 2010b; Kleinhans 2010). These units, meant to support the livelihoods of these families (Werner & Odendaal 2010), have been calculated according to grazing capacity maps from the 1970s (Department Landbou
Tegniese Dienste 1979). However, the general indications are that the grazing capacity has drastically reduced throughout the entire country over the past decades (Espach et al. 2010). This is due to a multitude of factors generally associated with land degradation [e.g. bush encroachment (De Klerk 2004)]. It is thus unlikely that these small farming units will be sustainable and severe degradation of grazing resources may result (Werner & Odendaal 2010).
Within the Nama-Karoo biome in Namibia, livestock farming depends mainly on browsing of dwarf shrub species (Espach et al. 2006), with grazing playing a secondary role. This does not hold true for Haribes. A comparison of the shrub, dwarf shrub, grass and total vegetation cover of the various associations (Figure 5) shows that farming on Haribes is more dependent on grazing than browsing. The pans and surrounding dunes are mostly covered by shrubs dominated by Salsola spp. and T. schenckii, which are both known to be poisonous (Mannheimer, Marais & Schubert 2008). This makes roughly 13% of the farm unsuitable for grazing. The Zygophylo pubescentis - Leucosphaeretum bainsii on the calcrete ridges provide some dwarf shrub browse, but is very limited in extent. The Monsonio umbellatae - Boscietum foetidae (torras) and the Acacio senegal - Catophractetum alexandri (ranteveld) provide the main grazing resources on the farm. However, as pointed out in the discussion, these seem to form a degradation gradient from each other; thus, extensive overgrazing will lead to desert-like torra plains.
Global climate change has been identified as a reality in recent years. Temperature increases coupled with less precipitation, in more erratic events, are predicted for southern Africa (Parry et al. 2007). Based on these forecasts, it is predicted that large parts of southern Namibia will become unsuitable for livestock farming, as it will turn into a virtual desert (Midgley et al. 2005). Similar scenarios have been developed for the Kalahari basin, with forecasts of remobilisation of sand dunes as far north as southern Angola (Thomas, Knight & Wiggs 2005). In addition, a human-induced drought, as described for other Nama-Karoo regions (Booysen & Rowswell 1983), threatens. This is aggravated by the fact that farming on Haribes depends largely on grazing, meaning that protection of the perennial grass cover is essential for sustainable use of the farm in the long term. Perennial grasses have been shown to be able to withstand prolonged droughts (i.e. periods of below-normal precipitation), but are unable to withstand heavy grazing together with arid conditions (O'Connor 1991, 1994). It is thus likely that overgrazing in the ranteveld may lead to a composition (and vegetation cover) similar to that of the present torras. When planning the redistribution of the farm Haribes, it should be kept in mind that only 75% of the farm (the ranteveld) provides adequate grazing and that even this grazing needs to be utilised judiciously to prevent severe degradation to desert-like conditions.
Acknowledgements
We are grateful to Mr and Mrs Poggenpoel, previous managers at the farm Haribes, for making Volk's data of 1956 available to us and for their kind support during fieldwork. The assistance of staff members at the National Herbarium of Namibia (WIND) during specimen identification is gratefully acknowledged. Many thanks also go to Mrs Marina Coetzee of the School of Natural Resources and Tourism, Polytechnic of Namibia for providing information on soil properties. This work was financially supported by the German Federal Ministry of Education and Research under their BIOLOG programme, promotion number 01 LC 0024A, as part of the BIOTA Southern Africa project.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this paper.
Authors' contributions
B.J.S. (National Botanical Research Institute) initiated the project. W.J.J. (School of Natural Resources and Tourism) was responsible for the vegetation surveys and the habitat and structural descriptions of the associations, whilst B.J.S. (National Botanical Research Institute) was responsible for data analysis, diagnostic descriptions of the associations, and mapping.
References
Bezuidenhout, H., 1996, 'The major vegetation communities of the Augrabies Falls National Park, Northern Cape. 1. The southern section', Koedoe 39, 7-24. [ Links ]
Boaler, S.B. & Hodge, C.A.H., 1962, 'Vegetation stripes in Somaliland', Journal of Ecology 50, 465-474. http://dx.doi.org/10.2307/2257457 [ Links ]
Booysen, J. & Rowswell, D.I., 1983, 'The drought problem in the Karoo areas', Proceedings of the Grassland Society of southern Africa 18, 40-45. [ Links ]
Botha, L., 1996, Rainfall as an indicator of the agroclimate of Namibia, M.Dip.Tech. thesis, Faculty of Engineering, Technikon Pretoria. [ Links ]
Bromley, J., Brouwer, J., Barker, A.P., Gaze, S.R. & Valentin, C., 1997, 'The role of surface water redistribution in an area of patterned vegetation in a semi-arid environment, south-west Niger', Journal of Hydrology 198, 1-29. http://dx.doi.org/10.1016/S0022-1694(96)03322-7 [ Links ]
Bullard, J.E. & Nash, D.J., 2000, 'Valley-marginal sand dunes in the south-west Kalahari: their nature, classification and possible origins', Journal of Arid Environments 45, 369-383. http://dx.doi.org/10.1006/jare.2000.0646 [ Links ]
Burke, A. & Strohbach, B.J., 2000, 'Vegetation studies in Namibia', Dinteria 26, 1-24. [ Links ]
Chytrý, M., Tichý, L., Holt, J. & Botta- Dukat, Z., 2002, 'Determination of diagnostic species with statistic fidelity measures', Journal of Vegetation Science 13, 79-90.http://dx.doi.org/10.1111/j.1654-1103.2002.tb02025.x [ Links ]
Cloete, L., 2010a, 'Government buys resettlement farm for N$10m', The Namibian, 06 August, p. 5. [ Links ]
Cloete, L., 2010b, 'Landless families become proud owners of land', The Namibian, 22 November, p. 7. [ Links ]
Couteron, P., Mahamane, A., Ouedraogo, P. & Seghieri, J., 2000, 'Differences between banded thickets (tiger bush) at two sites in West Africa', Journal of Vegetation Science 11, 321-328. http://dx.doi.org/10.2307/3236624 [ Links ]
Craven, P. (ed.), 1999, A checklist of Namibian plant species, Southern African Botanical Diversity Network, Windhoek (Southern African Botanical Diversity Network Report, no 7). [ Links ]
De Klerk, J.N., 2004, 'Bush encroachment in Namibia', report on Phase 1 of the Bush Encroachment Research, Monitoring and Management Project, Ministry of Environment and Tourism, Windhoek. [ Links ]
De Pauw, E., Coetzee, M.E., Calitz, A.J., Beukes, H. & Vits, C., 1998, 'Production of an agro-ecological zones map of Namibia (first approximation). Part II: Results', Agricola 10, 33-43. [ Links ]
Definiens version 5, 2006, computer software, Definiens AG, Munich. [ Links ]
Departement Landbou Tegniese Dienste, 1979, Die afbakening van redelike homogene boerderygebiede van die noordelike en sentrale substreke van S.W.A. met die heersende knelpunte en beoogde ontwikkleingsprogramme vir die verskillende bedryfstakke [The delimitation of relative homogenous farming areas in the northern and central subregions of SWA with the present constraints and intended development programmes for the different enterprises], Departement Landbou Tegniese Dienste, Windhoek. [ Links ]
Dunkerley, D.L., 1997a, 'Banded vegetation: survival under drought and grazing pressure based on a simple cellular automaton model', Journal of Arid Environments 35, 419-428. http://dx.doi.org/10.1006/jare.1995.0158 [ Links ]
Dunkerley, D.L., 1997b, 'Banded vegetation: development under uniform rainfall from a simple cellular automaton model', Plant Ecology 129, 103-111. http://dx.doi.org/10.1023/A:1009725732740 [ Links ]
Eastman, J.R., 2006, IDRISI. The Andes Edition, computer software, Clark University, Worcester, Massachusetts. [ Links ]
Edwards, D., 1983, 'A broad-scale structural classification of vegetation for practical purposes', Bothalia 14, 705-712. [ Links ]
Espach, C., 2006, 'Rangeland productivity modelling: Developing and customising methodologies for land cover mapping in Namibia', Agricola 16, 20-27. [ Links ]
Espach, C., Lubbe, L.G. & Ganzin, N., 2006, 'Determining grazing capacity in Namibia: Approaches and methodologies', Agricola 16, 28-39. [ Links ]
Espach, C., Lubbe, L.G. & Ganzin, N., 2010, 'Determining grazing capacity in Namibia with the aid of remote sensing', Agricola 20, 41-61. [ Links ]
FAO. See Food and Agriculture Organization.
Food and Agriculture Organization, 1995, Global and national soils and terrain digital databases (SOTER), Food and Agriculture Organization, Water Development Division, Rome (World Soil Resources Reports, no. 74 Rev. 1). [ Links ]
Geological Survey, 1980, South West Africa/Namibia geological map 1:1000000. [ Links ]
Giess, W., 1971, 'A preliminary vegetation map of South West Africa', Dinteria 4, 1-114. [ Links ]
Giess, W., 1998, 'A preliminary vegetation map of Namibia', Dinteria 4, 1-112. [ Links ]
Hennekens, S.M. & Schaminée, J.H.J., 2001, 'TURBOVEG, a comprehensive data base management system for vegetation data', Journal of Vegetation Science 12, 589591. http://dx.doi.org/10.2307/3237010 [ Links ]
Hill, M.O. & Gauch, H.G., 1980, 'Detrended correspondence analysis: An improved ordination technique', Vegetatio 42, 47-58. http://dx.doi.org/10.1007/BF00048870 [ Links ]
ICC, MAWRD & AECI, 2000, 'Project to support the Agro-Ecological Zoning Programme (AEZ) in Namibia', report, Institut Cartografie de Catalunya (ICC), Namibian Ministry of Agriculture, Water and Rural Development (MAWRD) and Spanish Agency for International Co-operation (AECI), Windhoek. [ Links ]
Irish, J., 1994, 'The biomes of Namibia, as determined by objective categorisation', Navorsinge van die Nasionale Museum Bloemfontein 10, 549-592. [ Links ]
Jarvis, A., Reuter, H.I., Nelson, A. & Guevara, E., 2008, Hole-filled seamless SRTM data v4, viewed 03 September 2010 from http://srtm.csi.cgiar.org/. [ Links ]
Jürgens, N., Haarmeyer, D.H., Luther-Mosebach, J., Dengler, J., Finckh, M. & Schmiedel, U. (eds.), 2010, Patterns at local scale: The BIOTA Observatories, Biodiversity in southern Africa, Klaus Hess, Göttingen & Windhoek. [ Links ]
Kleinhans, T., 2010, 'Staat gee Suide-grond - met voorwaardes [Government gives land in South - with conditions]', Republikein, 25 November, p. 5. [ Links ]
Lancaster, N., 1986, 'Pans in the southwestern Kalahari: A preliminary report', Palaeoecology of Africa and the surrounding islands 17, 59-67. [ Links ]
Le Roux, C.J.G., 1980, 'Vegetation classification and related studies in the Etosha National Park', DSc thesis, Dept. of Plant Production, University of Pretoria. [ Links ]
Lubbe, L.G., 2005, 'Towards an updated carrying capacity map for Namibia: A review of the methodologies currently used to determine carrying capacity in Namibia', Agricola 15, 33-39. [ Links ]
Mannheimer, C., Marais, A. & Schubert, S., 2008, Toxic plants of veterinary importance in Namibia, 2nd ed., Ministry of Agriculture, Water and Forestry, Windhoek. (Addendum to Agricola 2007) [ Links ]
McCune, B., Grace, J.B. & Urban, D.L., 2002, Analysis of ecological communities, MjM Software, Gleneden Beach, Oregon. [ Links ]
Mendelsohn, J., Jarvis, A., Roberts, C. & Robertson, T., 2002, Atlas of Namibia, David Phillips, Cape Town, pp. 55-100. [ Links ]
Midgley, G., Hughes, G., Thuiller, W., Drew, G. & Foden, W., 2005, 'Assessment of potential climate change impacts on Namibia's floristic diversity, ecosystem structure and function', report, Climate Change Research Group, South African National Biodiversity Institute for the Namibian National Biodiversity Programme, Directorate of Environmental Affairs, Cape Town. [ Links ]
Ministry of Agriculture, Water and Rural Development, 2001, Namibian Agricultural Resources Information System (NARIS), Ministry of Agriculture, Water and Rural Development, Windhoek. [ Links ]
Mucina, L. & Rutherford, M.C. (eds.), 2006, The vegetation of South Africa, Lesotho and Swaziland, South African National Biodiversity Institute, Pretoria (Strelitzia, no. 19). [ Links ]
Mueller-Dombois, D. & Ellenberg, H., 1974, Aims and methods of vegetation ecology, John Wiley & Sons, New York. [ Links ]
O'Connor, T.G., 1991, 'Influence of rainfall and grazing on the compositional change of the herbaceous layer of a Sandveld savanna', Journal of the Grassland Society of southern Africa 8, 103-109. [ Links ]
O'Connor, T.G., 1994, 'Composition and population responses of an African savanna grassland to rainfall and grazing', Journal of Applied Ecology 31, 155-171. http://dx.doi.org/10.2307/2404608 [ Links ]
Oldeman, L.R. & Van Engelen, V.W.P., 1993, 'A world soils and terrain digital database (SOTER): An improved assessment of land resources', Geoderma 60, 309-325. http://dx.doi.org/10.1016/0016-7061(93)90033-H [ Links ]
Parry, M.L., Canziani, O.F., Palutikof, J.P., Van der Linden, P.J. & Hanson, C.E. (eds.), 2007, Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, Cambridge University Press, Cambridge. [ Links ]
Pretorius, M., 2010, '50 100 hektaar in Suide vir hervestiging gekoop [50 100 hectares bought in South for resettlement]', Republikein, 10 Augustus, p. 7. [ Links ]
Roleček, J., Tichý, L., Zelený, D. & Chytrý, M., 2009, 'Modified TWINSPAN classification in which the hierarchy respects cluster heterogeneity', Journal of Vegetation Science 20, 596-602. http://dx.doi.org/10.1111/j.1654-1103.2009.01062.x [ Links ]
Shaw, P.A., 1988, 'Lakes and pans', in B.P. Moon & G.F. Dardis (eds.), The geomorphology of southern Africa, pp. 120-144, Southern Book Publishers, Johannesburg. [ Links ]
Shaw, P.A. & Thomas, D.S.G., 1997, 'Pans, playas and salt lakes', in D.S.G. Thomas (ed.), Arid zone geomorphology, pp. 293-317, John Wiley & Sons, Chichester. [ Links ]
Strohbach, B.J., 2001, 'Vegetation survey of Namibia', Journal of the Namibia Scientific Society 49, 93-124. [ Links ]
Strohbach, B.J. & Jürgens, N., 2010, 'Towards a user-friendly vegetation map of Namibia: Ground truthing approach to vegetation mapping', in U. Schmiedel & N. Jürgens (eds.), Biodiversity in southern Africa: Patterns and processes at regional scale, pp. 46-56, Klaus Hess Publishers, Göttingen & Windhoek. [ Links ]
Thiéry, J.M., D'Herbès, J.M. & Valentin, C., 1995, 'A model simulating the genesis of banded vegetation patterns in Niger', Journal of Ecology 83, 497-507. http://dx.doi.org/10.2307/2261602 [ Links ]
Thomas, D.S.G., Knight, M. & Wiggs, G.F.S., 2005, 'Remobilization of southern African desert dune systems by twenty-first century global warming', Nature 435, 12181221. http://dx.doi.org/10.1038/nature03717, PMid:15988522 [ Links ]
Tichý, L., 2002, 'JUICE, software for vegetation classification', Journal of Vegetation Science 13, 451-453. http://dx.doi.org/10.1111/j.1654-1103.2002.tb02069.x [ Links ]
Volk, O.H. & Leippert, H., 1971, Vegetationsverháltnisse im Windhoeker Bergland, Südwestafrika [Vegetation relationships in the Windhoek Mountainveld, Southwest Africa], Journal der S.W.A. Wissenschaftliche Gesellschaft XXV, 5-14. [ Links ]
Weber, H.E., Moravec, J. & Theurillat, J.P., 2000, 'International Code of Phytosociological Nomenclature, 3rd edn.', Journal of Vegetation Science 11, 739-768. http://dx.doi.org/10.2307/3236580 [ Links ]
Werner, W. & Odendaal, W., 2010, Livelihoods after land reform. Namibia country study, Land, Environment and Development Project, Legal Assistance Centre, Windhoek. [ Links ]
 Correspondence to:
Correspondence to:
Ben Strohbach
Email:bens@nbri.org.na
Private Bag 13184, Windhoek,
Namibia
Received: 14 Dec. 2010
Accepted: 16 Sept. 2011
Published: 29 Mar. 2011
Nomenclature follows Craven (1999) and subsequent name changes used by the National Herbarium of Namibia (WIND).
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}