










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkKoedoe
On-line version ISSN 2071-0771
Print version ISSN 0075-6458
Koedoe vol.52 n.1 Pretoria Jan. 2010
ORIGINAL RESEARCH
Plant diversity and flowering displays on old fields in the arid Namaqua National Park, South Africa
Margaretha (Gretel) W. van Rooyen; Ronel Henstock; Noel van Rooyen; Helga van der Merwe
Department of Plant Science, University of Pretoria, South Africa
ABSTRACT
The main aim of this study was to classify the vegetation on the abandoned fields in the Namaqua National Park and to investigate whether the time since abandonment has impacted on the diversity and mass flowering displays. Sixty-two old field sample sites were surveyed using the Braun-Blanquet method. Floristic data were analysed to produce a hierarchical classification, while a principal coordinates analysis was used to establish whether the ordination supported the classification. Species richness (total number of taxa), as well as the Shannon-Wiener diversity index, was calculated per vegetation unit for the different life forms. Four old field communities were identified, each containing a unique complement of species, which differed in their potential to produce a mass flowering display. On the Skilpad section of the park, Ursinia cakilefolia dominated the spectacular mass floral displays for which the park has become well known. Old fields in other parts of the park did not produce the same spectacular displays. Although total and perennial species richness did not differ significantly with time since abandonment, the richness of annual species, in particular of the showy annual species, decreased with time since abandonment. The relative contribution of the perennial species to vegetation cover increased with time since abandonment, whereas the relative contribution made by showy annual species to vegetation cover declined with time since abandonment. Although species composition and the potential to produce mass flowering displays of the four communities differed, diversity parameters were similar. However, to maintain a flower display for tourists on the Skilpad section, a degree of disturbance is essential.
Conservation implication: Although deliberate disturbance is not desirable in a national park, we find that some disturbance is essential to maintain a spectacular flowering display. However, these intentionally disturbed old fields cover only a small proportion of the total area of the park. All other old fields in the park should not be disturbed, and should be allowed to recover naturally.
Keywords: Abandoned croplands; annual species; disturbance; fallow lands; succession; vegetation recovery
INTRODUCTION
The Namaqua National Park is situated within the Succulent Karoo Biome - a biome recognised by the International Union for Conservation of Nature (IUCN) as one of the global hotspots of diversity (Critical Ecosystem Partnership Fund 2003; Mittermeier et al. 2000; Myers et al. 2000), and one of only two hotspots that are entirely arid (Conservation International, undated). The biome is unique because almost half of its plant species are endemic, while the high diversity of leaf succulents and bulbous species is unrivalled amongst similar deserts (Cowling et al. 1999; Mucina et al. 2006). The park forms an integral part of the conservation network in Namaqualand to conserve the rich biodiversity of the region.
Namaqualand has become synonymous with mass displays of wild flowers (Van Rooyen 1999). These mass displays of annuals are often a sign of land degradation and are especially prominent on croplands and other disturbed, trampled or overgrazed sites (Van Rooyen 2002). Flowering displays on cropland are often at their best in the fields lying fallow between cultivation (Van Rooyen et al. 1996). Whereas natural rangeland in Namaqualand contains a diverse array of showy annual species, the mass effect on disturbed sites is created by the dominance of a few pioneer species and, accordingly, mass displays are believed to be associated with a loss of plant diversity (Van Rooyen 2002).
The Namaqua National Park developed as an extension to the Skilpad Wildflower Reserve. The 930-ha farm, Skilpad, was originally selected by WWF-SA (at that stage the South African Nature Foundation) as a wildflower reserve because the farm was renowned for its springtime display on the fallow lands (Van Rooyen et al. 1996). This floral spectacle draws thousands of tourists annually and is a valuable source of income to the region, and especially to the park (James et al. 2007). The expansion of the park, from its small 930 ha beginning to its current size of 140 035 ha (South African National Parks 2010), has led to the inclusion of many more vegetation types present in the region, and it now includes old fields situated in many of these different vegetation types.
The objectives of the current study were, (1) to classify the old field vegetation, (2) to compare the communities in terms of their potential for producing mass flowering displays to draw tourists, (3) to compare diversity and life form composition across different communities and (4) to investigate the effect of time since abandonment on the diversity and life form composition of the old fields in the Skilpad section of the park.
METHODS
Study area
Geologically, the Namaqua National Park is located on granites and gneisses of the Bushmanland Terrane of the Namaqua-Natal Metamorphic Province (Cornell et al. 2006). The soils in the region are generally shallow and show minimal development, yet the diversity and spatial variability of the soils's physical and chemical properties are exceptionally high, and could be fundamental in governing biodiversity and ecosystem function (Francis et al. 2007; Petersen 2008). Although a diverse array of land types occurs in the park, croplands are found primarily on the Ag, Fc and Ib land types (AGIS 2007). Land Type Ag is characterised by red-yellow apedal, freely drained soils with a high base status, and is less than 300 mm deep; Land Type Ib comprises miscellaneous land classes and rocky areas with miscellaneous soils; and Land Type Fb comprises Glenrosa and/or Mispah soil forms, with lime rare or absent in upland soils, but generally present in the low-lying soils (Du Plessis 1987).
Croplands in the park are found primarily towards the east of the park on the high-lying escarpment areas (Figure 1). In the mountainous Namaqualand landscape suitable sites for crop cultivation are limited and farmers have selected sites with a level terrain or gentle slopes, little rock cover and deep soils. This particular habitat has therefore been largely transformed by agriculture throughout the region.
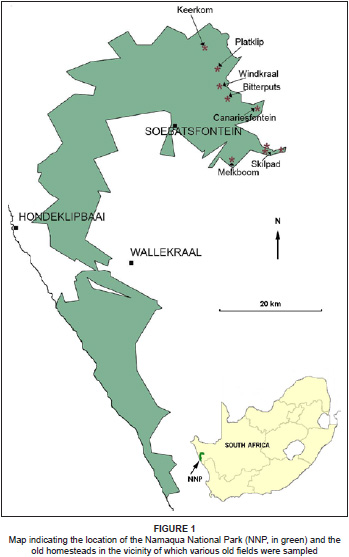
The climate of the region is characterised by relatively reliable, albeit low (50 mm - 250 mm per annum), winter rainfall (> 60% winter precipitation) arriving between May and September (Desmet 2007). The mean annual rainfall measured over 15 years in the Skilpad section is 340 mm, whereas the mean annual rainfall at Soebatsfontein located on the coastal plain below the escarpment is 140 mm per annum (South African National Parks 2010). Croplands are thus predominantly found on the escarpment in the eastern sections of the park (Figure 1), where the rainfall is higher. The presence of the cold Atlantic Ocean in the west moderates temperatures throughout Namaqualand and provides alternative sources of moisture in the form of coastal fog and heavy dew, experienced in the winter months (Desmet 2007).
Field surveys and statistical analysis
The Braun-Blanquet method of vegetation sampling (Werger 1974) was used to conduct a floristic analysis of the old fields of the Namaqua National Park. Forty-three old fields were sampled in 2006 and an additional 19 were surveyed in 2007. Plot sizes of 20 m x 10 m were used to quantify the vegetation on the old fields. Global Positioning System (GPS) coordinates were taken at each site and various environmental data such as aspect, slope, drainage, erosion, soil texture and rock cover were recorded.
A cover-abundance value, according to the Braun-Blanquet scale (Van der Maarel 2007; Werger 1974), was noted for each species present in a plot. An analysis of the floristic data was conducted using the TURBOVEG and MEGATAB computer packages (Hennekens & Schamineé 2001): TURBOVEG software was used to capture the vegetation data and a TWINSPAN (Two Way Species Indicator Analysis; Hill 1979) was run in MEGATAB as a first step in the classification of the vegetation data. The resulting table of sample plots against species was further refined using Braun-Blanquet tabulation procedures (Werger 1974) to produce a hierarchical classification.
To visualise the relationship between the communities the floristic data were ordinated using principal coordinates analysis (PCoA) (McCune & Grace 2002) in the SYN-TAX computer program (Podani 2001). Cover-abundance values were converted to percentages and the percentage values standardised using a natural logarithmic (loge) standardisation. The Bray-Curtis distance measure was applied for the ordination.
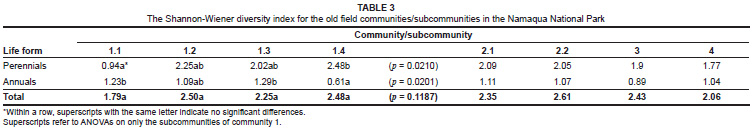
Species richness (total number of taxa), as well as the Shannon-Wiener index of diversity per community or subcommunity, was calculated as the mean for all sample sites for that vegetation unit. Both species richness values and the Shannon-Wiener diversity index were also calculated separately for the perennial, annual and geophyte life forms. The annual species richness was further subdivided into those species that have the potential for producing a flower display (referred to as showy species) and those species that do not contribute to a flower display (non-showy species). Diversity data were analysed using a one-way analysis of variance (ANOVA) in the STATISTICA computer program (Statistica version 8, Statsoft, Tulsa). The Tukey post hoc test was used to determine significant differences (p < 0.05) between communities or subcommunities. Diversity data were first compared across all vegetation units and thereafter the subcommunities within community 1 were compared to one another.
Nomenclature follows that of 'Plants of southern Africa (version 2.5: 'an online checklist' http://posa.sanbi.org). Non-flowering geophytes that could not be identified were excluded from the analysis. These geophytes were infrequently encountered and did not contribute to the delineation of communities.
RESULTS
Classification of plant communities
Four communities were identified and two of the communities were further divided into subcommunities. The communities and subcommunities are as follows (Table 1 [see online Appendix] and Figure 1):
1. Ursinia cakilefolia old field vegetation
1.1 Leysera gnaphalodes-Ursinia cakilefolia old field vegetation
1.2 Heliophila variabilis-Ursinia cakilefolia old field vegetation
1.3 Felicia bergeriana-Ursinia cakilefolia old field vegetation
1.4 Stipagrostis zeyheri-Ursinia cakilefolia old field vegetation
2. Felicia australis old field vegetation
2.1 Grielum humifusum-Felicia australis old field vegetation
2.2 Galenia africana-Felicia australis old field vegetation
3. Salsola kali old field vegetation
4. Oncosiphon grandiflorum old field vegetation
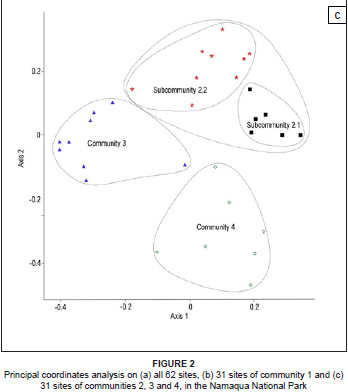
Overall, the classification of the floristic data provided in Table 1 was supported by the ordination (Figure 2a, 2b and 2c). The PCoA of the floristic data of all 62 relevés clearly indicated the separation between community 1 and communities 2 to 4 (Figure 2a). Two separate ordinations were then run, the first on the floristic data of community 1 and a second on the floristic data of communities 2, 3 and 4. In the ordination of community 1 the subcommunities were arranged along the first axis from right (subcommunity 1.1) to left (subcommunity 1.4) and this gradient was related to the time since abandonment (Figure 2b). The ordination of communities 2, 3 and 4 produced three separate clusters (Figure 2c).

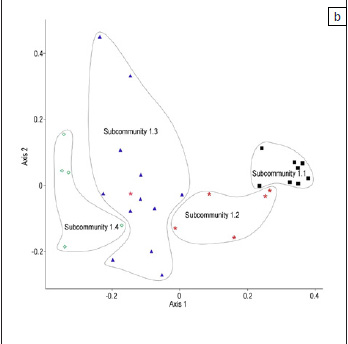
The current study focused on a vegetation 'state' in disturbed habitats, predominantly composed of annual plant species, and thus the vegetation descriptions were based strongly on annual species. Because of the dynamic nature of this disturbed vegetation, some of the subcommunities are arranged along a continuum (Figure 2b) and some of the vegetation units identified here may even be transient.
1. Ursinia cakilefolia old field vegetation
This plant community was located on the farm Skilpad in the Namaqualand Klipkoppe vegetation type (SKn1; Mucina et al. 2006) at altitudes from 620 m to 755 m above sea level. Most of this community was found on Land Type Ag94, although Land Type Ib235 also occurred. Community 1 was generally found on gentle or moderate slopes, on well-drained sandy loams or loamy sands with no rock cover.
The vegetation of this community was characterised by species group G with, generally, a high cover of Ursinia cakilefolia. Other common species included Gazania leiopoda and Felicia bergeriana (species group G), Heliophila variabilis, Leysera gnaphalodes, Conicosia elongata and Oxalis obtusa (species group AC). This community was divided into four subcommunities.
1.1 Leysera gnaphalodes-Ursinia cakilefolia old field vegetation: Subcommunity 1.1 was found along the circular tourist route on the Skilpad section of the Namaqua National Park and represented those fields in the park that are regularly disturbed (tilled), at approximately four-year intervals, to produce mass flowering displays. The subcommunity occurred exclusively on Land Type Ag94 and the gentle slopes and plains, with no rock cover, comprised well-drained loamy sand and sandy loam soils. Altitudes ranged from 686 m to 749 m above sea level.
Senecio arenarius and Helichrysum cf. hebelepis (species group A) differentiated this subcommunity. Lotononis leptoloba (species group D), Ursinia cakilefolia (species group G), Heliophila variabilis and Leysera gnaphalodes (species group AC) were present with high cover values. Pentaschistis tomentella (species group J), Arctotheca calendula (species group V), Lapeirousia silenoides (species group Y), Lessertia diffusa, Conicosia elongata, Wahlenbergia annularis and Dischisma spicatum (species group AC) were always present in subcommunity 1.1.
1.2 Heliophila variabilis-Ursinia cakilefolia old field vegetation: Subcommunity 1.2 was found towards the eastern part of the Skilpad section. This subcommunity occurred on Land Type Ag94, on old fields that had been actively cultivated until the Skilpad Wildflower Reserve was established, but have not been disturbed since then. In general, the old fields in subcommunity 1.2 were 12-15 years old. The loamy sands were well- drained and the fields occurred on level to gentle slopes at altitudes ranging from 620 m to 680 m above sea level. No rock cover was present, with the exception of one site with a rock cover of 5%.
The species of species group B, such as Gymnodiscus linearifolia and Lotononis pentaphylla, characterised this subcommunity. Ursinia cakilefolia, Gazania leiopoda (species group G), Pentaschistis tomentella (species group J), Heliophila variabilis and Leysera gnaphalodes (species group AC) often had high cover values. Species with a high constancy were Manulea altissima, Othonna pavonia (species group C), Arctotheca calendula (species group V), Limeum africanum, Lapeirousia silenoides (species group Y), Dimorphotheca sinuata and Conicosia elongata (species group AC).
1.3 Felicia bergeriana-Ursinia cakilefolia old field vegetation: Subcommunity 1.3 included old fields in the park (eastern part of the Skilpad section) as well as in the adjacent farmland at altitudes ranging from 624 m to 680 m above sea level. Field age of the sites in the park ranged from approximately 20-60 years. In contrast, the fields on the adjacent farmland were estimated to be approximately 10-20 years old at the time of the study. The gentle to moderately sloping old fields, on Land Type Ag94, were characterised by well-drained loamy sands with a low (< 1%) or zero rock cover. With the exception of the two sites abandoned approximately 60 years ago, all fields were still clearly distinguishable from the surrounding vegetation.
This subcommunity had no diagnostic species group and was transitional between subcommunities 1.2 and 1.4. Abundant species included Ursinia cakilefolia, Gazania leiopoda, Felicia bergeriana (species group G), Trichogyne glomerata (species group I), Pentaschistis tomentella, Cotula barbata (species group J), and Heliophila variabilis (species group AC). Species with a high constancy were Nylandtia spinosa (species group G), Trachyandra falcata, Lapeirousia silenoides (species group Y), Leysera gnaphalodes, Conicosia elongata, Oxalis obtusa and Ehrharta calycina (species group AC).
1.4 Stipagrostis zeyheri-Ursinia cakilefolia old field vegetation: Subcommunity 1.4 represented those fields that had been cultivated more than 50-80 years ago. Most of the sites were surveyed on the adjacent farmland. Land Type Ib235 dominated this subcommunity, which was found at altitudes ranging from 660 m to 755 m above sea level. The well-drained loamy sands occurred on level to gentle slopes and usually surface rocks were absent. The vegetation at these sites could hardly be distinguished from the natural vegetation and most sites were pointed out by the farmer.
Species group E, with species such as Searsia horrida, Ballota africana and Aspalathus cf. pulicifolia, characterised this subcommunity. Dorotheanthus bellidiformis (species group F), Dodonaea viscosa and Nylandtia spinosa (species group G), Microloma sagittatum (species group I), as well as Heliophila variabilis and Oxalis obtusa (species group AC) were generally present. Large perennial shrub species such as Searsia horrida, Euclea tomentosa (species group E), Wiborgia mucronata (species group F), Nylandtia spinosa and Dodonaea viscosa (species group G) and a high cover of Stipagrostis zeyheri (species group AB) distinguished this subcommunity from the others.
2. Felicia australis old field vegetation
This community was located in the vicinity of the farms Keerkom and Canariesfontein and the old homestead of Bitterputs. It occurred predominantly on Land Type Fb154 and in a few cases on Land Type Ib235. The sandy loam soils were usually moderately to well- drained but on occasion waterlogged or poorly drained. Rock cover varied between zero and < 10% on the gentle to moderate slopes of these old fields. Altitudes ranged from 473 m to 705 m above sea level. The natural vegetation was located within the Namaqualand Klipkoppe (SKn1) vegetation type (Mucina et al. 2006).
Community 2 was characterised by the presence and high cover of Felicia australis (species group Q) together with Galenia africana (species group AC). Other conspicuous species were Hermannia amoena (species group U), Calobota sericea (species group V), Zaluzianskya benthamiana (species group Y), Heliophila variabilis, Helichrysum tinctum, Wahlenbergia annularis and Dischisma spicatum (species group AC). Communities 2 and 3 showed strong floristic affinities as indicated by the presence of species groups T and U.
2.1 Grielum humifusum-Felicia australis old field vegetation: Subcommunity 2.1 was found on the farm Canariesfontein at altitudes ranging from 483 m to 705 m above sea level. The slopes were gentle to moderate and usually covered with moderately to well-drained, sandy loam soils, and no to little (2% - < 10%) rock cover.
This subcommunity did not have a strong species group differentiating it from other communities. Commonly occurring species shared with community 1 were Trichogyne glomerata (species group I) and Pentaschistis tomentella (species group J). Dominant species occurring in subcommunity 2.1 included Felicia australis (species group Q), Hermannia amoena (species group U), Calobota sericea (species group V), Gorteria diffusa and Hermannia trifurca (species group AB), Galenia africana, Leysera gnaphalodes, Dimorphotheca sinuata, Grielum humifusum and Helichrysum tinctum (species group AC).
2.2 Galenia africana-Felicia australis old field vegetation: Subcommunity 2.2 was located on the farm Keerkom and in the vicinity of the old Bitterputs and Rondeklip homesteads. It was found on both Land Type Fb154 and Ib235 and the sandy loam soils ranged from waterlogged to well-drained. Surface rocks were generally absent, but at two sites a low rock cover (3% - 5%) was found. The altitudes ranged from 473 m to 686 m above sea level.
Felicia australis (species group Q), Drosanthemum hispidum (species group T), and Galenia africana (species group AC) dominated subcommunity 2.2. Hermannia amoena (species group U), Senecio cardaminifolius (species group V), Zaluzianskya benthamiana (species group Y), Heliophila variabilis and Wahlenbergia annularis (species group AC) had a high constancy in this subcommunity. Two variants could be distinguished within this subcommunity: the first occurring on Keerkom and characterised by species group L, and the second mostly at Bitterputs and Rondeklip and characterised by species group O.
3. Salsola kali old field vegetation
Community 3 occurred in the vicinity of Platklip and Windkraal at altitudes ranging from 477 m to 622 m above sea level. The old fields of this community were predominantly found within Land Type Ib124 on level areas with no rock cover. The soils were usually well-drained loamy sands. The natural vegetation fell within the Namaqualand Klipkoppe (SKn1) vegetation type (Mucina et al. 2006).
Salsola kali and Helichrysum oxybelium (species group S) differentiated this community. Furthermore, the strong presence of Drosanthemum hispidum (species group T), Atriplex lindleyi, Galenia fruticosa (species group W), Zaluzianskya benthamiana, Aizoon canariense (species group Y), Calobota halenbergensis (species group AA) and Galenia africana (species group AC) also distinguished this group. Community 3 was closely related to community 2 (species groups T and U) as well as community 4 (species group AA), with species groups W and X indicating the affinities between all three communities.
4. Oncosiphon grandiflorum old field vegetation
Located on Melkboom, this community was the only community that did not occur in the Namaqualand Klipkoppe (SKn1) vegetation type of Mucina et al. (2006), but was located in the Namaqualand Heuweltjieveld (SKn4). With the exception of one plot, these old fields were found on Land Type Ag96, on gentle to moderate slopes, with a rock cover of less than 2%. A low altitude (264 m - 335 m above sea level) and well-drained sandy loams distinguished this community.
Species group Z, with species such as Oncosiphon grandiflorum and Arctotis fastuosa, differentiated this community. Lotononis parviflora, Karroochloa schismoides (species group AA) and Heliophila variabilis, Leysera gnaphalodes, Galenia africana, Dimorphotheca sinuata, Grielum humifusum and Helichrysum tinctum (species group AC) were common species that occurred across most, if not all, the relevés representing community 4.
Diversity
A comparison between the four communities revealed no significant differences in the species richness or Shannon-Wiener diversity index for all the species (p > 0.05), perennial species (p > 0.05) or annual species (p > 0.05).
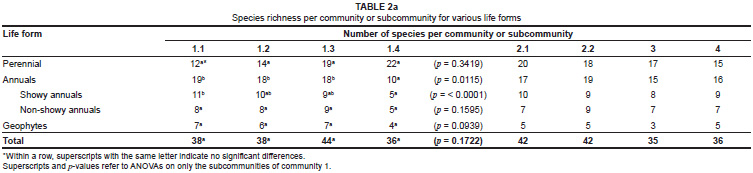
When only community 1 was considered there were, however, significant differences in the diversity parameters between the subcommunities. Annual species richness was significantly lower in subcommunity 1.4 than in the other subcommunities (Table 2a). These differences were primarily due to differences in the showy component of the annual species, with no differences observed between species richness of the non-showy annual species (Table 2a). The Shannon-Wiener diversity index confirmed the lower annual diversity in subcommunity 1.4 than in subcommunity 1.1 (see Table 3 superscripts for significance). The Shannon-Wiener index also indicated the opposite trend for the perennial species diversity, with subcommunity 1.4 having a significantly higher perennial diversity than subcommunity 1.1 (Table 3).
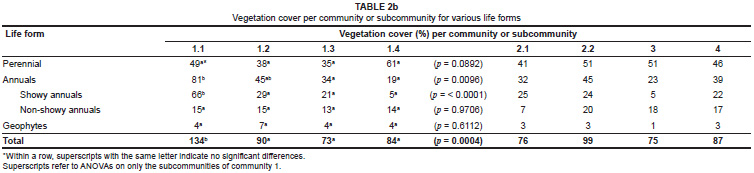
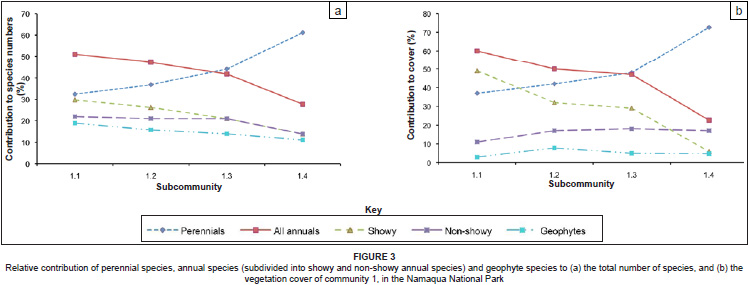
Most of the differences in species richness among the subcommunities of community 1 were reflected in the contributions made to the vegetation cover by the different life forms (Table 2b). In Figure 3a and Figure 3b the relative contributions of the species per life form to the total number of species (Figure 3a) and the relative contribution of vegetation cover per life form to the total vegetation cover (Figure 3b) are illustrated. These figures clearly show the increased contribution of perennial species to species richness and cover from subcommunity 1.1 to 1.4, and a concomitant decrease in the contribution by annual species to richness and cover from subcommunity 1.1 to 1.4. The changes in relative contribution to richness and cover were evident for the showy component of the annual species, but not for the non-showy component.
DISCUSSION
A clear distinction could be made between community 1 and the other three communities (Figure 2a). Community 1 was located on the farm Skilpad (in and adjacent to the park) at a fairly high altitude (620 m - 710 m), with subcommunities 1.1-1.3 on Land Type Ag94 and subcommunity 1.4 on Land Type Ib235. The subcommunities of community 1 were associated with time since abandonment, with subcommunity 1.1 being the youngest and subcommunity 1.4 the oldest (Figure 2b). The sample sites of subcommunity 1.1 were fairly closely clustered (Figure 2b), but as time progressed, the floristic composition of the fields became increasingly diverse indicating that there were divergent pathways along which succession on these old fields proceeded. Subcommunity 1.4 represented the oldest fields, however, these sites were also on a different land type to the other subcommunities, and this would have influenced the species assemblage patterns on the old fields in this subcommunity.
Although all the sample sites within community 1 have a similar ploughing history, divergence among the older abandoned fields could have been the result of different farming practices on the old fields, such as grazing history and intensity (Bonet 2004, Cadenasso et al. 2002), number of times ploughed, the various crops planted and the last crop planted (Bonet 2004; Myster & Pickett 1990), or the rainfall amount and temporal distribution in the year of abandonment (Myster & Pickett 1990; Pickett et al. 2008).
It is notable that old fields surveyed in the adjacent farmland were classified into the same subcommunities of community 1 as those in the park. However, most of the fields surveyed in the adjacent farmland had been abandoned for several decades and belonged to subcommunities 1.3 and 1.4. Therefore, for the first years after abandonment both the fields inside and outside the park would have been subjected to livestock grazing. It is only the more recently abandoned fields in the park that were subjected to a conservation land-use with a low grazing intensity after abandonment. These old fields were classified as subcommunity 1.2.
A comparison of the subcommunities of different ages in community 1 clearly indicated that secondary succession on these old fields proceeded very slowly. Such a slow rate of vegetation change is common in arid environmental conditions (Dean & Milton 1995; Otto et al. 2006). The vegetation on many old fields abandoned more than 50 years ago was still clearly visually distinguishable from the surrounding vegetation, although the vegetation on several sites in subcommunity 1.4 that had not been cultivated for more than 80 years could hardly be distinguished from the surrounding vegetation.
The remaining three communities (2, 3 and 4) were clearly separated from each other and from community 1 (Figure 2c). They were found in different vegetation types, on different land types and at different altitudes. Community 2 was characterised predominantly by Land Type Fb154 and occurred at intermediate altitudes, whereas community 3 occurred predominantly on Land Type Ib124, also at intermediate altitudes. Community 4 occurred in heuweltjieveld on Land Type Ag96 and at the lowest altitudes of all sites surveyed. Because rainfall is strongly influenced by the altitudinal gradient in this region, some of the differences between the communities could be ascribed to differences in available moisture.
Comparisons of the diversity among the four communities revealed no significant differences. Nevertheless, there were large differences in their potential to produce floral displays. Community 1 (subcommunities 1.1 through to 1.3 in particular) was the only community with the mass floral displays of Ursinia cakilefolia for which the Namaqua National Park has become world-renowned. The relative contribution of the perennial, annual (showy and non-showy) and geophyte life forms within community 2 and community 4 were similar. However, subcommunity 2.1 had a higher cover of showy annual plant species than subcommunity 2.2 and community 4. The most prominent annual species in subcommunity 2.1 were Felicia australis, Dimorphotheca sinuata and Grielum humifusum, whereas subcommunity 2.2 was dominated only by the annual Felicia australis, and community 4 had high cover values for Oncosiphon grandiflorum, Arctotis fastuosa and Grielum humifusum. Overall, community 3 offered the least to tourists by way of mass flowering displays. This community had a high cover of perennial species, largely dominated by Galenia africana, Calobota halenbergensis and Galenia fruticosa, and most of the annuals occurring in the community were inconspicuous. The rocky soils (Land Type Ib124) in this community could have contributed to the low cover of annual species.
Many different relationships between species richness and age since abandonment have been reported. Theoretically, an asymptotic relationship is predicted under environmentally limiting conditions (Peet 1992), although Bonet (2004) found a parabolic relationship in an arid region in southern Spain, whereas Otto et al. (2006) reported a linear increase under arid conditions. A parabolic relationship is believed to be typical of the vegetation in the Mediterranean Basin (Bonet 2004; Debussche et al. 1996,) and could therefore have been expected in Namaqualand with its Mediterranean climate, with winter rainfall. However, total and perennial species richness did not differ significantly among the subcommunities of community 1, which reflect a temporal gradient from less than 5 years to about 80 years since cultivation. The relative richness of annual species, and in particular of the showy annual species, showed a decrease with time since abandonment (Figure 3a).
Within community 1 it was clear that while the relative contribution of the perennial species to vegetation cover increased with time since abandonment, the relative contribution made by showy annual species to vegetation cover declined with age since abandonment (Figure 3b). These results support the general contention that spectacular mass floral displays rely on regular disturbance (Van Rooyen 2002), such as experienced in subcommunity 1.1. Whether the mass floral displays are associated with a loss in plant diversity depends on how diversity is measured. Subcommunity 1.1, with exceptional mass displays, did not have a lower total species richness or richness of perennial species than the other subcommunities of community 1, although the Shannon-Wiener diversity index indicated a lower diversity in the perennial component in subcommunity 1.1. In contrast, both species richness as well as the Shannon-Wiener index indicated a higher diversity for the annual component in subcommunity 1.1 than in subcommunity 1.4.
Understanding the process of natural vegetation change and diversity relationships of old fields in the Namaqua National Park is essential to evaluate the conservation outcomes of the park. However, the current study is not only relevant for the park. Worldwide, there is an increasing trend of land abandonment as a result of environmental and socio-economic changes (Cramer & Hobbs 2007; Cramer et al. 2007). A similar trend can be seen in Namaqualand (Hoffman & Rohde 2007). Consequently, many abandoned croplands lie scattered throughout the Namaqualand landscape. The insight gained into vegetation dynamics of old fields in the Namaqua National Park could provide a framework (Bartha et al. 2003; Pickett et al. 2008) for the investigation of restoration opportunities of old fields in the larger Namaqualand region.
CONCLUSION
The aim of this study was to classify the plant communities on the abandoned fields across different vegetation types in the Namaqua National Park and to investigate the impact of time since abandonment on plant diversity and mass flowering displays. The knowledge gained in this study provides insight that could aid future planning and effective management of the many old fields scattered over this extensive conservation area. Old fields located on the Skilpad section belong to a single community and produce the characteristic mass floral displays of Ursinia cakilefolia for which the Namaqua National Park has become world-renowned. Old fields in other parts of the park belong to other communities, and will not produce the same spectacular Ursinia cakilefolia dominated displays as the old fields on Skilpad but should contribute to the appeal of the park by increasing the diversity of the springtime displays of wild flowers. On the Skilpad section the best flowering displays were associated with the old fields that were tilled approximately every 4 years (subcommunity 1.1), although a variety of showy annual species were still commonly found on fields abandoned about 20 years ago. To maintain a flower display for tourists this type of management intervention appears to be essential. Apart from the old fields along the circular tourist route on the Skilpad section, all other old fields in the park should not be disturbed, but should be allowed to recover naturally.
Research significance
Understanding the process of natural vegetation change and diversity relationships of old fields in the Namaqua National Park is essential to evaluate the conservation outcomes of the park. The insight gained into vegetation dynamics of old fields in the park could, furthermore, provide a framework for the investigation of restoration opportunities of old fields in the larger Namaqualand region.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the support received from the German Federal Ministry of Education and Research (BMBF) through the BIOTA South Project, and the National Research Foundation under grant number 61277. SANParks and in particular, Dr H. Bezuidenhout and Dr H. Hendricks are thanked for permission to conduct research on the Namaqua National Park. We also thank various students from the Department of Plant Science, University of Pretoria, and Annelise le Roux (Succulent Karoo Knowledge Centre) for field assistance.
Editor: Llewellyn C. Foxcroft
REFERENCES
AGIS, 2007, Agricultural geo-referenced information system, viewed 10 April 2010, from www.agis.agric.za [ Links ]
Bartha, S., Meiners, S.T., Pickett, S.T.A. & Cadenasso, M.L., 2003, 'Plant colonization window in a mesic old field succession', Applied Vegetation Science 6, 205-212. [ Links ]
Bonet, A., 2004, 'Secondary succession of semi-arid Mediterranean old-fields in south-eastern Spain: Insights for conservation and restoration of degraded lands', Journal of Arid Environments 56, 213-233. [ Links ]
Cadenasso, M., Pickett, S.T.A. & Morin, P.J., 2002, 'Experimental test of the role of mammalian herbivores on old field succession: Community structure and seedling survival', Journal of the Torrey Botanical Society 129, 228-237. [ Links ]
Conservation International, n.d., Biodiversity hotspots, viewed 10 April 2010, from http://www.biodiversityhotspots.org [ Links ]
Cornell, D.H., Thomas, R.J., Moen, H.F.G., Reid, D.L., Moore J.M. & Gibson, R.L., 2006, 'The Namaqua-Natal Province', in M.R. Johnson, C.R. Anhaeusser & R.J. Thomas (eds.), The geology of South Africa, pp. 325-379, Council for Geoscience, Pretoria. [ Links ]
Cowling, R.M., Esler, K.J. & Rundel, P.W., 1999, 'Namaqualand, South Africa - An overview of a unique winter-rainfall desert ecosystem', Plant Ecology 142, 3-21. [ Links ]
Cramer, V.A. & Hobbs, R.J. (eds.), 2007, Old fields. Dynamics and restoration of abandoned farmland, Island Press, Washington. [ Links ]
Cramer, V.A., Hobbs, R.J. & Standish, R.J., 2007, 'What's new about old fields? Land abandonment and ecosystem assembly', Trends in Ecology and Evolution 23, 104-112. [ Links ]
Critical Ecosystem Partnership Fund, 2003, Ecosystem Profile: The Succulent Karoo hotspot, Namibia and South Africa, Critical Ecosystem Partnership Fund report. [ Links ]
Dean, W.R. & Milton, S.J., 1995, 'Plant and invertebrate assemblages on old fields in the arid southern Karoo, South Africa', African Journal of Ecology 33, 1-13. [ Links ]
Debussche, M., José, E., Lepart, J., Houssard, C., & Lavorel, S., 1996, 'Changes in Mediterranean plant succession: Old-fields revisited', Journal of Vegetation Science 7, 519-526. [ Links ]
Desmet, P.G., 2007, 'Namaqualand - A brief overview of the physical and floristic environment', Journal of Arid Environments 70, 570-587. [ Links ]
Du Plessis, H.M., 1987, 'Land Types of the maps 2816 Alexander Bay, 2818 Warmbad, 2916 Springbok, 2918 Pofadder, 3017 Garies, 3018 Loeriesfontein', Memoirs on the Agricultural Natural Resources of South Africa 9, 1-538. [ Links ]
Francis, M.L., Fey, M.V., Prinsloo, H.P., Ellis, F., Mills, A.J. & Medinski, T.V., 2007, Soils of Namaqualand: Compensations for aridity, Journal of Arid Environments 70, 588-603. [ Links ]
Hennekens, S.M. & Schaminée, J.H.J., 2001, 'TURBOVEG, a comprehensive data base management system for vegetation data', Journal of Vegetation Science 12, 589-591. [ Links ]
Hill, M.O., 1979, TWINSPAN - A FORTRAN program for arranging multivariate data in an ordered two-way table by classification of the individuals and attributes, Ecology & Systematics, Cornell University Ithaca, New York. [ Links ]
Hoffman, M.T. & Rohde, R.F., 2007, 'From pastoralism to tourism. The historical impact of changing land use practices in Namaqualand', Journal of Arid Environments 70, 641-658. [ Links ]
James, I., Munro, A., Hoffman, T., O'Farrell, P.J. & Smart, R. 2007. The economic value of flower tourism at the Namaqua National Park, South Africa, South African Journal of Economic and Management Sciences 10, 442-456. [ Links ]
McCune, B. & Grace, J.B., 2002, Analysis of Ecological Communities, MjM Software Design, Gleneden Beach, Oregon. [ Links ]
Mittermeier R.A., Myers, N., Robles Gil, P. & Mittermeier, C.G., 2000, Hotspots: Earth's biologically richest and most endangered terrestrial ecoregions, CEMEX, Mexico. [ Links ]
Mucina, L., Jürgens, N., Le Roux, A., Rutherford, M.C., Schmiedel, U., Esler, K.J. et al., 2006, 'Succulent Karoo Biome', in L. Mucina & M.C. Rutherford (eds.), The vegetation of South Africa, Lesotho and Swaziland, Strelitzia 19, pp. 220-299, South African National Biodiversity Institute, Pretoria. [ Links ]
Myers, N., Mittermeier, R.A., Mittermeier, C.G., De Fonseca, G.A.B. & Kent, J., 2000, 'Biodiversity hotspots for conservation priorities', Nature 403, 853-858. [ Links ]
Myster, R.W. & Pickett, S.T.A., 1990, 'Initial conditions, history and successional pathways in ten contrasting old fields', American Midland Naturalist 124, 231-238. [ Links ]
Otto, R., Krüsi, B.O., Burga, C.A., Fernández-Palacios, J.M., 2006, 'Old-field succession along a precipitation gradient in the semi-arid coastal region of Tenerife', Journal of Arid Environments 65, 156-178. [ Links ]
Peet, R.K., 1992, 'Community structure and persistence', in D.C. Glenn-Lewin, R.K. Peet & T.T. Veblen (eds.) Plant succession: Theory and prediction, pp. 103-151, Chapman & Hall, New York. [ Links ]
Petersen, A., 2008, 'Pedodiversity of southern African drylands', PhD thesis, Institute of Soil Science, University of Hamburg. [ Links ]
Pickett, S.T.A., Cadenasso, M.L. & Meiners, S.J., 2008, 'Ever since Clements: from succession to vegetation dynamics and understanding to intervention', Applied Vegetation Science 12, 9-21. [ Links ]
Podani, J., 2001, SYN-TAX 2000 Computer programs for data analysis in ecology and systematic, Scientia publishing, Budapest. [ Links ]
South African National Parks, 2010, 'Namaqua National Park: Park Management Plan', February 2010 (draft), SANParks, Pretoria. [ Links ]
Van der Maarel, E., 2007, 'Transformation of cover-abundance values for appropriate numerical treatment - Alternatives to the proposals by Podani', Journal of Vegetation Science 18, 767-770. [ Links ]
Van Rooyen, M.W., Theron G.K. & Van Rooyen, N., 1996, 'Skilpad Wildflower Reserve: A flower-lover's paradise', Veld and Flora 4, 40-42. [ Links ]
Van Rooyen, M.W., 1999. Functional aspects of short-lived plants, in W.R. Dean & S.J. Milton (eds.), The Karoo: Ecological patterns and processes, pp. 107-122, Cambridge University Press, Cambridge. [ Links ]
Van Rooyen, M.W., 2002, 'Management of the old field vegetation in the Namaqua National Park, South Africa: conflicting demands of conservation and tourism', Geographical Journal 168, 211-223. [ Links ]
Werger, M.J.A., 1974, 'On concepts and techniques applied in the Zürich-Montpellier method of vegetation survey', Bothalia 11, 309-323. [ Links ]
 Correspondence to:
Correspondence to:
Gretel van Rooyen
Department of Plant Science, University of Pretoria
Pretoria 0002, South Africa
email: gretel.vanrooyen@up.ac.za
Received: 18 May 2010
Accepted: 04 Aug. 2010
Published: 22 Nov. 2010
This article is available at: http://www.koedoe.co.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}