










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkKoedoe
versión On-line ISSN 2071-0771
versión impresa ISSN 0075-6458
Koedoe vol.51 no.1 Pretoria ene. 2009
ORIGINAL RESEARCH
Spiders and scorpions (Arachnida: Araneae, Scorpiones) of the Nylsvley Nature Reserve, South Africa
Ansie S. Dippenaar-SchoemanI, II; Annette van den BergI, II; Lorenzo PrendiniIII
IARC-Plant Protection Research Institute, Queenswood, South Africa
IIDepartment of Entomology and Zoology, University of Pretoria, South Africa
IIIDivision of Invertebrate Zoology, American Museum of Natural History, New York
ABSTRACT
Among other activities, the South African National Survey of Arachnida (SANSA) aims to survey the biodiversity of arachnids in protected areas of South Africa. The study presented here documents the diversity of spiders and scorpions collected from the Nylsvley Nature Reserve (NNR), South Africa over a 30-year period. The spider fauna of NNR contains 175 species (7.5% of the total recorded in South Africa), in 131 genera and 37 families. Thomisidae is the most diverse spider family in the reserve, with 33 species (18.9% of the total), followed by Salticidae, with 20 species (11.4%), and Araneidae, with 18 species (10.3%). The majority of species (125) are wandering spiders (71.4%), whereas 50 species (28.6%) build webs. Wandering ground-dwelling spiders comprise 52 species, whereas 73 wandering species have been collected from the vegetation. A total of 158 species are new records for the reserve and Oxyopes tuberculatus Lessert, 1915 is newly recorded for South Africa. Six spider species may be new to science. The scorpion fauna of NNR comprises five species (5% of the total recorded in South Africa) in three genera and two families. Buthidae are more diverse in the reserve, with four species and two genera represented. The scorpion fauna of the reserve includes two fossorial and three epigeic species, representing five ecomorphotypes: semi-psammophilous, pelophilous, lithophilous, corticolous and lapidicolous. Five additional scorpion species may be recorded if the reserve is sampled more intensively using appropriate techniques.
Keywords: Biodiversity; faunistics; Savanna Biome; South African National Survey of Arachnida
INTRODUCTION
The South African National Survey of Arachnida (SANSA) was initiated in 1997 to survey the arachnid diversity of South Africa (Dippenaar-Schoeman & Craemer 2000). SANSA encompasses several projects, including surveys and inventories of arachnids that are protected in parks and reserves; surveys and inventories of arachnid diversity in the floral biomes; and checklists of arachnid species for each province in South Africa. Although arachnids constitute an abundant and diverse group of invertebrates, knowledge of their diversity in South Africa remains poor (Dippenaar-Schoeman 2002).
The aim of the study presented here was to survey the spider and scorpion fauna of the Nylsvley Nature Reserve (NNR), situated in the Limpopo Province of South Africa. The reserve lies in the upper reaches of the Nyl River on the Nyl floodplain, the largest inland floodplain in South Africa, which was recognised as a Ramsar site in 1998.
The Savanna Biome, a vegetation type where trees and grass are equally important components, covers approximately half of Africa's land surface and about 35% of South Africa. NNR falls entirely within the biome and is one of the most intensively studied savanna sites in the world. More than 100 scientific papers and reports, many postgraduate degrees and several books were produced during the South African Savanna Biome project, conducted at NNR from 1974 to 1990. The aim of this project was to develop the understanding necessary to predict changes in the biome's stability (Scholes & Walker, 1993). However, little information is available for the arachnids. The only published data on the spiders of NNR are a survey of spiders from abandoned mammal holes (Heidger, 1988) and notes on the behaviour of a trapdoor spider, Ancylotrypa brevicornis (Hewitt 1919; Leroy & Leroy 2005). The study presented in this contribution is the eighth survey of the arachnid fauna of the Savanna Biome in South Africa and the first arachnid inventory for the NNR. Other surveys of arachnids in the Savanna Biome were undertaken at Roodeplaat Dam Nature Reserve (Dippenaar-Schoeman, van den Berg & van den Berg 1989), Makalali Nature Reserve (Whitmore, Slotow, Crouch & Dippenaar-Schoeman 2002; Druce, Hamer, Slotow & Prendini, 2004), Western Soutpansberg (Foord, Dippenaar-Schoeman & Van der Merwe 2002; in press), Kruger National Park (Dippenaar-Schoeman & Leroy 2003), the Inselberg at Polokwane (Pietersburg) (Modiba, Dippenaar & Dippenaar-Schoeman 2005), Ndumo Game Reserve (Haddad, Dippenaar-Schoeman & Wesolowska 2006) and Polokwane Nature Reserve (Dippenaar, Modiba, Khoza & Dippenaar-Schoeman 2008).
This contribution does not present the results of a quantitative survey, but instead summarises data from collections of spiders and scorpions made in the NNR over a period of 30 years, including specimens collected during the South African Savanna Biome project (Scholes & Walker 1993). Although this survey may not reflect the true diversity and species richness of spiders and scorpions in the reserve, it nonetheless provides an estimate of the species presently protected within its boundaries.
STUDY AREA
Nylsvley Nature Reserve (24º39'S 28º42'E) is situated 12 km south of Mole Mole (Nylstroom) in the Limpopo Province of South Africa. The altitude of NNR ranges between 1 080 m and 1 154 m above sea level, with an average altitude of 1 100 m. The reserve, which is located in the upper reaches of the Nyl River floodplain, comprises 3 970 ha, of which 500 ha (10%) falls within the NNR. The Nyl River floodplain, the largest inland wetland in South Africa (Noble & Hemens 1978), extends from Middelfontein, west of Mole Mole in the south-west, to Moorddrift, near Molopane (Potgietersrus) in the north-east. NNR is situated in the summer rainfall region, receives rainfall during the hot summer months and experience cool, dry winters. The rainfall is variable: the 69-year mean annual rainfall at NNR is 623 mm, with an annual coefficient of variation of 24%.
The mean annual temperature is 19ºC (Scholes & Walker 1993). The maximum daily temperature ranges from a mean of 29ºC in December/January to 21ºC in June/July, whereas the minimum daily temperature varies from 17ºC in December/January to 4ºC in June/July.
The Nyl River floodplain comprises short grassy plains, reed beds, stands of long rice grass, open water patches, marshes, acacia (fineleaf) woodland and broadleaf woodland. Approximately 600 plant species have been recorded in the NNR, and Scholes and Walker (1993) have distinguished nine vegetation types: Burkea africana savanna; Diplorhynchus condylocarpon savanna; Combretum savanna; Acacia tortilis savanna; old village sites; Acacia karroo savanna; floodplain grasslands; grasslands on vertic soils; and seepline grassland.
These vegetation types may be further grouped into two broad categories (Low & Rebelo 1996), namely mixed bushveld and clay thorn bushveld.
METHODS
Collection techniques
During the South African Savanna Biome project (1974 to 1990), spiders were collected from grass with sweepnets and from trees by beating. This material was donated to the National Collection of Arachnida, where it was sorted and identified. Additional sporadic collecting was undertaken in the NNR from 1985 to 2005. During this period, spiders were collected by hand (ground and plant search, turning rocks, peeling bark, sifting leaf litter) or by using a sweepnet for grass and a beating tray for low shrubs. Scorpions were collected by hand (mainly by turning rocks and peeling bark).
Material examined
The spiders examined during the course of the study were identified by the first author and are deposited in the National Collection of Arachnida (NCA) at the ARC-Plant Protection Research Institute, Pretoria. The scorpions were identified by the third author and are deposited in the NCA, the American Museum of Natural History in New York, and the Transvaal Museum in Pretoria.
The lack of taxonomic research on certain spider families (e.g. Lycosidae and Theridiidae) in southern Africa prevented the identification of some specimens to species. In some families, only immature specimens were collected and these were impossible to identify to species level.
Ecology
A guild is a group of species that potentially compete for jointly exploited limited resources (Uetz, Halaj & Cady 1999). Because most spiders live in a defined environment with limitations imposed by both abiotic and biotic factors, species can be grouped into guilds based on information about their habitat preferences and predation methods. Two main guilds of spiders were recognised in the present study, i.e. wandering spiders (W) and web builders (WB), with further subdivisions based on microhabitat and general behaviour, as defined by Dippenaar-Schoeman and Leroy (2003). Guilds of scorpions were defined and subdivided on the basis of the ecomorphotypes defined by Prendini (2001, 2005).
RESULTS AND DISCUSSION
Spiders
Diversity
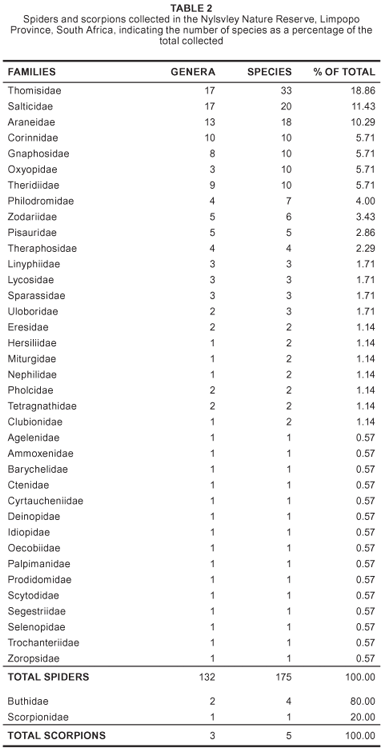
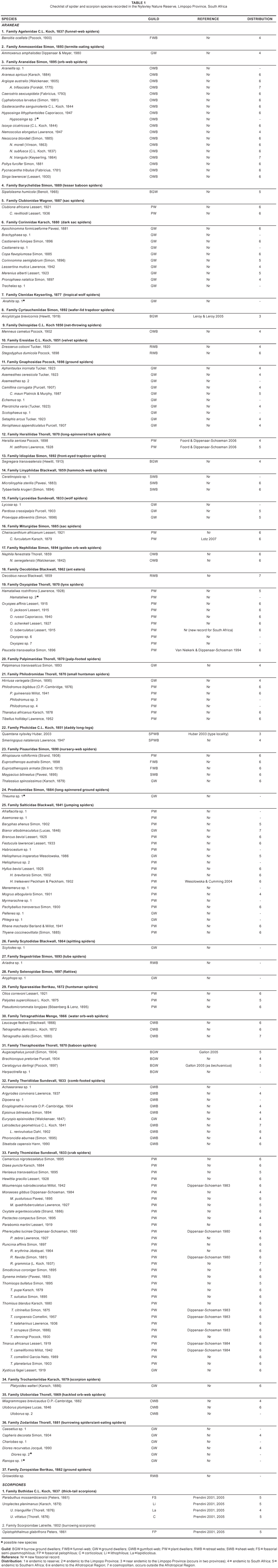
A total of 175 spider species (7.5% of the total recorded in South Africa), representing 132 genera and 37 families (tables 1 and 2) were collected in the NNR. Thomisidae was the most diverse spider family, with 33 species (18.9% of the total), followed by Salticidae, with 20 species (11.4%), Araneidae, with 18 species (10.3%), and Corinnidae, Gnaphosidae, Oxyopidae and Theridiidae, with 10 (5.7%) species each. Fifteen families were each represented by a single species (Table 2) and 159 species are new records for the reserve (Table 1). Oxyopes tuberculatus Lessert, 1915 is newly recorded for South Africa. The pholcid, Quamtana nylsvley Huber, 2003, was originally described from the reserve (Huber, 2003). Six species of the genera Anahita (Ctenidae), Diores and Ranops (Zodariidae), Hamataliwa (Oxyopidae), Hypsosinga (Araneidae) and Theuma (Prodidomidae), may be new to science (Table 1).
Guilds
Although the majority of the spiders collected in the NNR, namely 125 species (71.4%) representing 24 families, are wanderers, 50 species (28.6%) in 13 families build webs. A total of 52 species (30%) in 22 families live on the ground. Seven species (4%) in four families of ground-dwelling spiders are fossorial, whereas 45 species (26%) in 14 families are free-living. Seventy-three species (42%) in 12 families inhabit vegetation, of which 36 species (20.8%) in 11 families construct webs to catch prey in the grass and herbaceous layer.
Fossorial species: Seven species of the suborder Mygalomorphae (baboon and trapdoor spiders) inhabit silk-lined burrows and are presently protected in most provinces of South Africa. The baboon spider families Barychelidae and Theraphosidae are represented by one species, Sipalolasma humicola (Benoit, 1965), and four species, Augacephalus junodi (Simon, 1904), Brachionopus pretoriae Purcell, 1904, Ceratogyrus darlingi Pocock, 1897 and an unidentified species of Harpactirella (Gallon 2005) respectively. Two trapdoor spider families are known from the NNR: Segregara transvaalensis (Hewitt, 1913) (Idiopidae) and Ancylotrypa brevicornis (Hewitt, 1919) (Cyrtaucheniidae). Observations of the burrow-constructing behaviour of A. brevicornis showed that, in addition to constructing a thin wafer-lid trapdoor, these spiders use a hard, spherical plug (or marble) made of soil particles, held together by silk, to close the burrow entrance (Leroy & Leroy 2005).
Free-living ground species: Of the 45 species in 14 families of free-living ground spiders collected in the NNR, Gnaphosidae and Corinnidae, each represented by 10 species, are the most abundant, followed by Zodariidae, with six species, and Lycosidae, with three species.
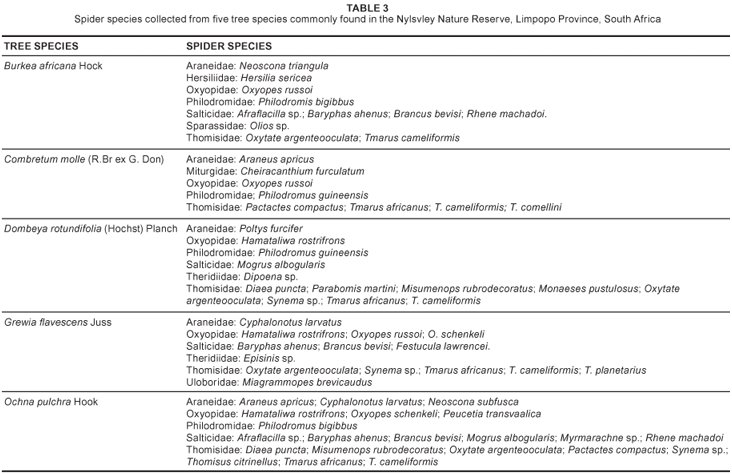
Plant-dwelling species: Thirty-six species (20.8%) in 10 families are commonly found on the five dominant tree species in the NNR (Table 3). Eleven species (seven families) occur on Burkea africana Hock (wild syringa), eight species (five families) on Combretum molle (R.Br ex G. Don) (velvet bush willow), 13 species (six families) on Dombeya rotundifolia (Hochst) Planch. (wild pear), 14 species (six families) on Grewia flavescens Juss. (raisin bush), and 21 species (five families) on Ochna pulchra Hook (peeling plane). Species that live permanently on tree bark include the long-spinnered bark spiders, Hersilia sericea Pocock, 1898 and H. setifrons Lawrence, 1927 (Foord & Dippenaar-Schoeman, 2006). Four Tmarus spp. (Thomisidae) and Oxytate argenteooculata (Strand, 1886) (Thomisidae) are also exclusively arboreal. Most of the araneid orb-web species found on vegetation are nocturnal and remove their webs early each morning.
Six araneid species, Araneus apricus (Karsch 1884), Caerostris sexcuspidata (Fabricius 1793), Cyphalonotus larvatus (Simon 1881), Neoscona subfusca (C.L. Koch 1837), N. triangula (Keyserling 1864) and Poltys furcifer Simon, 1881, were collected from trees to which they retreat when inactive during the day.
The NNR has a rich fauna of graminicolous spider species, most with elongated bodies resembling grass stems. These include the Runcinia spp. (Thomisidae), Tibellus hollidayi Lawrence, 1952 (Philodromidae) and Pseudomicrommata longipes (Bösenberg & Lenz, 1895) (Sparassidae). Other graminicolous spider species belong to the families Oxyopidae, Salticidae and Thomisidae.
Web-building species: The orb-web spiders, comprising 25 species in the families Araneidae (18 species), Nephilidae (two), Tetragnathidae (three) and Uloboridae (three), are the most diverse group recorded. Nephilidae and Tetragnathidae are often seen in their large webs during the day. Species of Tetragnathidae (Tetragnatha and Leucauge) are associated with wetlands and found commonly in the NNR.
Ten species construct webs on or close to the soil surface. Benoitia ocellata (Pocock, 1900) an agelenid, is often observed in the NNR, especially early in the mornings when dew collects on the sheet part of its web. The very large Euprosthenops australis Simon, 1898 (Pisauridae) is another common spider, which constructs very large funnel webs next to trees, the funnel usually originating from an abandoned termite nest or mammal burrow. The retreat webs of Eresidae (Dresserus colsoni Tucker, 1920), Oecobiidae, Segestriidae and Zoropsidae are constructed under rocks and plant debris, whereas the sheet webs of Linyphiidae and gumfoot webs of Theridiidae are constructed in grass close to the ground.
The space webs of Pholcidae are constructed mainly in abandoned mammal holes (Heidger, 1988). Stegodyphus dumicola Pocock, 1898 is a community web spider that constructs large nest-like retreats containing numerous spiders on vegetation.
Scorpions
Diversity
Five scorpion species (5% of the total recorded in South Africa), representing three genera and two families, have been collected in the NNR (Table 1). Family Buthidae is represented by two genera and three species, whereas Scorpionidae is represented by a single genus and species.
Guilds
The five scorpion species recorded in the NNR can be grouped into two different guilds: fossorial or burrowing species (40% of the species), and epigean species, which do not construct burrows (60%). Each of these guilds can be further subdivided into distinct ecomorphotypes.
Fossorial species: Two of the scorpion species collected in the NNR are adapted for constructing burrows and spend most of their time below the soil surface, i.e. they are fossorial. These species represent two distinct ecomorphotypes, associated with different substrata: semi-psammophilous and pelophilous species (Prendini 2001).
Parabuthus mossambicensis (Peters 1861) is a semi-psammophilous species that constructs burrows in semi-consolidated sand to sandy-loam substrata. Burrows are constructed in open ground using the thickened metasoma to loosen the soil and the anterior two pairs of legs to scrape and rake soil out of the burrow.
This species displays several ecomorphological adaptations to increase locomotor and burrowing efficiency in soft substrata, including carinae and spiniform processes on the metasoma and comb-like rows of long macrosetae on the prolateral and retrolateral margins of the tibia and basitarsi of the first and second pairs of legs.
Opistophthalmus glabrifrons Peters, 1861 is a pelophilous species that constructs burrows in hard sandy-loam and clay substrata. Burrows are constructed in open ground or under stones using the enlarged chelicerae to loosen the soil, and the anterior two pairs of legs to scrape and rake soil out of the burrow; the metasoma is also used for tail-scraping operations. This species displays several ecomorphological adaptations to increase burrowing efficiency in hard substrata, including short, robust legs and telotarsal ungues; stout, spiniform macrosetae distributed laterally and distally on the basitarsi; and carinae and spiniform processes on the metasoma.
Epigean species: Three (60%) of the scorpion species recorded in the NNR are found at or above the soil surface and are unable to construct burrows, i.e. they are epigean. These species represent three distinct ecomorphotypes associated with different substrata: lithophilous, corticolous and lapidicolous species (Prendini 2001).
Uroplectes planimanus (Karsch 1879) is a lithophilous species adapted to life in the narrow cracks and crevices of rocks and under rocks resting on bedrock. This species is characterised by moderate dorsoventral compression; elongation of the metasoma and pedipalps; and stout macrosetae on the telotarsi, operating in conjunction with curved telotarsal ungues to provide grip on rock surfaces.
These specialised adaptations facilitate rapid locomotion in any spatial plane of their rock habitats, but hinder locomotion on other substrata. These scorpions are therefore restricted to weathered rock outcrops.
Uroplectes vittatus (Thorell 1876) is an obligate corticolous species, found in holes or under the loose bark of old or dead trees (especially acacias), often several metres above the ground. This species exhibits a few ecomorphological adaptations, for example moderate dorsoventral compression, elongation of the metasoma and pedipalps, and well-developed telotarsal ungues.
Uroplectes triangulifer (Thorell 1876) is a lapidicolous species that shelters under loose stones, wood or debris at ground level, and displays few ecomorphological adaptations for this generalist lifestyle.
Species not recorded: Up to five additional scorpion species, representing five genera and two families, may be recorded in the NNR if the scorpion fauna is more intensively sampled using appropriate techniques, e.g. pitfall trapping and night collecting by ultraviolet detection. The NNR falls within the known distributional range of these widespread savanna scorpion species, and suitable habitat occurs within its boundaries, suggesting that at least some of them may be present. The species include two buthids, the semi-psammophilous Parabuthus transvaalicus Purcell, 1899 and lapidicolous Uroplectes carinatus (Pocock 1890), and three liochelids, the pelophilous Cheloctonus jonesii Pocock, 1892, lithophilous Hadogenes sp., and corticolous Opisthacanthus asper (Peters 1861).
CONCLUSION
Arachnids form an important component of healthy ecosystems. Any approach to conservation must consider the composition of the arachnid fauna. Inventories with resulting checklists provide valuable baseline data about the species present in reserves and are the first step towards a better understanding of the fauna.
Preliminary investigations of the biodiversity of arachnids in South Africa have highlighted the lack of baseline data on the ecology and diversity of most arachnid orders (Dippenaar-Schoeman 2002). The survey presented here forms part of the South African National Survey of Arachnida (SANSA) and the data gathered will be used in the Savanna Biome database.
The NNR has a rich fauna of arachnid species (180) and, although this contribution reports on sporadic collecting and probably represents only a subset of the arachnids present, we hope it will stimulate further research on the arachnids of the NNR and Savanna Biome.
ACKNOWLEDGEMENTS
We thank Gherda Ferreira, who collected arachnids during the Savanna Biome Survey, the Spider Club of Southern Africa, and the Friends of Nylsvley for additional collecting. This work forms part of the South African National Survey of Arachnida (SANSA), which is coordinated by the South African National Biodiversity Institute and the Agricultural Research Council.
REFERENCES
Dippenaar, S.M., Modiba, M.A., Khoza, T.T. & Dippenaar-Schoeman, A.S. 2008. A checklist of spiders (Arachnida, Araneae) of the Polokwane Nature Reserve, Limpopo Province, South Africa. Koedoe, 50, 10–17. [ Links ]
Dippenaar-Schoeman, A.S. 1980. The crab-spiders of southern Africa (Araneae: Thomisidae). 1. The genus Runcinia Simon, 1875. Journal of the Entomological Society of Southern Africa, 43, 303–326. [ Links ]
Dippenaar-Schoeman, A.S. 1983. The spider genera Misumena, Misumenops, Runcinia and Thomisus (Araneae: Thomisidae) of southern Africa. Entomology Memoirs of the Department of Agricultural Technical Services, Republic South Africa, 55, 1–66. [ Links ]
Dippenaar-Schoeman, A.S. 1984. The crab-spiders of southern Africa (Araneae: Thomisidae). 4. The genus Monaeses Thorell, 1869. Phytophylactica, 16, 101–116. [ Links ]
Dippenaar Schoeman, A.S. 2002. Status of South African Arachnida Fauna. Proceedings of the symposium on the Status of South African species organized by the Endangered Wildlife Trust (EWT) of South Africa, Rosebank, 4–7 September 2002. [ Links ]
Dippenaar-Schoeman, A.S. & Craemer, C. 2000. The South African National Survey of Arachnida. Plant Protection News, 56, 11–12. [ Links ]
Dippenaar-Schoeman, A.S. &. Leroy, A. 2003. A check list of the spiders of the Kruger National Park, South Africa (Arachnida: Araneae). Koedoe, 46, 91–100. [ Links ]
Dippenaar-Schoeman, A.S., Van den Berg, A.M. & Van den Berg, A. 1989. Species composition and relative seasonal abundance of spiders from the field and tree layers of the Roodeplaat Dam Nature Reserve. Koedoe, 32, 51–60. [ Links ]
Druce, D., Hamer, M., Slotow, R. & Prendini, L. 2004. Checklist of millipedes (Diplopoda), centipedes (Chilopoda) and scorpions (Arachnida: Scorpionida) from a savanna ecosystem, Limpopo Province, South Africa. African Invertebrates, 45, 315–322. [ Links ]
Foord, S.H. & Dippenaar-Schoeman, A.S. 2006. A revision of the Afrotropical species of Hersilia Audouin (Araneae: Hersiliidae). Zootaxa, 1347, 1–92. [ Links ]
Foord, S.H., Dippenaar-Schoeman, A.S. & Van der Merwe, M. 2002. A check list of the spider fauna of the Western Soutpansberg, South Africa (Arachnida: Araneae). Koedoe, 45, 35–43. [ Links ]
Foord, S.H., Mafadza, M.M., Dippenaar-Schoeman, A.S. & Van Rensburg, B.J. in press. Micro-scale heterogeneity of spiders (Arachnida: Araneae) in the Soutpansberg, South Africa: a comparative survey and inventory in representative habitats. African Zoology. [ Links ]
Gallon, R.C. 2005. A new species of theraphosid spider from Southern Africa (Araneae, Theraphosidae, Harpactirinae) with distributional notes on other harpactirines. Bulletin of the British Arachnological Society, 13, 179–184. [ Links ]
Haddad, C.R., Dippenaar-Schoeman, A.S. & Wesolowska, W. 2006. A checklist of the non-acarine arachnids (Chelicerata: Arachnida) of the Ndumo Game Reserve, Maputaland, South Africa. Koedoe, 49, 1–22. [ Links ]
Heidger, C. 1988. Ecology of spiders inhabiting abandoned mammal burrows in South African savanna. Oecologia, 76, 303–306. [ Links ]
Huber, B.A. 2003. Southern African pholcid spiders: revision and cladistic analysis of Quamtana gen. nov. and Spermophora Hentz (Araneae: Pholcidae), with notes on male-female covariation. Zoological Journal of the Linnean Society, 139, 77–527. [ Links ]
Leroy, A. & Leroy, J. 2005. Notes on the natural history of a trapdoor spider Ancylotrypa Simon (Araneae, Cyrtaucheniidae) that constructs a spherical burrow plug. Journal of Arachnology, 33, 558–562. [ Links ]
Lotz, L.N. 2007. The genus Cheiracanthium (Araneae: Miturgidae) in the Afrotropical Region. 1. Revision of known species. Navorsinge van die Nasionale Museum, Bloemfontein, 23, 1–76. [ Links ]
Low, A.B. & Rebelo, H.G. 1996. Vegetation of South Africa, Lesotho and Swaziland. Pretoria: Department of Environment Affairs and Tourism. [ Links ]
Modiba, M.A., Dippenaar, S.M. & Dippenaar-Schoeman, A.S. 2005. A checklist of spiders from Sovenga Hill, an inselberg in the Savanna Biome, Limpopo Province, South Africa (Arachnida: Araneae). Koedoe, 48, 109–115. [ Links ]
Noble, R.G. & Hemens, J. 1978. Inland water ecosystems in South Africa – a review of research needs. South African National Scientific Programmes Report, 34, 1–150. [ Links ]
Prendini, L. 2001. Substratum specialization and speciation in southern African scorpions: the Effect Hypothesis revisited. In: Fet, V. & Selden, P.A. (eds.). Scorpions 2001. In Memoriam Gary A. Polis. Burnham Beeches, Bucks, UK, British Arachnological Society, pp. 113–138. [ Links ]
Prendini, L. 2005. Scorpion diversity and distribution in southern Africa: pattern and process. In: Huber, B.A., Sinclair, B.J. & Lampe, K.-H. (eds.). African biodiversity: molecules, organisms, ecosystems. Proceedings of the 5th International Symposium on Tropical Biology, Museum Alexander Koenig, Bonn. New York: Springer Verlag, pp. 25–68. [ Links ]
Scholes, R.J. & Walker, B.H. 1993. An African savanna, synthesis of the Nylsvley study. Cambridge: Cambridge University Press. [ Links ]
Uetz, G.W., Halaj, J. & Cady, A.B. 1999. Guild structure of spiders in major crops. Journal of Arachnology, 27, 270–280. [ Links ]
Van Niekerk, P. & Dippenaar-Schoeman, A.S. 1994. A revision of the Afrotropical species of Peucetia (Araneae: Oxyopidae). Entomology Memoir (Department of Agriculture), 89, 1–50. [ Links ]
Wesolowska, W. & Cumming, M.S. 2004. A redescription and natural history of Hyllus treleaveni Peckham et Peckham, 1902, the largest jumping spider in Africa (Araneae: Salticidae). Annales Zoologici Fennici, 54, 579–586. [ Links ]
Whitmore, C., Slotow, R., Crouch, T.E. & Dippenaar-Schoeman, A.S. 2002. Diversity of spiders (Araneae) in a savanna reserve, Northern Province, South Africa. Journal of Arachnology, 30, 344–356. [ Links ]
 Correspondence to:
Correspondence to:
Ansie S. Dippenaar-Schoeman
ARC-Plant Protection Research Institute
Private Bag X134
Queenswood, 0121, South Africa
e-mail: dippenaara@arc.agric.za
Received: 26 June 2008
Accepted: 31 Aug. 2008
Published: 23 Jan. 2009
DOI: 10.4102/koedoe. v51i1.161
This article is available at: http://www.koedoe.co.za


{kind=link}
{kind=link}