










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Surgery
On-line version ISSN 2078-5151
Print version ISSN 0038-2361
S. Afr. j. surg. vol.53 n.3-4 Cape Town Dec. 2015
OPINION
Concepts in malignant transformation
B D Bebington
Wits Donald Gordon Medical Centre, Parktown, Johannesburg, South Africa
The precise biochemical nature of malignant transformation remains elusive. This article reviews some of the current theories of oncogenesis and aims to synthesize these concepts into a possible schema for the origins of malignant transformation, using a cell-programmed origin for neoplastic progression. It is proposed that cell transformation in malignancy is initiated by injury but finds ultimate promulgation through cellular, self-determined, epigenetic events. Explanations for this arrangement are sought in evolutionary models and our knowledge of cell biology. The proposed mechanisms have not been experimentally validated, but will hopefully stimulate further research.
Concepts of carcinogenesis
When considering any existing theory of carcinogenesis four hypotheses have been developed to explain its nature:
Firstly, the incidence of cancer in any population increases with age1 and this increase is probably exponential, suggesting an accumulative risk of malignant conversion. Incidence is also dependent on the dose of exposure to a carcinogen.2,3
Secondly, it seems that it is at the level of genetic expression that this cumulative injury is occurring, whether this is an injury to the DNA itself4 or to the phenotypic expression of the genome.5
Thirdly, the cells within the tumour have to evolve morphologically and functionally within the environment of a multi-cellular milieu. A Darwinian paradigm has been proposed to explain clonal selection of metastases.5
Finally, malignancy develops out of growing tissue and is itself an aberrant tissue form. It therefore spreads and grows within the framework of a metazoan cellular environment. The normal processes of embryogenesis and growth have to be the context in which proliferation and metastasis arises. This has given rise to the concept of morphogens and morphostats that govern growth. When aberrantly expressed, these might result in the abnormal cell proliferation of malignant transformation. Morphogens and morphostats are known to govern the transformation of cells to premalignant states such adenoma formation and metaplasia.6
However, a closer look at development and embryogenesis suggests that the final effectors of micro-environment and morphostat/morphostat production are the cells themselves.
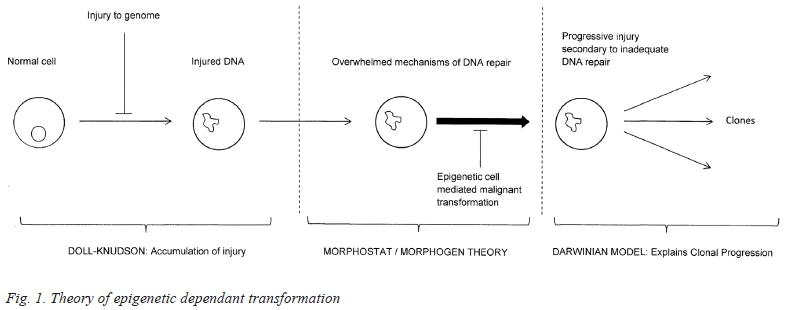
The orchestrated behaviour of cells in community is ultimately governed by the genome and its expression, implying that the initiation of malignant transformation is inherent within the cell. This coincides with another very obvious feature of malignant transformation, the predictability of the clinical progression of most cancers. The nature of the injury to the cell may be random but the response is specific. This response includes deregulation of growth and apoptosis, angiogenesis and metastasis. For any given malignancy these behavioural activities occur in a predictable fashion, a phenomenon familiar to all. Besides this, the orchestration is not only local but occurs wherever the cells metastasize, suggesting that this regulation is inherent within the cell and only secondarily dependent on the external environment. It is the eukaryote stem cell that initiates the event as a consequence of DNA injury when this has surpassed the capacity of internal repair mechanisms. Figure 1 combines the various theories into a consistent model.
The implication of these observations is that the eukaryote cell has retained an epigenetic capability that results in the demise of the organism. Why would an injurious event be programmed into the function of our cells? We know that the process of aging is the consequence of programmed cellular events and that this too results in the demise of the metazoan organism. Death of the individual is essential for the survival of the population. This may justify a biochemically mechanistic and cell-determined origin for events like aging and malignant transformation.
The chemoton model for the beginning of life proposes three chemical events: compartmentalisation by lipid layers or surface absorption, evolution of a metabolic pathway that helped convert energy into structure, and a chemical blue print enabling perpetual replication of systems.7 Theories of life's origin centre on the nature of these three biochemical domains as well as the ways in which they became associated. The most plausible basis of early biochemical reproduction is self-replicating RNA within lipid biospheres. RNA's stability, catalytic capabilities and ability to self-replicate make it the most likely starting point of biochemical evolution. In the postulated beginnings of life the redox difference between earth's crust and its atmosphere was a possible energy source that drove the whole early chemical process. Regardless of the details of the chemistry it is certain that any perpetual chemical process requires the provision of substrate perpetually.
In a closed system, life would only have been possible once the products of life's chemistry were converted back to substrate by a counter-reaction that produced the initial building blocks of life. This was probably not the same reaction in reverse but rather a completely different reaction dependent on different environmental variables, with an energy source independent of the "forward" reactions of life. Evolving biochemical expression had to have incorporated a message of timed termination so that once replication was complete the biochemical product was ready for conversion back into substrate. This message had to be biochemically mechanistic and not simply a passive process. In multicellular life forms malignancy may be one of these biochemical mechanisms that terminates the organism's life and provides for substrate in perpetuity. Thus malignancy has become an evolutionary advantage, even though it may be disadvantageous to the individual.
This replicative process is obviously present in all life forms. The bare RNA cycle of replicative growth followed by degeneration into substrate has probably grown in complexity with the accumulative assimilation of other biochemical groups (DNA, protein, fat, etc.). The cyclical replenishing of substrate may be why death is inevitable. The necessity for death may explain why malignant transformation has been preserved throughout evolution. But what then is the cellular event which initiates cancer? It is here proposed that its' beginnings can be traced to the origin of the eukarya. Regardless of the exact phylogeny of early life form, diversity has arisen out of evolutionary pressure and gives rise to the archaea and eubacteria.8 With a common origin, endosymbiotic fusion9 has been possible because the common language of replication and control has been shared in their RNA.10 The RNA dependent regulation of the fused prokaryotes was responsible not only for the amalgamation of function and form but also gave the new eukaryote the ability to express more than one phenotype from a wider choice of genetic information. When this regulation, as part of the cell's epigenetic expression, allows for demonstration of its' symbiotic origin, then, malignancy is made manifest.
Evolution of the eukaryote and its part in epigenetic expression of malignancy
There is no direct evidence that malignant transformation is the consequence of altered epigenetic expression. But are there any circumstantial observations to support this? The Warberg effect is the malignant cell's propensity to utilize anaerobic glycolysis over mitochondrion-dependent metabolism. It may be that in this phenomenon such indirect evidence exists. The mitochondrion is thought to have had its origin in the fusion of an anaerobic archebaterium and an aerobic eubacterium in the process of endosymbiosis. If this is the origin of all eukaryotes then mitochondrial DNA studies suggest this was a unique event and all eukaryotes have a single common ancestor, called the last eukaryote common ancestor (LECA). The event has even been given a date with reasonable confidence (2.5 billion years ago).11 At that time the earth was very different. Thermodynamic activity was more ubiquitous and earth's oxidative atmosphere was still new.12 It is thought that the Archaea, the prokaryote root of present day extremophiles, exploited the anaerobic energy of the reductive earth's core.13 The eubacteria on the other hand utilized the oxidative environment to bank on the huge energy advantages of oxidative phosphorylation. In the fusion of these two endosymbionts both reductive and oxidative environments were exploited, giving the combination an evolutionary advantage.14
In this fusion, major rearrangement in the genetic machinery would have had to occur. The eubacteria's genetic code was moved to the archebacterium's proto-nucleus. Hypothetically this was made possible by the noncoding genome characteristic of eukaryote genetic composition. Evolution of metazoan life forms has seen an expansion of exactly this part of the cell's DNA content.15 It is possible that the adaptive phenotypic expression in epigenetic variability seen in evolution, embryogenesis and growth has its origin in the genetic assimilation of endosymbiosis. Epigenetic expression may have had its origins in the addition of non-coding RNA made necessary in the process of endosymbiosis.
Cancer is a disturbance of the organisation of cells in community. Organisation of the community is dependent on epigenetic expression. The theoretical final cell-mediated step transforming a normal stem cell into a malignant cell might be an epigenetic transition. It is proposed that the epigenetic event that causes malignant transformation has its origins in the dawn of epigenetics and that the aberrant cell is expressing behaviour akin to the archebacterial half of the symbiont. The LECA may have been able to move from an oxidative environment in response to that circumstance's injurious effect on DNA. It would have used its newly acquired ability to vary its phenotype, move to a safer place, limit apoptosis-like death and replicate in reductive niche without aerobic metabolism. In other words, cancer may be a de-suppression of an ancient epigenetic instruction.
If we look at contemporary primitive organisms such as the protists, there are many that demonstrate behaviour that mirror these changes, especially the apicomplexa. It has fairly recently been discovered that plasmodia are partial if not complete facultative anaerobes with the end product of their anaerobic metabolism being glycerol. Transitions in the life cycles of these parasites are associated with migration, proliferation, reduction of apoptosis-like cell death and a change to anoxic metabolism. This offers no proof for the epigenetic capabilities of the LECA but may suggest that this putative organism had an adaptive advantage in the ability to behave as the plasmodia do. It is this behaviour retained as an epigenetic instruction in our cells that comes to the fore in the setting of cell injury and finds its expression in malignant transformation.
Competing Interests
None
Acknowledgements
I would like to thank Prof G J Oettle for reviewing various drafts of the paper. Dr Julie Herold helped with the format of the paper as well as the preparation of the illustrations.
REFERENCES
1. Armitage P, Doll R. The age distribution of cancer and a multistage theory of carcinogenesis. Br J Cancer 1954; 8:1-12. [http://dx.doi.org/10.1038/bjc.1954] [PMCID: PMC2007940] [ Links ]
2. Doll R, Peto R. Cigarette smoking and bronchial carcinoma: dose and time relationships among regular smokers and lifelong non-smokers. Journal of Epidemiology & Community Health. 1978 Dec 1;32(4):30313. [http://dx.doi.org/10.1136/jech.32.4.303] [PMID: 8422957] [ Links ]
3. Peto R. Smoking, smoking cessation, and lung cancer in the UK since 1950: combination of national statistics with two case-control studies. BMJ. 2000 Aug 5;321(7257):3239. [http://dx.doi.org/10.1136/bmj.321.7257.323] [PMID: 10926586] [PMCID: PMC27446] [ Links ]
4. Knudson A. Retinoblastoma: Teacher of Cancer Biology and Medicine. PLoS Medicine. 2005;2(10):e349. [http://dx.doi.org/10.1371/journal.pmed.0020349] [PMID: 16231982] [PMCID: PMC1261516] [ Links ]
5. Greaves M. Darwinian medicine: a case for cancer. Nature Reviews Cancer. 2007 Feb 15;7(3):21321. [http://dx.doi.org/10.1038/nrc2071] [PMID: 17301845] [ Links ]
6. Van den Brink GR, Offerhaus GJ. The Morphogenetic Code and Colon Cancer Development. Cancer Cell. 2007 Feb;11(2):10917. [http://dx.doi.org/10.1016/j.ccr.2007.01.003] [PMID: 17292823] [ Links ]
7. Szathmáry E, Demeter L. Group selection of early replicators and the origin of life. Journal of Theoretical Biology. 1987 Oct;128(4):46386. [http://dx.doi.org/10.1016/S0022-5193(87)80191-1][PMID: 17292823] [ Links ]
8. Woese CR, Fox GE. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proceedings of the National Academy of Sciences [Internet]. [ Links ] 1977 Nov 1;74(11):508890. [http://dx.doi.org/10.1073/pnas.74.11.5088][PMID: 270744]
9. Schwartz W, Lynn Margulis. Origin of Eukaryotic Cells. Evidence and Research Implications for a Theory of the Origin and Evolution of Microbial, Plant, and Animal Cells on the Precambrian Earth. XXII u. 349 S., 89 Abb., 49 Tab. New Haven-London 1970: Yale University Press. Zeitschrift für allgemeine Mikrobiologie. 1973;13(2):186186. [http://dx.doi.org/10.1002/jobm.3630130220] [ Links ]
10. Olsen GJ, Woese CR. Ribosomal RNA: a key to phylogeny. FASEB J 1993; 7:113-123 [PMID: 8422957] [ Links ]
11. Gu X. The age of the common ancestor of eukaryotes and prokaryotes: statistical inferences. Molecular Biology and Evolution. 1997 Aug 1;14(8):8616. [http://dx.doi.org/10.1093/oxfordjournals.molbev.a025827] [PMID: 9254924] [ Links ]
12. Wachtershauser G. From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya. Philosophical Transactions of the Royal Society B: Biological Sciences. 2006 Oct 29;361(1474):1787808. [http://dx.doi.org/10.1098/rstb.2006.1904] [PMID: 17008219] [PMCID: PMC1664677] [ Links ]
13. Thrash JC, Boyd A, Huggett MJ, Grote J, Carini P, Yoder RJ, et al. Phylogenomic evidence for a common ancestor of mitochondria and the SAR11 clade. Scientific Reports. 2011 Jun 14;1. [http://dx.doi.org/10.1038/srep00013] [PMID: 22355532] [PMCID: PMC3216501] [ Links ]
14. Kleine T, Maier UG, Leister D. DNA Transfer from Organelles to the Nucleus: The Idiosyncratic Genetics of Endosymbiosis. Annual Review of Plant Biology. 2009 Jun;60(1):11538. [http://dx.doi.org/10.1146/annurev.arplant.043008.092119] [PMID: 19014347] [ Links ]
15. Zmasek CM, Godzik A. Strong functional patterns in the evolution of eukaryotic genomes revealed by the reconstruction of ancestral protein domain repertoires. Genome Biology. 2011;12(1):R4. [http://dx.doi.org/10.1186/gb-2011-12-1-r4] [PMID: 19014347] [ Links ]
 Correspondence:
Correspondence:
Brendan Bebington
brendanbebington@telkomsa.net


{kind=link}