










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.120 n.3-4 Pretoria Mar./Apr. 2024
http://dx.doi.org/10.17159/sajs.2024/16387
RESEARCH ARTICLE
Jesse M. MartinI; A.B. LeeceI, II; Andy I.R. HerriesI, III; Stephanie E. BakerIII; David S. StraitIII, IV
IPalaeoscience Labs, Department of Archaeology and History, La Trobe University, Melbourne, Victoria, Australia
IIGeoarchaeology and Archaeometry Research Group, Faculty of Science and Engineering, Southern Cross University, Lismore, New South Wales, Australia
IIIPalaeo-Research Institute, University of Johannesburg, Johannesburg, South Africa
IVDepartment of Anthropology, Washington University in St Louis, St. Louis, Missouri, USA
ABSTRACT
Classic depictions of human evolutionary ecology cast Homo as predator and other hominins, including Paranthropus robustus, as prey. Such hypotheses rest on a small number of fossils that exhibit evidence of carnivore predation, including the iconic SK 54 cranium from Swartkrans in South Africa. Here we demonstrate that the SK 54 cranium shares its closest affinities with H. erectus sensu lato rather than P. robustus. Demonstrating that Homo was prey for leopards at Swartkrans weakens the historically significant hypothesis that Homo was better able to avoid predation because of being behaviourally and technologically advanced compared to Paranthropus. Subsequent ideas about hominin palaeobiology derived from this hypothesis warrant reconsideration.
SIGNIFICANCE:
• The small sample of early Homo from southern Africa is increased by the allocation of the SK 54 cranium to that genus.
• Evidence from Swartkrans suggests that Homo was prey for leopards.
• Hypotheses concerning the biology, behaviour, and technological capabilities of Homo and P. robustus stemming from Brain's seminal work, The Hunters or the Hunted?, should be reassessed.
Keywords: Homo erectus, Swartkrans, Paranthropus robustus, taphonomy, early Homo
Introduction
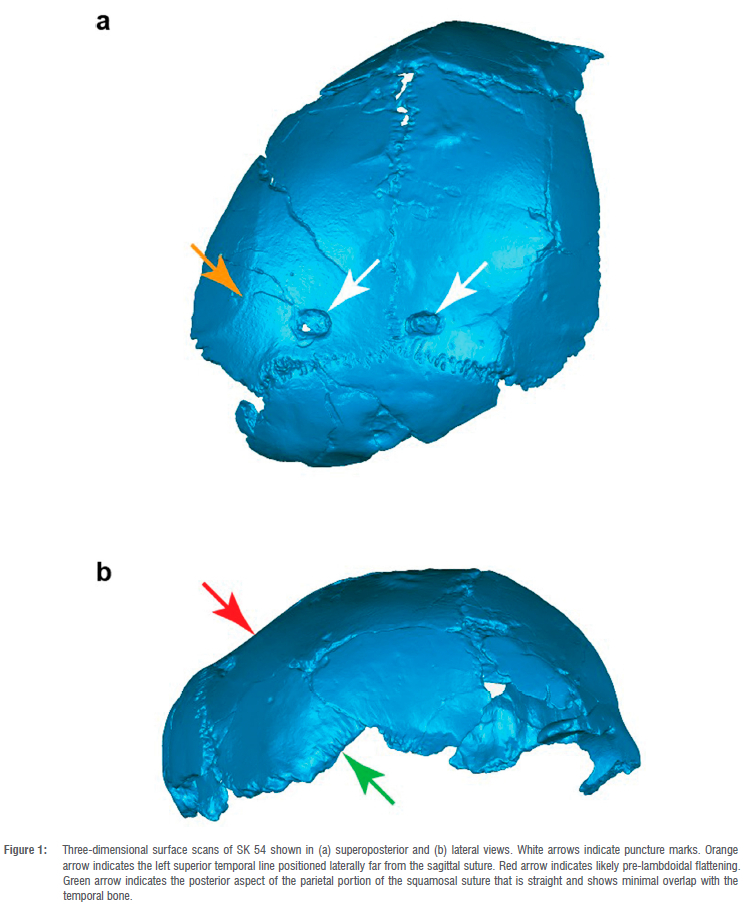
SK 54 is a partial hominin cranium that was recovered from the -1.9-1.8 Ma palaeocave deposits of Swartkrans Member 1 Hanging Remnant in South Africa in 1949 and subsequently prepared by John T. Robinson.13 This specimen is an iconic hominin fossil that has influenced both the development of the discipline of cave taphonomy and narratives concerning how multiple hominin species shared the landscape of Pleistocene southern Africa.1 Brain1,2 described two carnivore puncture marks on its left and right parietal bones (Figure 1) whose location, size, and spacing indicate strongly that they were inflicted by a leopard (famously, the marks conform well to the canines of leopard fossil SK 349 from the same deposit).4 Leopards are known to be predators rather than scavengers1,5, and thus this specimen preserves direct evidence of the predation of hominins. Even if future taphonomic analyses conclude that these puncture marks were not caused by a predator, the historical significance of Brain's4 assessment remains. For this reason, although there are other hominin fossils from Swartkrans that exhibit carnivore modification marks1,2,6,7, no other specimen has figured as centrally in hypotheses concerning carnivore predation on hominins than SK 541,2,6,8-17. The specimen has previously been attributed to Paranthropus robustus1, and that taxonomy has remained largely unchallenged and current18-21. Brain1 did not explain the reasoning underlying this taxonomic decision, although he did note that the specimen appeared to possess a relatively small brain. Tobias22 and Clarke23 noted that SK 54 lacked certain derived circumorbital morphology characteristic of P. robustus, but did not assign the specimen to a different species and interpreted the variation as a product of ontogeny. We do, however, note that Braga et al.24 have recently questioned SK 54's attribution to P. robustus based on differences between its circumorbital and postorbital morphology and that of the juvenile P. robustus specimen KW 9000/9600 from Kromdraai, and we largely agree with their conclusions. Notwithstanding this recent reassessment, SK 54's attribution to P. robustus has underwritten hypotheses that australopiths were prey while early Homo were transforming into predators, as elucidated in Brain's2 classic monograph, The Hunters or The Hunted? Here we provide further taxonomic evidence that challenges this narrative.
Materials and methods
Our analysis of SK 54 results principally from morphological observations conducted in South Africa in 2018, 2019 and 2022 on original fossil specimens of Australopithecus africanus (Taung 1; Sts 5, Sts 71, Stw 505), A. sediba (MH 1), P. robustus (DNH 7, DNH 155, DNH 152, SK 46, SK 48 and SK 52) and early Homo (Stw 53, SK 847) curated at the Evolutionary Studies Institute of the University of the Witwatersrand, and the Ditsong Museum of Natural History. Because SK 54 is a subadult, we closely compared SK 54 to available subadult specimens of H. erectus sensu lato (DNH 134) and a description of P. robustus (KW 9000/9600) from southern Africa. We additionally utilised published descriptions and/or casts of subadult specimens attributed to H. erectus (Mojokerto 1 and KNM-ER 42700) and P. aethiopicus (L338y-6).
Results
SK 54 is a fragmentary neurocranium preserving parts of the occipital, frontal, and left and right parietal bones. The degree of sutural fusion suggests that the specimen may have been a juvenile at the time of death. The specimen is plasticly deformed such that it is not possible to assess overall neurocranial shape, and this deformation precludes meaningful quantitative analysis. One measurement that can be confidently taken on the specimen is cranial vault thickness, which we assess as being minimally 3.3 mm at the posterior aspect of the parietal bone just above the squamosal suture. This measurement compares favourably to that of the subadult H. erectus specimen DNH 134 (3.2 mm) and is thinner than that of the subadult P. robustus specimen KW 9000 / 9600 at 4.0 mm.24 Although H. erectus specimens are often characterised as having thick vaults, this characteristic is more strongly expressed in Asian rather than early African specimens25 and in any case the taxonomic valence of this trait is compromised by SK 54's likely young ontogenetic age (see also Antón26). Notably, vault thickness in the vicinity of bregma is qualitatively thin in Modjokerto27, a juvenile specimen conventionally attributed to H. erectus26. SK 54's neurocranial vault is distorted on both the right and left sides, making a digital reconstruction highly subjective and of little diagnostic value. The individual evidently had a generally small brain, but estimates of its cranial capacity cannot be made with confidence.28 Nonetheless, aspects of preserved morphology challenge its traditional taxonomic attribution to Paranthropus and suggest affinities with Homo. The temporal lines are well separated and laterally positioned (Figure 1) - a configuration that is incompatible with adult P. robustus specimens that exhibit either a sagittal crest (in putative male specimens) or nearly convergent temporal lines (in female specimens).29 The KW 9000/9600 fossil from Kromdraai provides the only evidence for temporal line configuration in a subadult P. robustus, and the superior temporal line is clearly closer to midline in the vicinity of bregma24 than the weakly expressed and more laterally positioned temporal lines exhibited by SK 54. Similarly, the juvenile P. aethiopicus specimen L338y-6 (whose sutures are as or more open than those of SK 54, implying a coarse similarity in age) exhibits well-developed, strongly convergent temporal lines30, further suggesting that SK 54 may not be Paranthropus. Kimbel et al.20 have previously argued that SK 54 preserves rugose striae parietalis that they suggest are correlated with a high degree of overlap between the temporal squama and parietal at the squamosal suture. They based their inference on the observation by Rak31 that juvenile H. sapiens from a Holocene population exhibited fine rather than rugose striae. However, the length of the striae parietalis preserved in SK 54 (Figure 1) are notably less than those of L338y-6 and adult P. robustus specimens DNH7, DNH152, and DNH155, and both the length and rugosity of SK 54's striae closely resemble the condition in the juvenile H. erectus sensu lato specimen DNH 134. In our assessment, enough of the inferior bevelled edge of the right parietal (Figure 1) is preserved to indicate that the temporal and parietal portions of the squamosal suture would not have overlapped extensively, unlike the extensive overlap seen in both adult and juvenile Paranthropus.30-32 Finally, contra Kimbel et al.20, the very wide separation of the superior temporal lines on the frontal bone argues against the inference that a frontal trigon would have developed in adulthood.
Two discrete traits of SK 54 may suggest affinities with Homo erectus sensu lato. The preserved posterior portion of the squamosal suture is straight as it rises anteriorly and superiorly (Figure 1). Rightmire et al.33 note that, "A low temporal squama with a straight upper border passing downward toward asterion is a consistent marker for [H. erectus]." Moreover, SK 54's parietal bones are flattened anterior to the lambdoidal suture (Figure 1), although there is distortion present in this region. Pre-lambdoidal flattening is a derived characteristic of many H. erectus specimens; in their description of the skulls from Dmanisi, Rightmire et al.33 state that:
There are also good indications that the Dmanisi skulls share at least a few (specialized?) characters with populations from the Far East. An example is provided by parasagittal flattening of the posterior vault. Flattening or even depression of the parietal surfaces is pronounced at Dmanisi, and it is common in the [H.] erectus crania from Sangiran in Java.
The fragmentary and deformed nature of SK 54 precludes a definitive taxonomic allocation but, heuristically, superimposing SK 54 onto H. erectus sensu lato specimen KnM-ER 4270034 demonstrates a striking similarity between the two specimens (Figure 2). We agree with Braga et al.24 that a provisional assignment of SK 54 to Homo seems warranted, and we argue that a tentative species-level allocation to H. erectus sensu lato is plausible. This assignment adds to the evidence for Homo at Swartkrans Member 1 Hanging Remnant that includes the juvenile cranium SK 27 that Clarke35 reclassified from P. robustus to Homo, and the suggested partial skull consisting of individual fossils SK 80, SK 846b, SK 84736 and sometimes also the mandible fragment SK 452 . At 1.9-1.8 Ma3, these fossils are slightly younger than the 2.041.95 Ma DNH 134 cranium from Drimolen Main Quarry that also shows affinities to Homo erectus32.
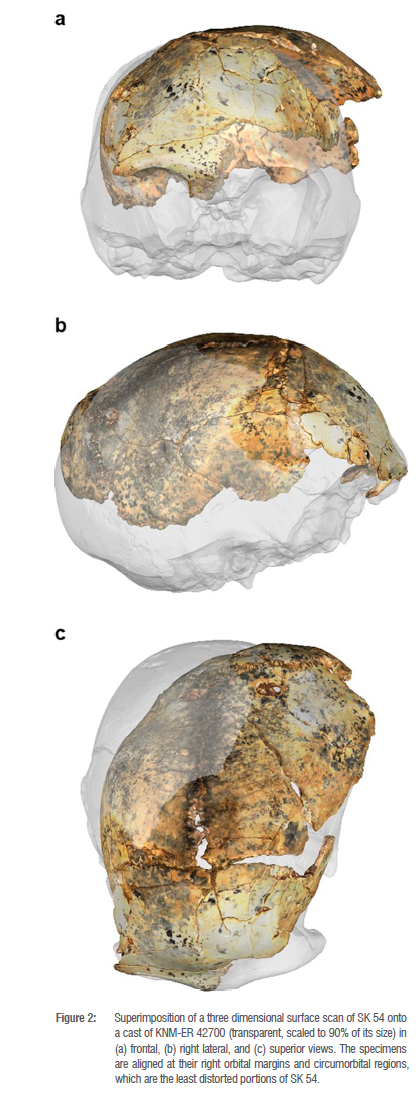
Discussion
Brain's work on cave taphonomy remains seminal, and his taphonomic assessment linking SK 54 with leopard predation is currently unchallenged. Assuming that those taphonomic conclusions remain valid, our taxonomic reassessment of SK 54 demonstrates that Homo was also prey for leopards in the early Pleistocene, and this characterisation could not be further removed from classic depictions of Homo the hunter and Paranthropus the hunted. Allocating SK 54 to Homo tempers the impetus for supposing that early Homo and P robustus were differentially predated because of the former's behavioural and technological advancement.
It is impossible to know with certainty how the history of palaeoanthropology might have been different had SK 54 been recognised as Homo when it was first discovered, but it is reasonable to infer that the impact of such a realisation would have been significant. Only 6 years prior to the publication of Brain's1 now-classic paper, the description of the newly discovered H. habilis36 included an assessment of the relative tool-making skills and trophic positions of the new species and its contemporary, Zinjanthropus boisei (i.e. P boisei):
While it is possible that Zinjanthropus and Homo habilis both made stone tools, it is probable that the latter was the more advanced tool maker and that the Zinjanthropus skull represents an intruder (or a victim) on a Homo habilis living site.37
Shortly thereafter, the highly influential Man the Hunter conference was held, followed by the publication of its accompanying edited volume38,39 that described hunting as a fundamentally important human adaptation. Brain's2,3 interpretation of SK 54, based on an incorrect taxonomy, was therefore compatible with the thinking of the time, but it could have instead been a powerful challenge to conventional wisdom. Ideas, like species, evolve and have descendants, so the evidence presented here should prompt a reassessment of hypotheses concerning the biology, behaviour, and technological capabilities of Homo and Paranthropus that are derived from earlier ideas positing Homo as predator and Paranthropus as prey (for example, Lockwood et al.29). Our findings complement a recent zooarchaeological analysis showing that the appearance of H. erectus is not associated with increased evidence for hominin carnivory.38
Acknowledgements
We thank the DITSONG Natural History Museum and the hominin fossil curator Mirriam Tawane for facilitating access to the fossil hominin assemblage. We also thank the University of the Witwatersrand Evolutionary Studies Institute and the hominin fossil curators Bernhard Zipfel and Sifelani Jirah for facilitating access to the hominin fossil collection. We thank Kieran McNulty for sharing comparative data and contributing to discussions concerning hominin diversity at Drimolen. This research was funded and supported by HDR fee waivers and living scholarships from La Trobe University to J.M.M. and an Australian Research Council Discovery Grant DP170100056 to A.I.R.H. and D.S.S.
Competing interests
We have no competing interests to declare.
Authors' contributions
J.M.M., A.B.L., A.I.R.H., S.E.B., and D.S.S. wrote the paper. J.M.M. and D.S.S. additionally participated in conceptualisation, data collection, sample analysis, and data analysis.
References
1. Brain CK. New finds at the Swartkrans australopithecine site. Nature. 1970;255:1112-1119. https://doi.org/10.1038/2251112a0 [ Links ]
2. Brain CK. The hunters or the hunted? Chicago, IL: Chicago University Press; 1981. [ Links ]
3. Herries AIR. Chronology of the hominin sites of southern Africa. In: Oxford Research Encyclopedia of Anthropology. Oxford: Oxford University Press; 2022. https://doi.org/10.1093/acrefore/9780190854584.013.57 [ Links ]
4. Brain CL. The probable role of leopards as predators of the Swartkrans australopithecines. S Afr Archaeol Soc. 1969;224:170-171. https://doi.org/10.2307/3888296 [ Links ]
5. Hayward MW, Henschel P O'Brien J, Hofmeyr M, Balme G, Kerley GIH. Prey preferences of the leopard (Panthera pardus). J Zool. 2006;270:298-313. https://doi.org/10.1111/j.1469-7998.2006.00139.x [ Links ]
6. White TD. The comparative biology of "robust" Australopithecus: Clues from context. In: Grine FE, editor. Evolutionary history of the "robust" australopithecines. Piscataway, NJ: Transaction Publishers; 1988. p. 449-484. [ Links ]
7. Pickering TR. Taphonomy of the Swartkrans hominid postcrania and its bearing on issues of meat-eating and fire management. In: Stanford CB, Bunn HT, editors. Meat-eating and human evolution. New York: Oxford University Press; 2001. p. 33-51. https://doi.org/10.1093/oso/9780195131390.003.0003 [ Links ]
8. Holloway RL. New australopithecine endocast, SK 1585, from Swartkrans, South Africa. Am J Phys Anthropol. 1972;37:173-186. https://doi.org/10.1002/ajpa.1330370203 [ Links ]
9. Blumenberg B, Todd NB. On the association between Homo and Australopithecus. Curr Anthropol. 1974;15(4):386-388. https://doi.org/10.1086/201493 [ Links ]
10. Susman RL. Pygmy chimpanzees and common chimpanzees: Models for the behavioral ecology of the earliest hominids. In: Kinzey WG, editor. Evolution of human behviour. Albany, NY: State University of New York Press; 1987. p. 72-86. [ Links ]
11. Lee-Thorp J, Thackeray JF, Van der Merwe N. The hunters and the hunted revisited. J Hum Evol. 2000;39:565-576. https://doi.org/10.1006/jhev.2000.0436 [ Links ]
12. Pickering TR, Dominquez-Rodrigo M, Egeland CP Brain CK. Beyond leopards: Tooth marks and the contribution of multiple carnivore taxa to the accumulation of the Swartkrans Member 3 fossil assemblage. J Hum Evol. 2004;46:595-604. https://doi.org/10.1016/j-jhevol.2004.03.002 [ Links ]
13. Pickering TR. Underrated prey? Evol Anthropol. 2005;14:159-164. https://doi.org/10.1002/evan.20070 [ Links ]
14. Gommery D, Pickford M, Senut B. A case of carnivore-inflicted damage to a fossil femur from Swartkrans, comparable to that on a hominid femur representing Orrorin tugenensis, BAR 1003'00 (Kenya): Scientific notes. Ann Transvaal Mus. 2007;44:215-218. https://hdl.handle.net/10520/EJC83650 [ Links ]
15. Pobiner BL. Paleoecological information in predator tooth marks. J Taphonomy. 2008;6:373-397. [ Links ]
16. Val A, Taru P, Steininger C. New taphonomic analysis of large-bodid primate assemblages from Cooper's D, Bloubank Valley, South Africa. S Afr Archaeol Bull. 2014;69:49-58. [ Links ]
17. Arriaza MC, Aramendi J, Mate-Gonzalez MA, Yravedra J, Stratford D. The hunted or the scavenged? Australopith accumulation by brown hyenas at Sterkfontein (South Africa). Quat Sci Rev. 2021;273, Art. #107252. https://doi.org/10.1016/j.quascirev.2021.107252 [ Links ]
18. Wolpoff MH. Is the new composite cranium from swartkrans a small robust australopithecine? Nature. 1971;230:398-401. https://doi.org/10.1038/230398a0 [ Links ]
19. Wolpoff MH. Sagittal cresting in the South African australopithecines. Am J Phys Anthropol. 1974;40:397-408. https://doi.org/10.1002/ajpa.1330400312 [ Links ]
20. Kimbel WH, White TD, Johanson DC. Cranial morphology of Australopithecus afarensis: A comparative study based on a composite reconstruction of the adult skull. Am J Phys Anthropol. 1984;64:337-388. https://doi.org/10.1002/ajpa.1330640403 [ Links ]
21. Rak Y Kimbel WH, Moggi-Cecchi J, Lockwood CA, Menter C. The DNH 7 Skull of Australopithecus robustus from Drimolen (Main Quarry), South Africa. J Hum Evol. 2021;151:102913. https://doi.org/10.1016/j.jhevol.2020.102913 [ Links ]
22. Tobias PV. Olduvai Gorge, Vol 2. The cranium and maxillary dentition of Australopithecus (Zinjanthropus) boisei. Cambridge: Cambridge University Press; 1967. [ Links ]
23. Clarke RJ. The cranium of the Swartkrans hominid SK 847 and its relevance to human origins. Johannesburg: University of the Witwatersrand; 1977. [ Links ]
24. Braga J, Wood BA, Zimmer VA, Moreno B, Miller C, Thackeray JF, et al. Hominin fossils from Kromdraai and Drimolen inform Paranthropus robustus craniofacial ontogeny. Sci Adv. 2023;9, eade7165. https://doi.org/10.1126/sciadv.ade7165 [ Links ]
25. Antón S. Natural history of Homo erectus. Am J Phys Anthropol. 2003; 122:126-170. https://doi.org/10.1002/ajpa.10399 [ Links ]
26. Antón S. Developmental age and taxonomic affinity of the Mojokerto child, Java, Indonesia. Am J Phys Anthropol. 1997;102:497-514. https://doi.org/10.1002/(SICI)1096-8644(199704)102:4<497::AID-AJPA6>3.0.CO;2-P [ Links ]
27. Coqueugniot H, Hiblin J-J, Veillon F, Houet F, Jacob T. Early brain growth in Homo erectus and implications for cognitive ability. Nature. 2004;431:299-302. https://doi.org/10.1038/nature02852 [ Links ]
28. Holloway RL, Broadfield DC, Yuan MS. The human fossil record: Brain endocasts - The paleoneurological evidence. Vol. 3. New York: Wiley-Liss; 2004. https://doi.org/10.1002/0471663573 [ Links ]
29. Lockwood CA, Menter CG, Moggi-Cecchi J, Keyser AW. Extended male growth in a fossil hominin species. Science. 2007;318:1443-1446. https://doi.org/10.1126/science.1149211 [ Links ]
30. Rak Y Howell CF. Cranium of a juvenile Australopithecus boisei from the Lower Omo Basin, Ethiopia. Am J Phys Anthropol. 1978;48:345-365. https://doi.org/10.1002/ajpa.1330480311 [ Links ]
31. Rak Y The functional significance of the squamosal suture in Australopithecus boisei. Am J Phys Anthropol. 1978;49:71-78. https://doi.org/10.1002/ajpa.1330490111 [ Links ]
32. Herries AIR, Martin JM, Leece AB, Adams JW, Boschian G, Joannes-Boyau R, et al. Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa. Science. 2020;368:47. https://doi.org/10.1126/science.aaw7293 [ Links ]
33. Rightmire PG, Lordkipanidze D, Vekua A. Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia. J Hum Evol. 2006;50:115-141. https://doi.org/10.1016/j.jhevol.2005.07.009 [ Links ]
34. Spoor F, Leakey MG, Gathogo PN, Brown FH, Anton SC, McDougall I, et al. Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature. 2007;448:688-691. https://doi.org/10.1038/nature05986 [ Links ]
35. Clarke RJ. A juvenile cranium and some adult teeth of early Homo from Swartkrans, Transvaal. S Afr J Sci. 1977;73:46-19. [ Links ]
36. Clarke RJ, Howell FC, Brain CK. More evidence of an advanced hominid at Swartkrans. Nature. 1970;225:1219-1222. https://doi.org/10.1038/2251219a0 [ Links ]
37. Leakey LSB, Tobias PV, Napier JR. A new species of the genus Homo from Olduvai Gorge. Nature. 1964;202:7-9. https://doi.org/10.1038/202007a0 [ Links ]
38. Lee RB, DeVore I, editors. Man the hunter. The first intensive survey of a single, crucial stage of human development - man's once universal hunting way of life. Chicago, IL: Aldine; 1968. [ Links ]
39. Barr WA, Pobiner B, Rowan J, Faith JT. No sustained increased in zooarchaeological evidence for carnivory after the appearance of Homo erectus. Proc Natl Acad Sci USA. 2022;119, e2115540119. https://doi.org/10.1073/pnas.2115540119 [ Links ]
 Correspondence:
Correspondence:
Jesse Martin
Email:jesse.martin@latrobe.edu.au
Received: 26 Jun. 2023
Revised: 20 Nov. 2023
Accepted: 08 Dec. 2023
Published: 27 Mar. 2024
Editor: Jemma Finch
Funding: La Trobe University, Australian Research Council (Discovery Grant DP170100056)


{kind=link}