










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.120 no.3-4 Pretoria mar./abr. 2024
http://dx.doi.org/10.17159/sajs.2024/14917
RESEARCH ARTICLE
Sanguinarine highly sensitises breast cancer cells to doxorubicin-induced apoptosis
Manisha du PlessisI; Carla FourieI; Heloise le RouxI; Anna-Mart EngelbrechtI, II
IDepartment of Physiological Sciences, Faculty of Science, Stellenbosch University, Stellenbosch, South Africa
IIAfrican Cancer Institute (ACI), Department of Global Health, Faculty of Medicine and Health Sciences, Stellenbosch University, Cape Town, South Africa
ABSTRACT
Breast cancer is the most commonly diagnosed cancer and the second most common cause of cancer death in women. The anthracycline, doxorubicin, is a well-known and highly effective treatment for breast cancer patients; however, many patients present with resistance to chemotherapeutic drugs, which ultimately results in treatment failure and contributes to high mortality rates. It is well established that the mitogen-activated protein kinase phosphatase 1 (MKP-1) mediates the response to chemotherapy, where upregulated MKP-1 is associated with chemoresistance. We investigated whether MKP-1 inhibition or silencing can sensitise triple-negative MDA-MB-231 breast cancer cells to doxorubicin therapy. We found that MKP-1 inhibition and silencing sensitises breast cancer cells to doxorubicin-induced apoptosis. Additionally, the inhibition of MKP-1 in combination with doxorubicin treatment promotes autophagy induction, while doxorubicin and not MKP-1 modulation increased lysosomal acidic compartments. As such, this study demonstrated that MKP-1 inhibition has a potential therapeutic benefit for breast cancer patients by increasing the efficacy of conventional chemotherapy. Therefore, MKP-1 inhibition should be developed as a clinically relevant adjuvant therapy, which could provide a novel avenue for therapeutic intervention in combination with chemotherapy in breast cancer patients.
SIGNIFICANCE:
• MKP-1 inhibition with sanguinarine and silencing sensitises breast cancer cells to doxorubicin-induced apoptosis.
• The inhibition of MKP-1 with sanguinarine in combination with doxorubicin treatment promotes autophagy induction.
• MKP-1 inhibition can have a potential therapeutic benefit for breast cancer patients by increasing the efficacy of conventional chemotherapy.
Keywords: breast cancer, MKP-1, doxorubicin, MAPKs, autophagy, apoptosis
Introduction
Currently, 90% of chemotherapy failures are related to drug resistance.1 Resistance to cytotoxic chemotherapeutic agents may often be random and unpredictable and becomes apparent only at clinical assessment.2 One such chemotherapeutic agent is the anthracycline, doxorubicin (Dox), which is often used as a first-line therapy for the treatment of breast cancer.3-5 However, its clinical effects are limited due to its dose-dependent side effects such as cardiotoxicity, gastrointestinal problems, acute vomiting, liver damage and nephropathy.6,7 Therefore, adjuvant therapies are required that can improve the effectiveness of Dox while conferring a protective effect on normal cells.
The mitogen-activated protein kinase phosphatase 1 (MKP-1) is a member of the MKP family that consists of 11 dual-specificity phosphatases that negatively regulate MAPk. Upon activation, MKP-1 dephosphorylates and deactivates MAPKs, ERK, JNK, and p38, by dephosphorylating specific Thr/Tyr residues.8 As such, MKP-1 plays an important role in tumorigenesis9 and negates the cytotoxicity of anticancer drugs in various models of cancer10-16. It has been reported that the knockdown of MKP-1 sensitises cancer cells through the upregulation of MAPK activity.14 The MAPK family mediates cell death through the mitochondrial pathway during conditions of cellular stress, for example, treatment with chemotherapeutic drugs, which is characterised by increased reactive oxygen species (ROS) production in the case of doxorubicin. Here, elevated ROS levels primarily activate the JNK and p38 pathways, resulting in mitochondrial-mediated apoptosis. Furthermore, MKP-1 also regulates the cell cycle and apoptosis through the modulation of mitochondrial function, oxidative stress and autophagy.17,18 As such, targeting MKP-1 has the potential to effectively sensitise breast cancer cells to Dox treatment.
Autophagy is a cellular degradation process characterised by the sequestration of cytoplasm, long-lived proteins and cellular organelles in double-membrane vesicles called autophagosomes, which are delivered to and subsequently degraded in lysosomes. However, its role in chemoresistance is complex, and often cell type and context dependent.19-21
Sanguinarine is a bioactive quaternary benzophenanthridine alkaloid plant extract from Sanguinaria canadensis (blood root), Poppy fumaria, Bocconia frutescens and Macleaya cordata and is a structural homologue of chelerythrine.22-26 Sanguinarine extracts have long been used in herbal medicine and have antimicrobial, antioxidant, anti-inflammatory and pro-apoptotic activity, and Sanguinarine can downregulate MKP-1 expression.23,26-29 Sanguinarine has been reported to decrease anti-apoptotic proteins, such as Bcl-2, and increase levels of pro-apoptotic Bax in pancreatic carcinoma cells, while decreasing the expression of tumour-promoting NFkP in cervical cancer cells, indicating a promising possibility for breast cancer treatment.28,29 Therefore, the aim of this study was to investigate whether the modulation of MKP-1 improves the potential of Dox to induce apoptosis and autophagy in cancer cells in vitro.
Materials and methods
Cell culture
The triple-negative human mammary adenocarcinoma cell line, MDA-MB-231, obtained from American Type Culture Collection (Rockville, MD, USA), was used in this study. The MDA-MB-231 cells were cultured in Glutamax-Dulbecco's Modified Eagle's Medium (DMEM; Celtic Molecular Diagnostics, Cape Town, South Africa) supplemented with 10% foetal bovine serum (FBS) (Sigma Chemical Co., St Louis, MO, USA) and penicillin streptomycin (PenStrep; Sigma Chemical Co., St Louis, MO, USA). The cells were maintained in a humidified incubator set at 5% CO2 at 37 °C and were routinely sub-cultured upon reaching 70-80% confluency.
Experimental protocol
Doxorubicin (D1515, Sigma Chemical Co., St Louis, MO, USA) was diluted to 5 in Glutamax-DMEM in ready to use aliquots; sanguinarine (Sang; Sigma Chemical Co., St Louis, MO, USA) was first diluted in methanol to a concentration of 0.01 M followed by dilution to 0.5 μM in growth medium; and dexamethasone (Dex; Sigma Chemical Co., St Louis, MO, USA) was dissolved to 0.1 M in distilled water and subsequently in growth medium to a final concentration of 100 nM; methanol was used as the vehicle. The concentration of Dox was determined with dose- and time-responses, which is presented in Supplementary figure 1. The following experimental groups were used in this study: (1) control, (2) Dex, (3) Dex and Dox, (4) Dox, (5) Sang, (6) Sang and Dox, (7) MKP-1 siRNA, (8) MKP-1 siRNA and Dox, (9) negative control (stealth RNAi) and (10) vehicle. The Sang groups were treated for 24 h, after which the Dox and Dex groups, including the combination treatment groups, were added and left to incubate for an additional 24 h.
MKP-1 siRNA Transfection
Silencing of MKP-1 gene expression was performed using reverse transfection. A volume of 20 pmol of MKP-1 siRNA duplex was used per culture well. And 20 pmol of MKP-1 siRNA duplexes (DUSP1VHS40581 and DUSP1VHS40583; Invitrogen™, USA, MKP-1 siRNA #159527B10, #159527B11) was diluted into transfection medium (Glutamax-DMEM containing no antibiotics or serum), after which Lipofectamine™ RNAiMAX (13778075; Invitrogen™, USA) was added and gently mixed. The RNAi duplex-Lipofectamine RNAiMAX complexes were allowed to incubate for 40 min and then made up to a final volume using DMEM (no antibiotics). MDA-MB-231 cells were plated directly into culture plates containing the RNAi duplex-Lipofectamine RNAiMAX complexes. Stealth™ RNAi (STEALTH RNAI NEG CTL MED GC, 12935300; Invitrogen™, USA) was used as a negative control. The cells were left to incubate for 48 h, after which Dox was added for a duration of 24 h.
Cell viability with trypan blue
The trypan blue exclusion technique30 was used to determine the viability of MDA-MB-231 cells. Following completion of the treatment protocol, the cell monolayer was rinsed with warm sterile phosphate-buffered saline (PBS). Cells were trypsinised and centrifuged for 3 min at 1500 rpm. The supernatant was decanted, and the pellets were resuspended in sterile PBS. The cell suspension was mixed in a ratio of 1:1 with 0.4% trypan blue and loaded into a haemocytometer for counting. The total number of cells was counted, and the number of blue cells was used as an indication of cell death. The results were expressed as the percentage (%) of viable cells.
Caspase 3/7 Glo assay
The Caspase-3/7 activity was measured in MDA-MB-231 cells using the Caspase-Glo® 3/7 assay (Promega, Madison, WI, USA). The Caspase-Glo® 3/7 reagent was prepared and equilibrated at room temperature and mixed with the lyophilised substrate before the reconstituted working buffer reagent was stored at -20 °C. The working buffer reagent and the culture plate were equilibrated to room temperature prior to use. Following the addition of the working buffer solution, the culture plate was incubated at 22 °C for 1 h without exposure to light. The content of each well was transferred to a white-walled 96-well plate, and the luminescence was measured using a luminometer.
Morphological analysis with Hoechst33342 and LysoTracker
For morphological determination of cell death, MDA-MB-231 cells were stained with Hoechst 33342 (10 mg/mL). Hoechst 33342 was made up in the growth medium and was left to incubate for 5 min. Thereafter, LysoTracker™ Red (Invitrogen™, USA) was added to each well and was left to incubate for an additional 5 min. Images were acquired directly after the incubation period through an Olympus Cell® system attached to an IX-81 inverted fluorescence microscope equipped with an F-view-II cooled CCD camera (Soft Imaging Systems). By using a Xenon-Arc burner (Olympus Biosystems GMBH) as a light source, images were excited with the 360 nm DAPI and 572 nm excitation filter. Emission was collected using a UBG triple band pass emission filter cube. Images were processed and background subtracted using the Cell® software. A minimum of three images were taken in randomly chosen fields, where at least three independent experiments per treatment group were conducted. Morphological changes such as condensed nuclear chromatin and apoptotic bodies were quantified using the Cell® system.
Flow cytometry
LysoTracker™ Red (Invitrogen™, USA) was used to analyse the lysosomal acidic compartment in MDA-MB-231 cells. Following treatment, the cell monolayer was rinsed with warm sterile PBS. Cells were trypsinised and centrifuged for 3 min at 1500 rpm. Each pellet was re-suspended in 0.5 mL of the LysoTracker™ Red (Invitrogen™, USA) working solution (final concentration of 100 nmol/L made up in growth medium) and was left to incubate for 15 min at 37 °C. Following incubation, the cells were gently re-suspended before being filtered through a 50μm nylon mesh into FACS tubes and analysed immediately on the flow cytometer. Analyses were performed on a FACSAria I flow cytometer (Becton Dickinson Biosciences, San Jose, CA) equipped with a 488 nm Coherent Sapphire solid state laser (13-20 mW), 633 nm JDS Uniphase HeNe air-cooled laser (10-20 mW) and 407 nm Point Source Violet solid state laser (10-25 mW). For each sample, population information from a minimum of 10 000 events was acquired using FACSDiva Version 6.1 software. Data were expressed in arbitrary values as a percentage relative to the untreated control.
Western blot analysis
After treatment, the cells were placed on ice, whereafter cell monolayers were rinsed with 1 mL cold PBS. Modified radio-immuno precipitation (RIPA) buffer (50 mM Tris-HCL, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM benzamidine, 4 μg/mL SBTI-1, 1 μg/mL leupeptin, 1% NP40, and 0.25% Na-deoxycholate; pH 7.4) was added to each well and was left to incubate on ice for 5 min. The cell suspension was transferred to pre-chilled Eppendorf tubes. Protein concentration was quantified using the Bradford protein method.31 Following protein determination, lysates were diluted in Laemmli's sample buffer, boiled for 5 min and then centrifuged for 5 s. The prepared cell lysates were separated on 10% polyacrylamide gels by sodium dodecyl sulfate polyacrylamide gel electrophoresis at 200 V (Mini Protean System, Bio-Rad, Hercules). Following sodium dodecyl sulfate polyacrylamide gel electrophoresis, proteins were transferred to polyvinylidene fluoride (PVDF) membranes (Immoblin-P, Millipore) using a semi-dry electro-transfer system (Bio-Rad, USA). Membranes were blocked in 5% (w/v) fat-free milk in 0.1% Tris Buffered Saline-Tween20 (TBS-T) for 1 h at room temperature with gentle agitation. Membranes were then incubated overnight at 4 °C in the appropriate primary antibodies, including caspase 3 (Cell Signalling, MA, USA), cleaved PARP (Cell Signalling, MA, USA), LC3 (Cell Signalling, MA, USA), p62 (Cell Signalling, MA, USA) and MKP-1 (Santa Cruz Biotechnology, CA, USA), while p-actin (Cell Signalling, MA, USA) was used as a loading control. The membranes were incubated in anti-rabbit horseradish peroxidase-conjugated secondary antibody (1:10 000), Amersham Biosciences, UK, and Dako Cytomation, Denmark) for 1 h at room temperature the following day. The membranes were developed with the ECL detection Kit (Bio Vision Inc.) and imaged using the CL-Xposure (Thermo Scientific) X-ray film. Exposed bands were quantified by densitometry using the UN-SCAN-IT© densitometry software (Silk Scientific Corporation, Utah, USA). Bands were expressed as optical density readings relative to the untreated control present on the same blot.
Statistical analysis
All values are expressed as a percentage of the control. The results are presented as mean ± standard error of the mean (SEM) for an n = 3. Comparisons between different groups were made by one-way analysis of variance (ANOVA), followed by the Bonferroni post-hoc test. Statistical analyses were performed using GraphPad Prism version 5.01 (Graphpad Software, Inc, CA, USA), and a value of p < 0.05 was considered statistically significant.
Results
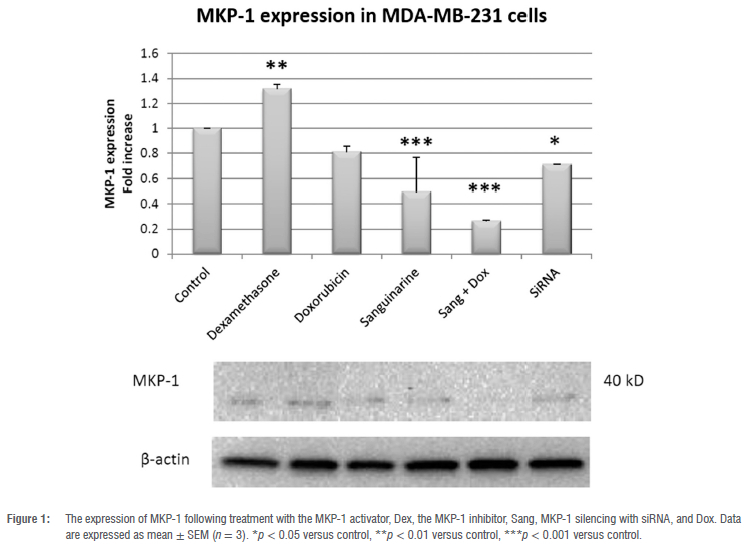
MKP-1 expression modulated by dexamethasone, sanguinarine and siRNA
Treatment with the MKP-1 inducer, Dex (100 nM), for 24 h significantly increased MKP-1 expression in MDA-MB-231 cells (p < 0.01; Figure 1). In contrast, treatment with the MKP-1 inhibitor, Sang (0.5 μM), for 48 h (p < 0.001) and treatment with the MKP-1 silencer, MKP-1 siRNA (20 pmol), for 24 h downregulated MKP-1 expression in vitro (p < 0.05). Additionally, the expression of MKP-1 was also downregulated following co-treatment with Sang and Dox (p < 0.001), but not with Dox alone.
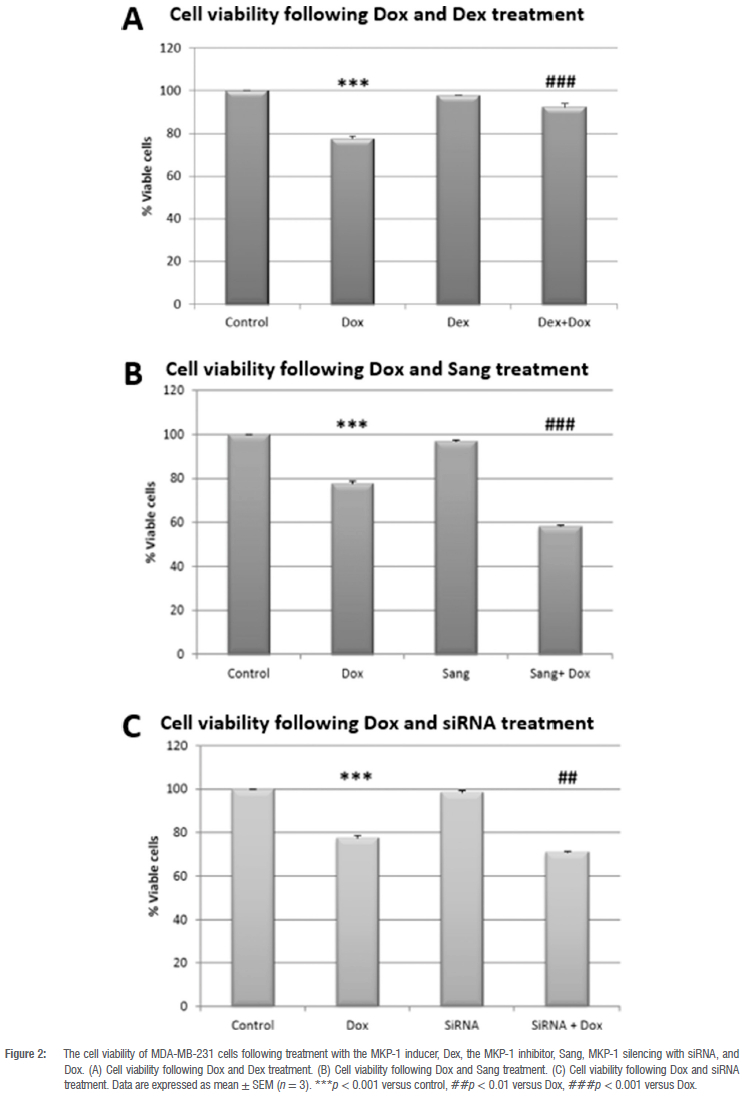
MKP-1 inhibition and silencing in combination with Dox decreases cell viability
Dox treatment significantly decreased cell viability of MDA-MB-231 cells when compared to the control group (p < 0.001; Figure 2A). While treatment with the activator of MKP-1, Dex, alone had no effect on cell viability, the combination of Dex with Dox resulted in a significant increase in cell viability when compared to Dox treatment alone (p < 0.001; Figure 2A). In contrast, treatment with the MKP-1 inhibitor, Sang, alone did not have a significant effect on cell viability (Figure 2B); however, the combination of Dox and Sang resulted in a significant decrease in cell viability (p < 0.001) when compared to Dox treatment alone. Similarly, treatment with MKP-1 siRNA alone resulted in no significant change in cell viability when compared to the control (Figure 2C); however, a significant decrease in cell viability was observed when comparing the combination treatment of MKP-1 siRNA and Dox with Dox treatment alone (p < 0.01; Figure 2C). Therefore, the inhibition of MKP-1 in conjunction with Dox treatment induced a greater reduction of viable cells than Dox alone.
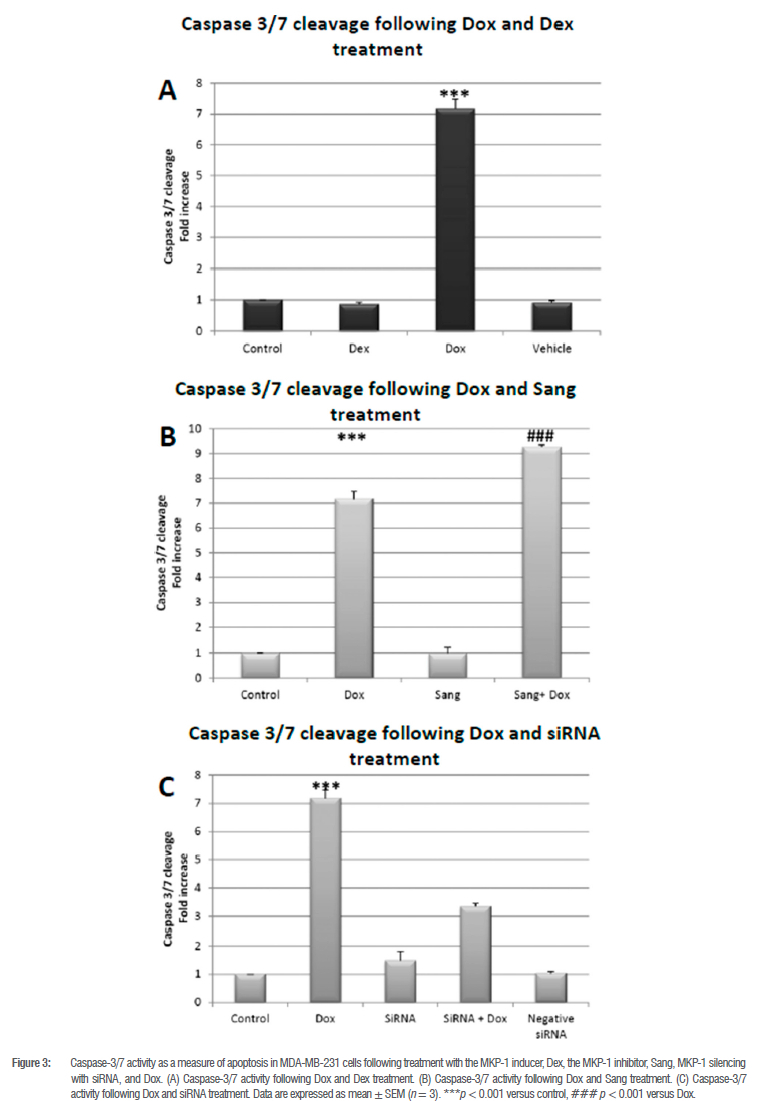
MKP-1 inhibition during doxorubicin treatment promotes caspase 3/7 cleavage
As expected, Dox treatment resulted in a significant increase in caspase 3/7 activity, indicating the execution of apoptotic cell death (p < 0.001; Figure 3A, B and C). Both Dex and Sang as well as MKP-1 siRNA treatment alone had no effect on caspase 3/7 activity, as observed by the cell viability data (Figure 3B). However, under conditions of MKP-1 inhibition with Sang in combination with Dox, there was a significant increase in caspase 3/7 activity compared to Dox alone (p < 0.001; Figure 3B), also indicating that MKP-1 inhibition in conjunction with Dox treatment resulted in apoptotic cell death, which is in line with the cell viability results. However, neither MKP-1 siRNA nor the combination of siRNA with Dox had any significant effect on caspase 3/7 (Figure 3C).
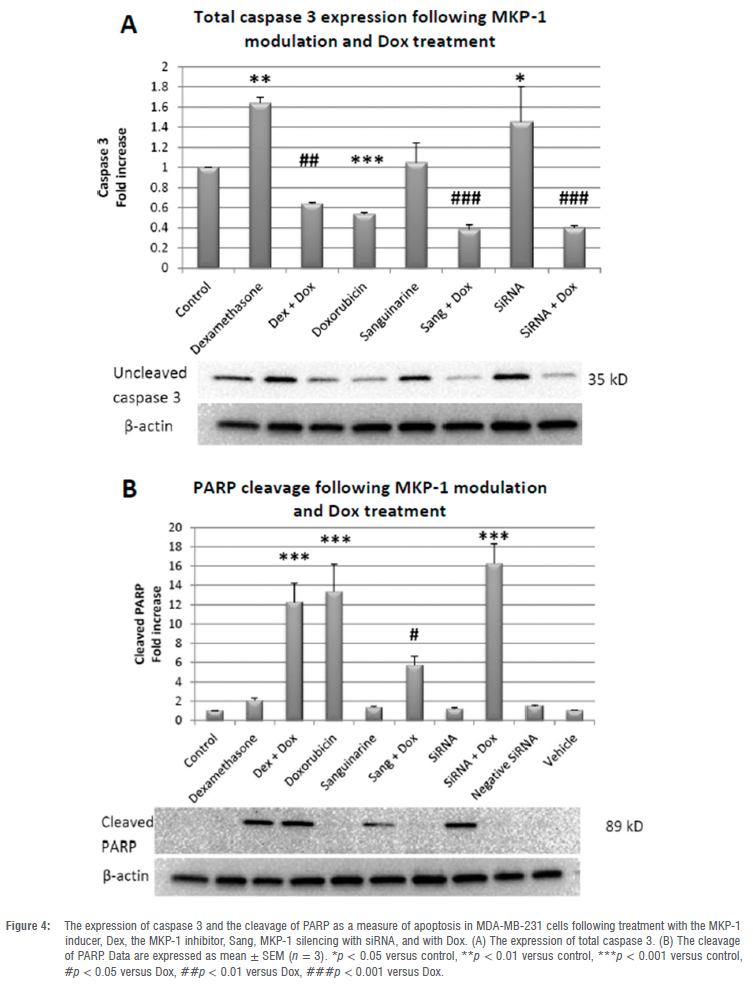
Total caspase 3 and PARP cleavage display differential effects under conditions of MKP-1 induction, inhibition and silencing
The expression of total caspase 3 following Dox treatment was significantly decreased (p < 0.001), while PARP cleavage was increased (p < 0.001), which supports the Caspase-Glo® 3/7 and viability assays. The activator of MKP-1, Dex, significantly increased caspase 3 expression when compared to the control (p < 0.01) without PARP cleavage. In contrast, the combination of Dox and Dex treatment resulted in the decreased expression of caspase 3 (p < 0.01) and increased cleavage of PARP (p < 0.001). Therefore, Dex confers protection against apoptosis, where the addition of Dox abolishes this effect. The inhibitor of MKP-1, Sang, resulted in no significant change in caspase 3 expression or PARP cleavage when compared to the control. However, the co-treatment of Sang and Dox resulted in a significant decrease in caspase 3 expression (p < 0.001) and PARP cleavage (p < 0.01) when compared to the Dox group, which is supported by our findings for the Caspase-Glo® 3/7 and viability assays. The expression of caspase 3 was significantly increased following treatment with MKP-1 siRNA when compared to the control (p < 0.05) with no change in PARP cleavage; however, co-treatment with the MKP-1 silencer and Dox significantly reduced caspase 3 expression (p < 0.001), while increasing PARP cleavage (p < 0.001) when compared to Dox (Figure 4A,B). Therefore, MKP-1 inhibition and silencing sensitise breast cancer cells to apoptosis during Dox treatment.
MKP-1 inhibition and silencing during Dox treatment promotes changes in cell morphology
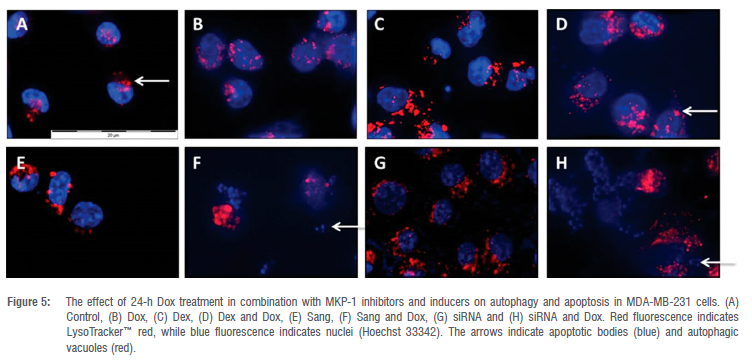
Morphological analysis revealed an increase in the size and number of acidic vesicles in response to both Sang and MKP-1 siRNA (Figure 5B), as well as in cells treated with both Sang and Dox (Figure 5F) or MKP-1 siRNA and Dox (Figure 5H). An increase in cell death markers, such as pyknosis and apoptotic bodies, was observed in the groups treated with Dex and Dox (Figure 5D) or MKP-1 siRNA and Dox (Figure 5H) where the highest degree of cell death was seen in the cells treated with Sang in combination with Dox (Figure 5F).
MKP-1 inhibition promotes autophagy
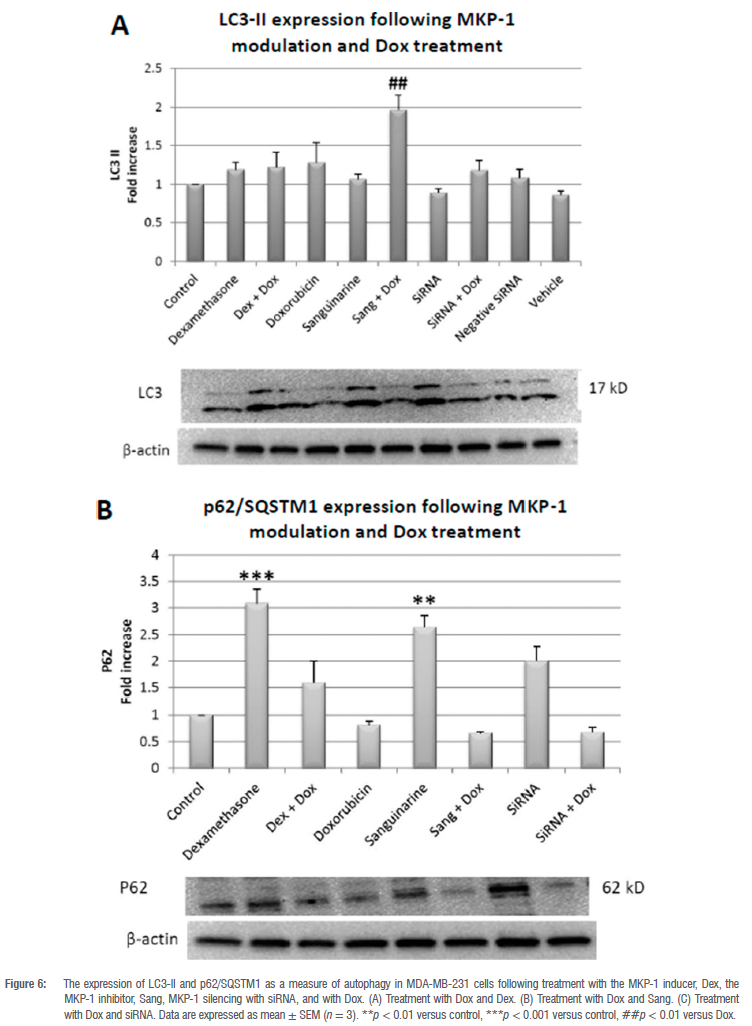
A significant increase in the conversion of LC3-1 to LC3-II was observed when MKP-1 was inhibited with Sang during Dox treatment when compared to Dox treatment alone (p < 0.01; Figure 6A). Furthermore, the expression of the cargo recruiter, p62/SQSTM1, was significantly increased in the Dex (p < 0.001) and Sang (p < 0.01) treated groups when compared to the control (Figure 6B). Evident in the increased conversion of LC3-1 to LC3-II and the twofold reduction in p62 expression, autophagy was induced.
Lysosomal acidic compartments undergo differential changes under conditions of MKP-1 induction, inhibition and silencing during Dox treatment
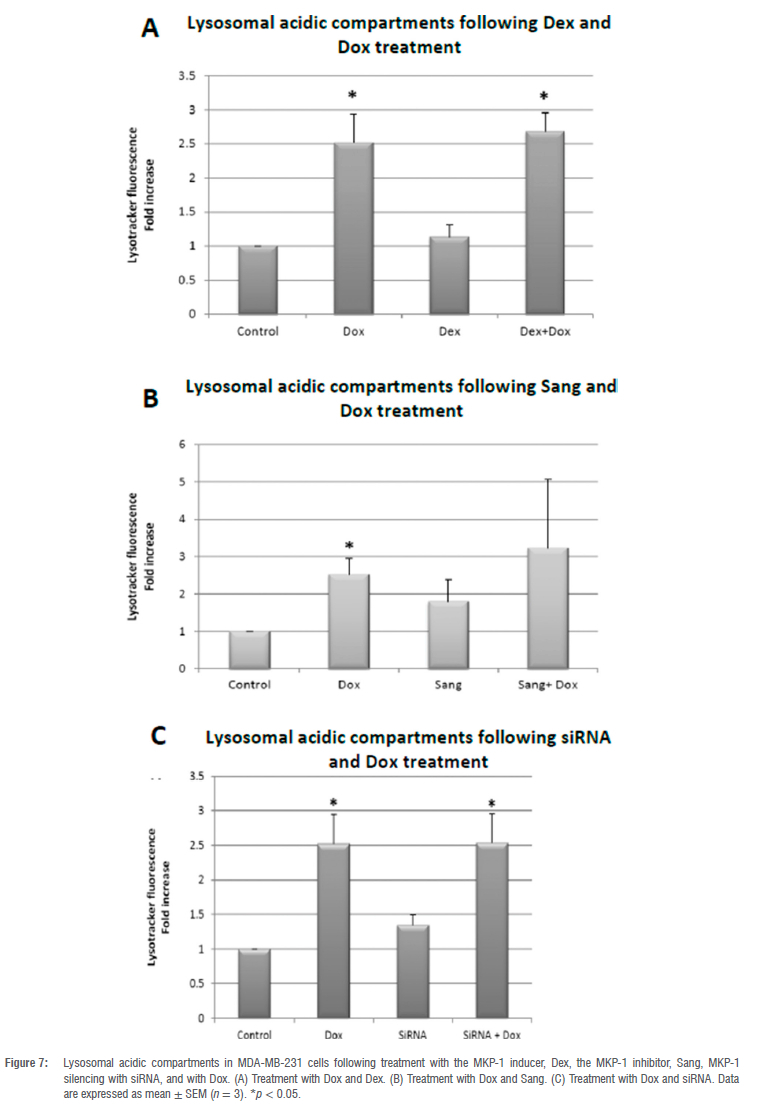
Treatment with Dox significantly increased lysosomal acidic compartments compared to the control (p < 0.05; Figure 7A). Dex treatment, a known MKP-1 inducer, had no effect on lysosomal acidic compartments compared to the control. However, the combination treatment of Dex and Dox resulted in a significant increase in lysosomal acidic compartments compared to the control (p < 0.05), although to the same extent as Dox treatment alone (Figure 7A). Sang treatment alone and in combination with Dox did not result in a significant change in lysosomal acidic compartments (Figure 7B). MKP-1 siRNA resulted in no significant change in lysosomal acidic compartments when compared to the control (Figure 7C). However, a significant (p < 0.05) increase in lysosomal acidic compartments was observed with Dox in conjunction with MKP-1 siRNA compared to control.
Discussion
It is well established that MKP-1 is associated with cancer progression and resistance to chemotherapy.13,14 It has been reported that MKP-1 is overexpressed by more than fivefold in primary samples from breast cancer patients.32-34 We, therefore, investigated whether doxorubicin treatment and treatment with the MKP-1 inhibitor, sanguinarine, and silencing of MKP-1 using siRNA can inhibit MKP-1 expression in MDA-MB-231 cells (Figure 1). Doxorubicin treatment did not alter the expression levels of MKP-1, which is in contrast to a study performed by Rojo et al.35 who reported that doxorubicin decreased MKP-1 expression in a time- and dose-dependent manner. However, treatment with the glucocorticoid dexamethasone significantly increased MKP-1 expression in the MDA-MB-231 cells. This is supported by studies that confirmed that dexamethasone treatment is an inducer of MKP-1 expression.36,37 Additionally, we investigated whether the bioactive plant extract sanguinarine can inhibit MKP-1 expression in breast cancer cells. Treatment with sanguinarine significantly decreased the expression of MKP-1 in triple negative breast cancer cells. This is supported by a study performed by Vogt et al.26 in which sanguinarine treatment inhibited MKP-1 expression in PANC-1 and HeLa cells. Furthermore, the silencing of MKP-1 with MKP-1 siRNA also significantly decreased the expression of MKP-1 in vitro. This confirmed dexamethasone as an inducer of MKP-1 expression, while sanguinarine and MKP-1 siRNA effectively inhibited MKP-1 expression. We also found that co-treatment with sanguinarine and doxorubicin inhibits the expression of MKP-1, which is supported by existing reports that both doxorubicin and sanguinarine are inhibitors of MKP-1 expression.10,26,35
Based on these findings, we investigated the effects of MKP-1 modulation on the viability and apoptosis of breast cancer cells following doxorubicin treatment in vitro. Doxorubicin treatment significantly decreased cell viability of breast cancer cells (Figure 2A-C), which is supported by our findings of the increased activity of caspase 3 and 7 in vitro (Figure 3A). Furthermore, doxorubicin treatment significantly decreased total caspase 3 and increased PARP cleavage (Figure 4A,B). It is therefore concluded that doxorubicin induced apoptosis in vitro in our model. The sensitivity of these cell lines to doxorubicin has also been well established where increased levels of apoptosis are evident following doxorubicin treatment.38-40 We then investigated whether the modulation of MKP-1 during doxorubicin treatment promotes or inhibits apoptosis induction in breast cancer cells. MKP-1 upregulation by dexamethasone alone did not affect cell viability or induce apoptosis (Figure 2A, 3A and 4), which is supported by previous studies which reported that glucocorticoids protect mammary gland epithelial cells, hepatocytes and ovarian follicular cells against cell death signals and apoptosis.41 This protective effect may be mediated by the upregulation of MKP-1, which can inhibit MAPK-induced apoptosis. However, co-treatment with dexamethasone and doxorubicin promoted cell death, indicating that the upregulation of MKP-1 alone confers a protective effect against apoptosis, but doxorubicin treatment abolishes this effect. Furthermore, the inhibition of MKP-1 with sanguinarine alone did not affect breast cancer cell apoptosis (Figure 2B, 3B and 4A-B). This suggests that the inhibition of MKP-1 alone is not an effective inducer of apoptosis. However, it has been reported that sanguinarine inhibits cancer cell proliferation with different degrees of sensitivity in different triple negative breast cancer cell lines.42,43 Co-treatment with sanguinarine and doxorubicin decreased the amount of viable cancer cells and induced apoptosis (Figure 2B, 3B and 4A-B). These results indicate that sanguinarine improves the efficacy of doxorubicin treatment by sensitising breast cancer cells to doxorubicin-induced cell death in vitro. This is supported by studies showing that MKP-1 inhibition induced apoptosis in cervical and renal cell carcinomas16, as well as in mesangial cells44. As with sanguinarine treatment, the silencing of MKP-1 with MKP-1 siRNA alone did not affect apoptosis (Figure 2C, 3C and 4B). However, the co-administration of doxorubicin and siRNA significantly decreased cell viability and induced apoptosis. This supports our findings for sanguinarine and doxorubicin co-treatment. Furthermore, Wang et al.45 reported that the inhibition of MKP-1 sensitised ovarian cancer cells to cisplatin-induced apoptosis, which is mediated through the activation of the MAPKs. Therefore, the inhibition of MKP-1 during doxorubicin treatment may have important therapeutic value. Various chemotherapeutic agents induce apoptosis through the activation of the MAPKs, namely ERK, JNK and p38. These MAPKs are negatively regulated by MKP-1. MAPKs are involved in apoptosis induction, and their activation by chemotherapies induces cytochrome c release from the mitochondria. This results in the oligomerisation of apoptotic protease activating factor-1 (Apaf-1), recruiting pro-caspase-9 and inducing its autoactivation, which results in the downstream activation of the effector caspase 3.46 Furthermore, JNK family members are involved in the activation of apoptosis by transcription-dependent processes, including death receptor induction.47 Therefore, loss or inhibition of MKP-1 reduces its inhibition on members of the MAPK family, which may facilitate apoptosis following doxorubicin treatment. Furthermore, Small and co-authors10 reported that MKP-1 inhibition enhanced chemosensitivity to doxorubicin through the activation of JNK. To further validate our findings for apoptosis, morphological changes commonly associated with the progression of apoptosis, such as chromatin condensation and the formation of apoptotic bodies, were also investigated (Figure 5A-H). The doxorubicin-treated groups displayed increased morphological characteristics that are associated with cell death, including pyknosis and the formation of apoptotic bodies. Co-treatment with sanguinarine and doxorubicin displayed the highest degree of apoptosis induction when compared to the other treatment groups. This further supports our findings that under conditions of MKP-1 inhibition, MDA-MB-231 cells are sensitised to doxorubicin-induced apoptosis.
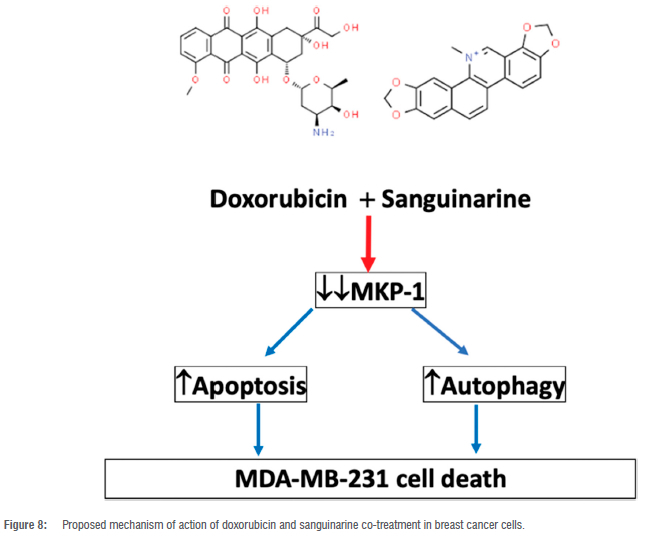
Autophagy has been shown to play a dual role in doxorubicin-induced cell death.48 We therefore assessed whether the modulation of MKP-1 and doxorubicin treatment can regulate autophagy. The co-administration of sanguinarine and doxorubicin induced autophagy in vitro (Figure 6A,B). This may be a synergistic effect between sanguinarine and doxorubicin, as treatment with sanguinarine or doxorubicin alone was not sufficient to induce autophagy. In line with this, sanguinarine has been reported to induce ROS-dependent autophagy and apoptosis in hepatocellular carcinoma cells.49 This may be mediated through the resulting upregulation of MAPK activity following MKP-1 inhibition. The MAPK family are known regulators of autophagy, either inducing or inhibiting autophagy. It has been reported that ERK activation can induce autophagy by promoting the dissociation of Beclin-1 and Bcl-2, through the direct activation of the Ga-interacting protein (GAIP) and through the activation of ATF4, resulting in the increased transcription of the ATG proteins.50-52 Furthermore, p38 activation can also induce autophagy through the activation of the ULK complex and LAMP2A translocation to the lysosomal membrane.53,54 Lastly, JNK activation promotes autophagy through the dissociation of Beclin-1 and Bcl-2, the direct activation of Atg5 and Atg7, and by inducing Sestrin2 transcription, which activates AMPK.55 Furthermore, at low doses, doxorubicin elicits an autophagic response in breast cancer cells.56 Additionally, increased levels of ATG2A mRNA have been observed following doxorubicin treatment57, suggesting increased autophagy activation at the transcriptional level. Conversely, activation of autophagy in cultured cardiomyocytes following 1 doxorubicin treatment mediated its cardiotoxic effect.58 Here, it was shown that autophagy inhibition resulted in decreased cell death, and it was postulated that autophagy directly contributed to doxorubicin-induced toxicity. Apoptosis and autophagy are finely regulated, and MKP-1 inhibition with sanguinarine may promote doxorubicin-induced apoptosis and autophagy as an anti-neoplastic mechanism. However, only doxorubicin, irrespective of MKP-1 modulation, induced an increase in lysosomal acidity (Figure 7A-C). This finding is in line with a study performed by Thomas et al.59 who showed that doxorubicin increased lysosomal acidity in MDA-MB-231 cells. Our results are also supported by the findings of other authors which show that doxorubicin induces an autophagic response in cancer cells.60 Figure 8 summarises the main findings of this study and the proposed mechanism of action of doxorubicin and sanguinarine in breast cancer cells.
Conclusions
We have reported a role for MKP-1 modulation during doxorubicin treatment in vitro. It is clear that both apoptosis and autophagy remain central in tumourigenesis. In conclusion, the findings of this study clearly indicate the significant role of MKP-1 in cancer treatment and the importance of modulating MKP-1 in a targeted way in cancer patients receiving chemotherapy.
Future perspectives
The role of MKP-1 in autophagy induction should be further explored to elucidate how the upregulation of autophagy can contribute to apoptosis. Future studies should assess autophagic flux over time and include autophagy inhibitors to quantify autophagosomal accumulation.
Acknowledgements
This work was supported by the National Research Foundation of South Africa (grant number 118566), the Cancer Association of South Africa and the South African Medical Research Council.
Competing interests
We have no competing interests to declare.
Authors' contributions
M.d.P: Writing - original draft, writing - review and editing. C.F.: Writing -review and editing. HI: Methodology, data collection, validation, sample and data analysis, data curation. A-M.E.: Conceptualisation, resources, writing - review and editing, supervision, funding acquisition, project leadership.
References
1. Mansoori B, Mohammadi A, Davudian S, Shirjang S, Baradaran B. The different mechanisms of cancer drug resistance: A brief review. Adv Pharm Bull. 2017;7(3):339-348. https://doi.org/10.15171/apb.2017.041 [ Links ]
2. Crook T, Gaya A, Page R, Limaye S, Ranade A, Bhatt A, et al. Clinical utility of circulating tumor-associated cells to predict and monitor chemo-response in solid tumors. Cancer Chemother Pharmacol. 2021;87(2):197-205. https://doi.org/10.1007/s00280-020-04189-8 [ Links ]
3. Thorn CF, Oshiro C, Marsh S, Hernandez-Boussard T, McLeod H, Klein TE, et al. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharmacogenet Genomics. 2011;21(7):440-446. https://doi.org/10.1097/FPC.0b013e32833ffb56 [ Links ]
4. Ferreira AL, Matsubara LS, Matsubara BB. Anthracycline-induced cardiotoxicity. Cardiovasc Hematol Agents Med Chem. 2008;6(4):278-281. https://doi.org/10.2174/187152508785909474 [ Links ]
5. Taymaz-Nikerel H, Karabekmez ME, Eraslan S, Kirdar B. Doxorubicin induces an extensive transcriptional and metabolic rewiring in yeast cells. Sci Rep. 2018;8(1):13672. https://doi.org/10.1038/s41598-018-31939-9 [ Links ]
6. Octavia Y Tocchetti CG, Gabrielson KL, Janssens S, Crijns HJ, Moens AL. Doxorubicin-induced cardiomyopathy: From molecular mechanisms to therapeutic strategies. J Mol Cell Cardiol. 2012;52(6):1213-1225. https://doi.org/10.1016/j.yjmcc.2012.03.006 [ Links ]
7. Tacar O, Sriamornsak P Dass CR. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J Pharm Pharmacol. 2013;65(2):157-170. https://doi.org/10.1111/j.2042-7158.2012.01567.x [ Links ]
8. Farooq A, Zhou MM. Structure and regulation of MAPK phosphatases. Cell Signal. 2004;16(7):769-779. https://doi.org/10.1016/j.cellsig.2003.12.008 [ Links ]
9. Keyse SM. Dual-specificity MAP kinase phosphatases (MKPs) and cancer. Cancer Metastasis Rev. 2008;27(2):253-261. https://doi.org/10.1007/s10555-008-9123-1 [ Links ]
10. Small GW, Shi YY Higgins LS, Orlowski RZ. Mitogen-activated protein kinase phosphatase-1 is a mediator of breast cancer chemoresistance. Cancer Res. 2007;67(9):4459-4466. https://doi.org/10.1158/0008-5472.CAN-06-2644 [ Links ]
11. Vogt A, McDonald PR, Tamewitz A, Sikorski RP, Wipf P, Skoko JJ 3rd, et al. A cell-active inhibitor of mitogen-activated protein kinase phosphatases restores paclitaxel-induced apoptosis in dexamethasone-protected cancer cells. Mol Cancer Ther. 2008;7(2):330-340. https://doi.org/10.1158/1535-7163.MCT-07-2165 [ Links ]
12. Awada A, Albanell J, Canney PA, Dirix LY Gil T, Cardoso F, et al. Bortezomib/ docetaxel combination therapy in patients with anthracycline-pretreated advanced/metastatic breast cancer: A phase I/II dose-escalation study. Br J Cancer. 2008;98(9):1500-1507. https://doi.org/10.1038/sj.bjc.6604347 [ Links ]
13. Ma G, Pan Y, Zhou C, Sun R, Bai J, Liu P et al. Mitogen-activated protein kinase phosphatase 1 is involved in tamoxifen resistance in MCF7 cells. Oncol Rep. 2015;34(5):2423-2430. https://doi.org/10.3892/or.2015.4244 [ Links ]
14. Lei S, Xu H, Chen N, Pan H, Xie W, He Y, et al. MKP-1 overexpression is associated with chemoresistance in bladder cancer via the MAPK pathway. Oncol Lett. 2020;20(2):1743-1751. https://doi.org/10.3892/ol.2020.11741 [ Links ]
15. Shi YY, Small GW, Orlowski RZ. Proteasome inhibitors induce a p38 mitogen-activated protein kinase (MAPK)-dependent anti-apoptotic program involving MAPK phosphatase-1 and Akt in models of breast cancer. Breast Cancer Res Treat. 2006;100(1):33-17. https://doi.org/10.1007/s10549-006-9232-x [ Links ]
16. Mizuno R, Oya M, Shiomi T Marumo K, Okada Y, Murai M. Inhibition of MKP-1 expression potentiates JNK related apoptosis in renal cancer cells. J Urol. 2004;172(2):723-727. https://doi.org/10.1097/01.ju.0000124990.37563.00 [ Links ]
17. Franklin CC, Srikanth S, Kraft AS. Conditional expression of mitogen-activated protein kinase phosphatase-1, MKP-1, is cytoprotective against UV-induced apoptosis. Proc Natl Acad Sci USA. 1998;95(6):3014-3019. https://doi.org/10.1073/pnas.95.6.3014 [ Links ]
18. Fu XH, Chen CZ, Li S, Han DX, Wang YJ, Yuan B, et al. Dual-specificity phosphatase 1 regulates cell cycle progression and apoptosis in cumulus cells by affecting mitochondrial function, oxidative stress, and autophagy. Am J Physiol Cell Physiol. 2019;317(6):C1183-C1193. https://doi.org/10.1152/ajpcell.00012.2019 [ Links ]
19. Degenhardt K, Mathew R, Beaudoin B, Bray K, Anderson D, Chen G, et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell. 2006;10(1):51-64. https://doi.org/10.1016/j.ccr.2006.06.001 [ Links ]
20. Dasari SK, Bialik S, Levin-Zaidman S, Levin-Salomon V, Merrill AH Jr, Futerman AH, et al. Signalome-wide RNAi screen identifies GBA1 as a positive mediator of autophagic cell death. Cell Death Differ. 2017;24(7):1288-1302. https://doi.org/10.1038/cdd.2017.80 [ Links ]
21. Liu PF, Tsai KL, Hsu CJ, Tsai WL, Cheng JS, Chang HW, et al. Drug repurposing screening identifies tioconazole as an Atg4 inhibitor that suppresses autophagy and sensitizes cancer cells to chemotherapy. Theranostics. 2018;8(3):830-845. https://doi.org/10.7150/thno.22012 [ Links ]
22. Aburai N, Yoshida M, Ohnishi M, Kimura K. Sanguinarine as a potent and specific inhibitor of protein phosphatase 2C in vitro and induces apoptosis via phosphorylation of p38 in HL60 cells. Biosci Biotechnol Biochem. 2010;74(3):548-552. https://doi.org/10.1271/bbb.90735 [ Links ]
23. Jang BC, Park JG, Song DK, Baek WK, Yoo SK, Jung KH, et al. Sanguinarine induces apoptosis in A549 human lung cancer cells primarily via cellular glutathione depletion. Toxicol In Vitro. 2009;23(2):281-287. https://doi.org/10.1016/j.tiv.2008.12.013 [ Links ]
24. Park H, Bergeron E, Senta H, Guillemette K, Beauvais S, Blouin R, et al. Sanguinarine induces apoptosis of human osteosarcoma cells through the extrinsic and intrinsic pathways. Biochem Biophys Res Commun. 2010;399(3):446-451. https://doi.org/10.1016/j.bbrc.2010.07.114 [ Links ]
25. Serafim TL, Matos JA, Sardão VA, Pereira GC, Branco AF, Pereira SL, et al. Sanguinarine cytotoxicity on mouse melanoma K1735-M2 cells-nuclear vs. mitochondrial effects. Biochem Pharmacol. 2008;76(11):1459-1475. https://doi.org/10.1016/j.bcp.2008.07.013 [ Links ]
26. Vogt A, Tamewitz A, Skoko J, Sikorski RP Giuliano KA, Lazo JS. The benzo[c]phenanthridine alkaloid, sanguinarine, is a selective, cell-active inhibitor of mitogen-activated protein kinase phosphatase-1. J Biol Chem. 2005;280(19):19078-19086. https://doi.org/10.1074/jbc.M501467200 [ Links ]
27. Adhami VM, Aziz MH, Reagan-Shaw SR, Nihal M, Mukhtar H, Ahmad N. Sanguinarine causes cell cycle blockade and apoptosis of human prostate carcinoma cells via modulation of cyclin kinase inhibitor-cyclin-cyclin-dependent kinase machinery. Mol Cancer Ther. 2004;3(8):933-940. https://doi.org/10.1158/1535-7163.933.3.8 [ Links ]
28. Xu J, Meng Q, Chong Y Jiao Y Zhao L, Rosen EM. Sanguinarine inhibits growth of human cervical cancer cells through the induction of apoptosis. Oncol Rep. 2012;28:2264-2270. https://doi.org/10.3892/or.2012.2024 [ Links ]
29. Ahsan H, Reagan-Shaw S, Breur J, Ahmad N. Sanguinarine induces apoptosis of human pancreatic carcinoma AsPC-1 and BxPC-3 cells via modulations in Bcl-2 family proteins. Cancer Lett. 2007;249(2):198-208. https://doi.org/10.1016/j.canlet.2006.08.018 [ Links ]
30. Kitakaze M, Fong M, Yoshitake M, Minamino T, Node K, Okuyama Y, et al. Vesnarinone inhibits adenosine uptake in endothelial cells, smooth muscle cells and myocytes, and mediates cytoprotection. J Mol Cell Cardiol. 1997;29(12):3413-3417. https://doi.org/10.1006/jmcc.1997.0581 [ Links ]
31. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248-254. https://doi.org/10.1006/abio.1976.9999 [ Links ]
32. Wang HY, Cheng Z, Malbon CC. Overexpression of mitogen-activated protein kinase phosphatases MKP1, MKP2 in human breast cancer. Cancer Lett. 2003;191(2):229-237. https://doi.org/10.1016/S0304-3835(02)00612-2 [ Links ]
33. Loda M, Capodieci P Mishra R, Yao H, Corless C, Grigioni W, et al. Expression of mitogen-activated protein kinase phosphatase-1 in the early phases of human epithelial carcinogenesis. Am J Pathol. 1996;149(5):1553-1564. [ Links ]
34. Kurt RA, Urba WJ, Smith JW, Schoof DD. Peripheral T lymphocytes from women with breast cancer exhibit abnormal protein expression of several signalling molecules. Int J Cancer. 1998;78(1):16-20. https://doi.org/10.1002/(SICI)1097-0215(19980925)78:1 <16::AID-IJC4>3.0.CO;2-# [ Links ]
35. Rojo F, González-Navarrete I, Bragado R, Dalmases A, Menéndez S, Cortes-Sempere M, et al. Mitogen-activated protein kinase phosphatase-1 in human breast cancer independently predicts prognosis and is repressed by doxorubicin. Clin Cancer Res. 2009;15(10):3530-3539. https://doi.org/10_1158/1078-0432.CCR-08-2070 [ Links ]
36. Chen P, Li J, Barnes J, Kokkonen GC, Lee JC, Liu Y. Restraint of proinflammatory cytokine biosynthesis by mitogen-activated protein kinase phosphatase-1 in lipopolysaccharide-stimulated macrophages. J Immunol. 2002;169(11):6408-6416. https://doi.org/10.4049/jimmunol.169.11.6408 [ Links ]
37. Clark AR. MAP kinase phosphatase 1: A novel mediator of biological effects of glucocorticoids? J Endocrinol. 2003;178(1):5-12. https://doi.org/10.1677/joe.0.1780005 [ Links ]
38. Aroui S, Brahim S, De Waard M, Bréard J, Kenani A. Efficient induction of apoptosis by doxorubicin coupled to cell-penetrating peptides compared to unconjugated doxorubicin in the human breast cancer cell line MDA-MB 231. Cancer Lett. 2009;285(1):28-38. https://doi.org/10.1016/j.canlet.2009.04.044 [ Links ]
39. Kim TH, Shin YJ, Won AJ, Lee BM, Choi WS, Jung JH, et al. Resveratrol enhances chemosensitivity of doxorubicin in multidrug-resistant human breast cancer cells via increased cellular influx of doxorubicin. Biochim Biophys Acta. 2014;1840(1):615-625. https://doi.org/10.1016/j.bbagen.2013.10.023 [ Links ]
40. Mussi SV, Sawant R, Perche F, Oliveira MC, Azevedo RB, Ferreira LA, et al. Novel nanostructured lipid carrier co-loaded with doxorubicin and docosahexaenoic acid demonstrates enhanced in vitro activity and overcomes drug resistance in MCF-7/Adr cells. Pharm Res. 2014;31(8):1882-1892. https://doi.org/10.1007/s11095-013-1290-2 [ Links ]
41. Redondo M, Téllez T, Roldan MJ, Serrano A, García-Aranda M, Gleave ME, et al. Anticlusterin treatment of breast cancer cells increases the sensitivities of chemotherapy and tamoxifen and counteracts the inhibitory action of dexamethasone on chemotherapy-induced cytotoxicity. Breast Cancer Res. 2007;9(6):R86. https://doi.org/10.1186/bcr1835 [ Links ]
42. Ghauri MA, Su Q, Ullah A, Wang J, Sarwar A, Wu Q, et al. Sanguinarine impedes metastasis and causes inversion of epithelial to mesenchymal transition in breast cancer. Phytomedicine. 2021;84, Art. #153500. https://doi.org/10.1016/j.phymed.2021.153500 [ Links ]
43. Messeha S, Zarmouh N, Noel S, Soliman K. Gene expression alterations associated with sanguinarine -induced antiproliferative effect and apoptosis in triple-negative breast cancer cells. FASEB J. 2021;35(S1). https://doi.org/10.1096/fasebj.2021.35.S1.00463 [ Links ]
44. Xu Q, Konta T, Nakayama K, Furusu A, Moreno-Manzano V Lucio-Cazana J, et al. Cellular defense against H2O2-induced apoptosis via MAP kinase-MKP-1 pathway. Free Radic Biol Med. 2004;36(8):985-993. https://doi.org/10.1016/j.freeradbiomed.2004.01.009 [ Links ]
45. Wang Z, Xu J, Zhou JY, Liu Y Wu GS. Mitogen-activated protein kinase phosphatase-1 is required for cisplatin resistance. Cancer Res. 2006;66(17): 8870-8877. https://doi.org/10.1158/0008-5472.CAN-06-1280 [ Links ]
46. Aoki H, Kang PM, Hampe J, Yoshimura K, Noma T, Matsuzaki M, et al. Direct activation of mitochondrial apoptosis machinery by c-Jun N-terminal kinase in adult cardiac myocytes. J Biol Chem. 2002;277(12):10244-10250. https://doi.org/10.1074/jbc.M112355200 [ Links ]
47. Roos WP Kaina B. DNA damage-induced cell death by apoptosis. Trends Mol Med. 2006;12(9):440-450. https://doi.org/10.1016/j.molmed.2006.07.007 [ Links ]
48. Koleini N, Kardami E. Autophagy and mitophagy in the context of doxorubicin-induced cardiotoxicity. Oncotarget. 2017;8(28):46663-46680. https://doi.org/10.18632/oncotarget.16944 [ Links ]
49. Wang J, Su Q, Wu Q, Chen K, Ullah A, Ghauri MA, et al. Sanguinarine impairs lysosomal function and induces ROS-dependent mitophagy and apoptosis in human hepatocellular carcinoma cells. Arch Pharm Res. 2021;44(11):1025-1036. https://doi.org/10.1007/s12272-021-01356-0 [ Links ]
50. Feng W, Huang S, Wu H, Zhang M. Molecular basis of Bcl-xLs target recognition versatility revealed by the structure of Bcl-xL in complex with the BH3 domain of Beclin-1. J Mol Biol. 2007;372(1):223-235. https://doi.org/10.1016/j.jmb.2007.06.069 [ Links ]
51. Shin S, Buel GR, Wolgamott L, Plas DR, Asara JM, Blenis J, et al. ERK2 mediates metabolic stress response to regulate cell fate. Mol Cell. 2015;59(3):382-398. https://doi.org/10.1016/j-molcel.2015.06.020 [ Links ]
52. Zheng W, Xie W, Yin D, Luo R, Liu M, Guo F. ATG5 and ATG7 induced autophagy interplays with UPR via PERK signaling. Cell Commun Signal. 2019;17(1):42. https://doi.org/10.1186/s12964-019-0353-3 [ Links ]
53. Slobodnyuk K, Radic N, Ivanova S, Llado A, Trempolec N, Zorzano A, et al. Autophagy-induced senescence is regulated by p38a ignalling. Cell Death Dis. 2019;10(6):376. https://doi.org/10.1038/s41419-019-1607-0 [ Links ]
54. Li W, Zhu J, Dou J, She H, Tao K, Xu H, et al. Phosphorylation of LAMP2A by p38 MAPK couples ER stress to chaperone-mediated autophagy. Nat Commun. 2017;8(1):1763. https://doi.org/10.1038/s41467-017-01609-x [ Links ]
55. Zhang XY, Wu XQ, Deng R, Sun T, Feng GK, Zhu XF. Upregulation of sestrin 2 expression via JNK pathway activation contributes to autophagy induction in cancer cells. Cell Signal. 2013;25(1):150-158. https://doi.org/10.1016/j.cellsig.2012.09.004 [ Links ]
56. Akar U, Chaves-Reyez A, Barria M, Tari A, Sanguino A, Kondo Y et al. Silencing of Bcl-2 expression by small interfering RNA induces autophagic cell death in MCF-7 breast cancer cells. Autophagy. 2008;4(5):669-679. https://doi.org/10.4161/auto.6083 [ Links ]
57. Levy JM, Thorburn A. Targeting autophagy during cancer therapy to improve clinical outcomes. Pharmacol Ther. 2011;131(1):130-141. https://doi.org/10.1016/j.pharmthera.2011.03.009 [ Links ]
58. Kobayashi S, Volden P Timm D, Mao K, Xu X, Liang Q. Transcription factor GATA4 inhibits doxorubicin-induced autophagy and cardiomyocyte death. J Biol Chem. 2010;285(1):793-804. https://doi.org/10.1074/jbc.M109.070037 [ Links ]
59. Thomas M, Davis T, Nell T, Sishi B, Engelbrecht AM. Amino acid starvation sensitizes resistant breast cancer to doxorubicin-induced cell death. Front Cell Dev Biol. 2020;8, Art. #565915. https://doi.org/10.3389/fcell.2020.565915 [ Links ]
60. Lambert LA, Qiao N, Hunt KK, Lambert DH, Mills GB, Meijer L, et al. Autophagy: A novel mechanism of synergistic cytotoxicity between doxorubicin and roscovitine in a sarcoma model. Cancer Res. 2008;68(19):7966-7974. https://doi.org/10.1158/0008-5472.CAN-08-1333 [ Links ]
 Correspondence:
Correspondence:
Manisha du Plessis
Email: manishadup@sun.ac.za
Received: 03 Oct. 2022
Revised: 08 Nov. 2023
Accepted: 13 Nov. 2023
Published: 27 Mar. 2024
Editors: Pascal Bessong, Shane Redelinghuys
Funding: South African National Research Foundation (118566), Cancer Association of South Africa, South African Medical Research Council
Supplementary Data
The supplementary data is available in pdf: [Supplementary data]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}