










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.119 n.1-2 Pretoria Jan./Feb. 2023
http://dx.doi.org/10.17159/sajs.2023/15108
COMMENTARY
Immunological interaction during helminth and HIV co-infection: Integrative research needs for sub-Saharan Africa
Miranda N. Mpaka-MbathaI, II, III; Pragalathan NaidooII, III; Md Mazharul IslamIV; Ravesh SinghV; Khethiwe N. BhenguI, II; Nomzamo Nembe-MafaII; Roxanne PillayI, II, III; Zamathombeni DumaII, III, V; Abraham J. NiehausVI; Zilungile L. Mkhize-KwitshanaII, III
IDepartment of Biomedical Sciences, Mangosuthu University of Technology, Durban, South Africa
IIDepartment of Medical Microbiology, School of Laboratory Medicine and Medical Sciences, College of Health Sciences, Nelson R. Mandela Medical School, University of KwaZulu-Natal, Durban, South Africa
IIIDivision of Research Capacity Development, South African Medical Research Council (SAMRC), Cape Town, South Africa
IVDepartment of Animal Resources, Ministry of Municipality, Doha, Qatar
VDepartment of Medical Microbiology, School of Laboratory Medicine and Medical Sciences, College of Health Sciences, Howard College, University of KwaZulu-Natal, Durban, South Africa
VIDepartment of Microbiology, Ampath Laboratories, Cape Town, South Africa
ABSTRACT
SIGNIFICANCE: Sub-Saharan Africa is heavily burdened with human immunodeficiency virus (HIV) and helminth infections, which are potent activators of pro-inflammatory and anti-inflammatory immune responses, respectively. Considering that helminths (such as Necator americanus, Ancylostoma duodenale, Ascaris lumbricoides, Trichuris trichiura, Schistosoma haematobium, and Schistosoma mansoni) can potentially dampen the production and expression of vital anti-viral pro-inflammatory cytokines, leading to enhanced HIV replication and severity, well-designed intervention studies are needed which will offer conclusive data on the nature of these interactions and the impact of deworming in HIV-infected patients. Such research will impact governmental health policies and deworming programmes, allowing for the implementation of integrated systems that will contribute to the overall improvement of African health systems
Keywords: helminths, HIV, co-infection, immunology, intervention programmes
Background
Sub-Saharan Africa has an extensive epidemiological overlap between parasitic helminth infections and human immunodeficiency virus/acquired immune deficiency syndrome (HIV/AIDS).1 According to the World Health Organization, more than 1.5 billion people worldwide are infected with soil-transmitted helminths, and sub-Saharan Africa has the highest burden.2 It is also estimated that 90% of people in need of schistosomiasis treatment live in Africa.3 Approximately 85% of helminth infections in sub-Saharan Africa occur in areas exposed to poverty, overcrowding, inadequate sanitation, poor hygiene practices and unsafe water sources. These living conditions also promote the spread of HIV.1 The sub-tropical climate in sub-Saharan Africa also contributes to high transmission and infection rates.
During the early stages of helminth infection and pre-adult worm development, epithelial cells secrete thymic stromal lymphopoietin, interleukin (IL)-33 and IL-25, which stimulate the activation and differentiation of type 2 innate lymphoid cells (ILC2) and polyfunctional CD4+ T-helper type 2 (Th2) cells, resulting in the secretion of Th2 anti-inflammatory cytokines (IL-4, IL-5, IL-9, IL-10 and IL-13).4
CD4+ T-cells play an important role in the host defence against HIV.5 Unlike the anti-inflammatory Th2 immune response to helminths, HIV is controlled by a pro-inflammatory T-helper type 1 (Th1) immune response.6 CD4+ T-cell exposure to IL-12 triggers a Th1 response that leads to the secretion of vital anti-viral pro-inflammatory cytokines, including IL-2, tumour necrosis factor (TNF)-a, interferon (INF)-y, macrophage inflammatory protein (MIP)-1 a/1 p, and regulated on activation, normal T-cell expressed and secreted (RANTES).6
The burden, severity and public health implications of helminth and HIV co-infections in South Africa have not been given the attention they warrant. This raises important research questions about the public health implications of helminth co-infection with HIV in terms of pathogenesis and treatment outcomes. Furthermore, although individual helminth and HIV infection-specific immune responses have been the target of extensive investigation, the specific immune mode of protection during co-infection remains unknown. There is no conclusive evidence to confirm whether helminth-induced immunity modulates HIV-specific immune responses or vice versa.7,8 By 2017, a number of studies carried out in southern Africa (Mozambique, South Africa, Zambia, Zimbabwe), West Africa (Nigeria) and East Africa (Ethiopia, Kenya, Uganda, Tanzania) reported contrasting findings, with some indicating the detrimental effects of helminthiasis on HIV immune responses, while others reported no evidence of harmful immunological interactions.9 Many of these studies were cross-sectional and observational in nature, which may have limited the ability of the results to be conclusive. The West and East African regions have the lowest HIV/AIDS burden when compared to the worst affected southern region, which may explain the lack of studies in the West African region. However, surprisingly, the East African region had the highest number of studies when compared to other African regions.9 Epidemiological data support a bidirectional interaction between helminths and HIV; however, it is challenging to distinguish the direction of effects in observational studies, and so in order to reach a definitive conclusion, well-designed, randomised and controlled intervention studies are urgently needed.10
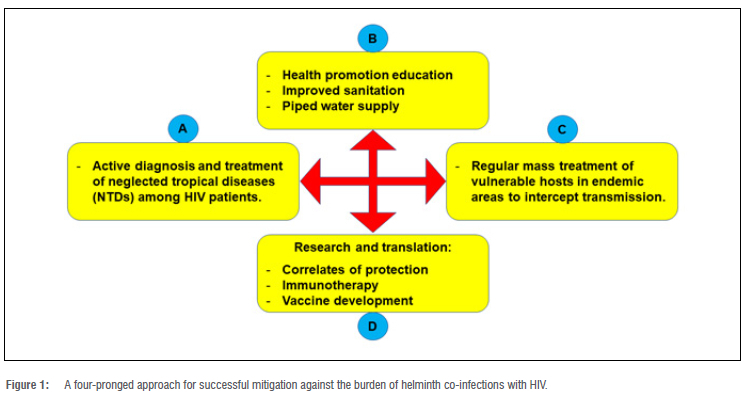
Treatment of helminthic infections is simple, widely available, relatively inexpensive, and has recently become a priority for public health intervention in Africa.6 Albendazole and mebendazole are widely used anti-helminthic drugs.11 Hence, it would be more feasible if more concerted research is done to demonstrate the impact of deworming on HIV/AIDS disease progression. Additionally, a more effective strategy that can be employed to mitigate against the burden of helminth co-infections with HIV is described in the four-pronged approach in Figure 1. The results of such research would point towards identification of correlates of transmission, the immunological responses during co-infection, development of immunotherapeutic interventions, and development of locally relevant vaccines that are geared towards populations that are exposed to helminths.
Helminth and HIV/AIDS co-infection immune responses
Helminth-infected individuals have shown increased expression of co-receptors of HIV-1 chemokines on T-lymphocytes and monocytes.12,13 This is one of the contributing mechanisms in facilitating easy HIV-1 entry and increasing the pool of cells susceptible to infection. Peripheral blood mononuclear cells (PBMCs) obtained from helminth-infected individuals showed increased susceptibility to HIV-1 infection.14,15 Furthermore, a decreased CD8+ cytolytic HIV-1-specific T-cell response and increased IL-10 production were observed in HIV patients co-infected with Schistosoma mansoni compared to those with HIV-1 infection only.16,17 Similarly, reduction of CD4+ cells and increased CD8+ cells were reported among helminth-infected and HIV-1 uninfected individuals, suggesting that helminths on their own reduce the pool of CD4+ cells, which is critical in orchestrating effective immune responses against all pathogens.18 The predominance of Th2 cellular and molecular components resulting from helminth infections downregulates the dominance of the Th1 component, which is essential for the control of HIV infection.19
Another classic feature of helminth infection is marked eosinophilia. Eosinophils have been shown to express CD4+ molecules upon activation and are easily infectable by HIV.20 The increased number of eosinophils in the mucosa (gut, genital and rectal) may facilitate easy acquisition and cell-cell transmission of HIV in helminth-infected hosts.20 In a placebo-controlled trial, Ascaris lumbricoides co-infected HIV-positive patients treated with albendazole showed a significant increase in peripheral CD4+ T-cells and a decreasing trend in HIV-1 RNA levels compared to the placebo group.21 In summary, the above non-exhaustive account of immune dysregulation, Th2 bias and generalised immunosuppression caused by chronic helminth infections describes a favourable environment where HIV can rapidly replicate and spread in and amongst individuals chronically exposed to helminth infections.22
In South Africa, there is a striking paucity of data on the immunological consequences and therapeutic outcome of infection with helminths among HIV/AIDS infected individuals. For example, a South African study reported that dual HIV and helminth infection, together with helminth egg excretion and/or high levels of Ascaris-specific IgE, may be linked to a poor proliferative capacity and deleterious cytokine profile with regard to HIV control.23 In addition, results from the same study population concluded that people with both helminth egg excretion and high anti-Ascaris IgE levels had dysregulated immune cells and high viral loads.23 A modified Th2 helminth response in individuals with eggpositive stools and low levels of anti-Ascaris IgE showed an improved HIV-related immune profile.23 The association between HIV and helminth co-infections and lower biochemical micronutrients, protein and carbohydrate levels in humans has also been shown.24 Additionally, several nutrient deficiencies have been found to increase the likelihood of HIV and helminth co-infection.24 It is understood that various nutrient deficiencies cause impaired immunity.23 These findings illustrate the need for further research that may better describe and explain the effects of helminth immune modulation in individuals co-infected with HIV, leading to successful development of helminth and HIV-related vaccines which will be effective and timely accessible to save lives and improve the general health of those co-infected.
Helminth treatment effect on HIV/AIDS co-infected patients
Treatment recommendations in regions where soil-transmitted helminths and schistosomiasis are co-prevalent include albendazole and praziquantel, respectively.25 Helminths are known to reduce the efficacy of the tuberculosis vaccine, Bacillus Calmette-Guerin, and reduce tumour immunosurveillance activities.14 Effective deworming has been associated with a reduction in HIV viraemia, leading to the slower progression to AIDS, and has been shown to normalise immunological parameters related to immune activation, Th2 immune response, and Treg-induced T-cell hypo-responsiveness.26 The synergy between antiretroviral therapy and albendazole in down-regulating the Th2-biased immune response strongly suggests that deworming can be a safe and effective strategy for improving the health and quality of life of co-infected individuals.26 More interventional research to demonstrate the impact of deworming on HIV/AIDS disease progression is thus required to improve our understanding of the detrimental and beneficial effects of helminth co-infection with HIV.
Knowledge gaps and way forward
The burden and severity of helminth and HIV co-infections have not been given the attention they warrant. Many questions remain unanswered regarding the immunological and pathological consequences of helminth and HIV co-infections. Several human studies investigated the immunological interactions between these infections but results thus far remain inconclusive. This raises important research questions about the public health implications of helminth co-infection with HIV in terms of pathogenesis and treatment outcomes. Furthermore, despite extensive research into individual helminth and HIV infection-specific immune response, the specific immune mode of protection by helminths remains unknown. It is, however, conceivable that the mechanisms involved in both pro-inflammatory and regulatory responses may impact the outcome of HIV/helminth exposure and infection.
There is a real need for interventions that can address the burden of helminth and HIV co-infections. The 95-95-95 targets set by UNAIDS were designed to end the AIDS epidemic by 2030. The three focal targets are that: (1) 95% of those living with HIV must be aware of their status, (2) 95% of those who are aware of their status must be receiving treatment, and (3) 95% of those receiving treatment must have suppressed viral loads. In addition, other targets include achieving zero discrimination and reducing the annual number of new HIV infections among adults to under 200 000 per year. In addition to our proposed four-pronged approach to helminth elimination, similar to the UNAIDS 2030 target goals, the United Nations should also consider having a 95-95-95 target approach to eliminate helminthiasis by 2030. The three proposed focal targets to eliminate helminthiasis should include (1) having 95% of those living with helminth infections be aware of their status, (2) having 95% of those who are aware of their helminth infection status be on anti-helminthic treatment (albendazole, mebendazole and praziquantel) and (3) 95% of those receiving anti-helminthic treatment be educated on good hygiene practices to reduce the cycle of re-infection.
In addition, more transdisciplinary studies are needed to investigate the (1) immunomodulating effects of chronic helminth infection and the impact of such effects on HIV infection, with a focus on the functional and molecular profile of HIV-specific immune responses, (2) association of immunologic markers of helminths and HIV, with control of pathogen replication and disease, (3) impact of helminth co-infections on measures of disease activity for HIV, to promote the understanding of interactions between helminths and HIV, their influence on disease activity, and (4) impact of co-infection on treatment outcomes for HIV.
In conclusion, medical and technological advancements, as well as strengthened multi-pronged interventions, such as (1) early infection detection, (2) mass deworming programmes, (3) continuous surveillance, (4) proper sanitation and clean water provision, (5) development of sensitive diagnostic tools, (6) allocation of more research funds, (7) new therapeutic agents, and (8) prophylactic vaccines, are required for successful helminthiasis elimination.27
Disclaimer
The views and opinions expressed in this article are those of the authors and do not necessarily reflect the official policy or position of any affiliated institutions of the authors.
Acknowledgements
Research reported in this publication was supported by the South African Medical Research Council (SAMRC) through its Division of Research Capacity Development under the Research Capacity Development Initiative from funding received from the South African National Treasury. The content and findings reported / illustrated are the sole deduction, view and responsibility of the researchers and do not reflect the official position and sentiments of the SAMRC.
Competing interests
We have no competing interests to declare.
References
1. Simon GG. Impacts of neglected tropical disease onincidence and progression of HIV/AIDS, tuberculosis, and malaria: Scientific links. Int J Infect Dis. 2016;42:54-57. https://doi.org/10.1016/j.ijid.2015.11.006 [ Links ]
2. World Health Organization. Soil-transmitted helminth infections [webpage on the Internet]. c2020 [cited 2022 Aug 04]. Available from: https://www.who.int/news-room/fact-sheets/detail/soil-transmitted-helminth-infections [ Links ]
3. World Health Organization. Schistosomiasis and soil-transmitted helminthiases: Number of people treated in 2016. Wkly Epidemiol Rec. 2017;49(92):749-760. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19219964%5Cnhttp://orton.catie.ac.cr/cgi-bin/wxis.exe/ [ Links ]
4. Ramalingam TR, Pesce JT, Mentink-Kane MM, Madala S, Cheever AW, Comeau MR, et al. Regulation of helminth-induced Th2 responses by thymic stromal lymphopoietin. J Immunol. 2009;182(10):6452. https://doi.org/10.4049/jimmunol.0900181 [ Links ]
5. Okoye AA, Picker LJ. CD4+ T cell depletion in HIV infection: Mechanisms of immunological failure. Immunol Rev. 2013;254(1):54. https://doi.org/10.1111/imr.12066 [ Links ]
6. Salgame P Yap GS, Gause WC. Effect of helminth-induced immunity on infections with microbial pathogens. Nat Immunol. 2013;14(11):1118-1126. https://doi.org/10.1038/ni.2736 [ Links ]
7. Brown M, Mawa PA, Kaleebu P Elliott AM. Helminths and HIV infection: Epidemiological observations on immunological hypotheses. Parasite Immunol. 2006;28(11):613-623. http://doi.wiley.com/10.1111/j.1365-3024.2006.00904.x [ Links ]
8. Fincham JE, Markus MB, Adams VJ. Could control of soil-transmitted helminthic infection influence the HIV/AIDS pandemic. Acta Trop. 2003;86(2-3):315-333. https://doi.org/10.1016/S0001-706X(03)00063-9 [ Links ]
9. Mkhize-Kwitshana ZL, Tadokera R, Mabaso MHL. Helminthiasis: A systematic review of the immune interactions present in individuals coinfected with HIV and/or tuberculosis. IntechOpen; 2017. https://doi.org/10.5772/66124 [ Links ]
10. Brown M, Mawa PA, Kaleebu P, Elliott AM. Helminths and HIV infection: Epidemiological observations on immunological hypotheses. Parasite Immunol. 2006;28(11):613-623. https://doi.org/10.1111/j.1365-3024.2006.00904 [ Links ]
11. Chai JY, Jung BK, Hong SJ. Albendazole and mebendazole as anti-parasitic and anti-cancer agents: An update. Korean J Parasitol. 2021;59(3):189. https://doi.org/10.3347/kjp.2021.59.3.189 [ Links ]
12. Sundstrom JB, Hair GA, Ansari AA, Secor WE, Gilfillan AM, Metcalfe DD, et al. IgE-FceRI interactions determine HIV coreceptor usage and susceptibility to infection during ontogeny of mast cells. J Immunol. 2009;182(10):6401-6409. https://doi.org/10.4049/jimmunol.0801481 [ Links ]
13. Secor WE, Shah A, Mwinzi PMN, Ndenga BA, Watta CO, Karanja DMS. Increased density of human immunodeficiency virus type 1 coreceptors CCR5 and CXCR4 on the surfaces of CD4+ T cells and monocytes of patients with Schistosoma mansoni infection. Infect Immun. 2003;71(11):6668-6671. https://doi.org/10.1128/IAI.71.11.6668-6671.2003 [ Links ]
14. Elias D, Wolday D, Akuffo H, Petros B, Bronner U, Britton S. Effect of deworming on human T cell responses to mycobacterial antigens in helminth-exposed individuals before and after bacille Calmette-Guerin (BCG) vaccination. Clin Exp Immunol. 2001;123(2):219-225. https://doi.org/10.1046/j.1365-2249.2001.01446.x [ Links ]
15. Borkow G, Weisman Z, Leng Q, Stein M, Kalinkovich A, Wolday D, et al. Helminths, human immunodeficiency virus and tuberculosis. Scand J Infect Dis. 2001;33(8):568-571. https://doi.org/10.1080/00365540110026656 [ Links ]
16. Elrefaei M, Baker CAR, Jones NG, Bangsberg DR, Cao H. Presence of suppressor HIV-specific CD8+ T cells is associated with increased PD-1 expression on effector CD8+ T cells. J Immunol. 2008;180(11):7757-7763. https://doi.org/10.4049/jimmunol.180.11.7757 [ Links ]
17. McElroy MD, Elrefaei M, Jones N, Ssali F, Mugyenyi P, Barugahare B, et al. Coinfection with Schistosoma mansoni is associated with decreased HIV-specific cytolysis and increased IL-10 production. J Immunol. 2005;174(8):5119-5123. https://doi.org/10.4049/jimmunol.174.8.5119 [ Links ]
18. Eggena MP, Barugahare B, Jones N, Okello M, Mutalya S, Kityo C, et al. Depletion of regulatory T cells in HIV infection is associated with immune activation. J Immunol. 2005;174(7):4407-4414. https://doi.org/10.4049/jimmunol.174.7.4407 [ Links ]
19. Borkow G, Bentwich Z. Chronic immune activation associated with chronic helminthic and human immunodeficiency virus infections: Role of hyporesponsiveness and anergy. Clin Microbiol Rev. 2004;17(4):1012-1030. https://doi.org/10.1128/CMR.17.4.1012-1030.2004 [ Links ]
20. Lucey DR, Dorsky DI, Nicholson-Weller A, Weller PF. Human eosinophils express CD4 protein and bind human immunodeficiency virus 1 gp120. J Exp Med. 1989;169(1):327. https://doi.org/10.1084/jem.169.L327 [ Links ]
21. Walson JL, Herrin BR, John-Stewart G. Deworming helminth co-infected individuals for delaying HIV disease progression. Cochrane Database of Systematic Reviews. 2009;3. https://doi.org/10.1002/14651858.CD006419.pub3 [ Links ]
22. Maizels RM, McSorley HJ. Regulation of the host immune system by helminth parasites. J Allergy Clin Immunol. 2016;138(3):666-675. https://doi.org/10.1016/j.jaci.2016.07.007 [ Links ]
23. Mkhize-Kwitshana ZL, Mabaso MLH, Walzl G. Proliferative capacity and cytokine production by cells of HIV-infected and uninfected adults with different helminth infection phenotypes in South Africa. BMC Infect Dis. 2014;14(1):499. https://doi.org/10.1186/1471-2334-14-499 [ Links ]
24. Mkhize BT, Mabaso MHL, Madurai S, Mkhize-Kwitshana ZL. The investigation of the use of prealbumin as a tool for nutritional assessment in adults coinfected with HIV and intestinal helminth parasites in KwaZulu-Natal, South Africa. Biomed Res Int. 2018;2018:1-8. https://doi.org/10.1155/2018/7805857 [ Links ]
25. Fincham JE, Markus MB, Adams VJ. Could control of soil-transmitted helminthic infection influence the HIV/AIDS pandemic. Acta Trop. 2003;86(2-3):315-333. https://doi.org/10.1016/S0001-706X(03)00063-9 [ Links ]
26. Brown M, Mawa PA, Joseph S, Bukusuba J, Watera C, Whitworth JAG, et al. Treatment of Schistosoma mansoni infection increases helminth-specific type 2 cytokine responses and HIV-1 loads in coinfected Ugandan adults. J Infect Dis. 2005;191(10):1648-1657. https://doi.org/10.1086/429668 [ Links ]
27. Karunamoorthi K, Almalki M, Ghailan K. Schistosomiasis: A neglected tropical disease of poverty: A call for intersectoral mitigation strategies for better health. J Health Res Rev. 2018;5(1):1. https://doi.org/10.4103/jhrr.jhrr_92_17 [ Links ]
 Correspondence:
Correspondence:
Miranda Mpaka-Mbatha
Email:mpakamiranda@gmail.com
Funding: South African Medical Research Council
Published: 31 January 2023


{kind=link}