










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.118 no.3-4 Pretoria mar./abr. 2022
http://dx.doi.org/10.17159/sajs.2022/12544
RESEARCH LETTER
Mandibular ramus morphology and species identification in Australopithecus sediba
John HawksI; Lee R. BergerII
IDepartment of Anthropology, University of Wisconsin-Madison, Madison, Wisconsin, USA
IICentre for the Exploration of the Deep Human Journey, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
The site of Malapa, South Africa, has produced fossil evidence from multiple individuals of Australopithecus sediba including the partial skeletons designated as MH1 (holotype) and MH2 (paratype). A recent article in this Journal presented the hypothesis that MH1 and MH2 are not one species but instead represent two different genera: Australopithecus and Homo, respectively. Here we briefly evaluate this claim. We review the evidence from across the skeleton that demonstrates that MH1 and MH2 represent a single species, and we highlight other fossil samples that show the same pattern of mandibular ramus variation as observed in MH1 and MH2. The evidence shows that there is no reason to separate MH1 and MH2 into different species or genera based upon mandibular ramus morphology. This case illustrates how misleading small fragments of anatomy can be, why researchers should not use such fragments particularly for species and genus-level diagnoses, and why it is essential to use all available evidence.
SIGNIFICANCE:
This study shows that the mandibular variation that is present in fossils from Malapa attributed to Australopithecus sediba has parallels in both Australopithecus africanus and in Homo. This helps to demonstrate that mandibular form is not sufficient to provide evidence of species diagnosis, but also that the development and adaptations to diet in Au. sediba were overlapping with those present in other related species of hominins.
Keywords: species, Australopithecus, Malapa, Homo
The MH1 and MH2 skeletons are among the most complete known for Australopithecus, dating to approximately 1.977 million years ago.1,2 The preserved elements of each skeleton include portions of upper and lower limb, thorax, pelvis, mandible, dentition and, for MH1, the face and cranial vault.1,3 These remains are among the most studied of any early hominin specimens. Excavation at Malapa has recovered substantial evidence of the burial position of each skeleton, including joints found in articulation or in close anatomical proximity, with all recovered parts showing a low degree of post-mortem dispersion.4 Additional context comes from the different ontogenetic stages and biological sex of the two skeletons. MH2 is adult and MH1 is juvenile with postcranial and dental elements consistent with a maturational age of between 9 and 11 years when compared to a chimpanzee maturational pattern.5 The pelvic remains of MH1 and MH2 are closely similar in size, and similar in most aspects of morphology, but differ in features related to sex, suggesting female sex for MH2 and male sex for MH1.6,7 The slightly larger size of MH1 in many dental and postcranial measurements is consistent with this sex difference. The metric differences between MH1 and MH2 are consistently slight in comparison to the variation observed within other hominin fossil samples that represent single species, within living humans, and within species of other living great apes. The mandibles of both skeletons preserve most of their mandibular dentition, and the teeth of both individuals are very similar in size and morphology.8
Not only are these two skeletons very similar to each other across all aspects of their anatomy, together they exhibit the same differences from other hominin species. The lateral plantar process of the MH2 calcaneus has a dorsal position not observed in any other fossil hominin except for the immature calcaneal apophysis of MH1.9 Both MH1 and MH2 share a more human-like parasagittal orientation of the ilium, contrasting not only with other Australopithecus pelves but also with pelvic material from Homo erectus and Homo naledi.7Both the MH1 and MH2 proximal femora exhibit superoinferiorly short femoral neck diameters compared to other fossils attributed to Australopithecus.10Both MH1 and MH2 share small molar size and small mandibular corpus area compared to samples of other species of Australopithecus.1,11Both MH1 and MH2 have similar dental microwear textures, and both have near-identical enamel carbon-13 values that are far below any other contemporary sample of Australopithecus, Paranthropus, or Homo.12All of this evidence strongly supports the diagnosis of MH1 and MH2 as belonging to the same species, Au. sediba.
Rak et al.13 claim that MH1 and MH2 belong to different genera. In their assessment, MH2 shares a generalised mandibular ramus form with modern humans, chimpanzees, bonobos, and orangutans, which they infer to be ancestral in hominins. In their description, MH1 shares a derived ramus morphology with Australopithecus afarensis and Paranthropus robustus. In their opinion, these differences are best explained by the hypothesis that MH1 is Australopithecus and MH2 is Homo.
But Rak et al.13 include only 12 fossil mandibles for comparison in addition to MH1 and MH2 in their analysis. Only five of these are used to represent the entire variation exhibited by the genus Homo, while three represent Australopithecus afarensis and three represent Paranthropus robustus. This tiny number omits many well-known mandibular fossils of Australopithecus and Homo. Previous work has shown the extensive variation in ramus form within Homo, including both variation within species and populations, as well as overlap between them.14,15 In this short comment we cannot reiterate this extensive work, but we can bring attention to a few critical omissions.
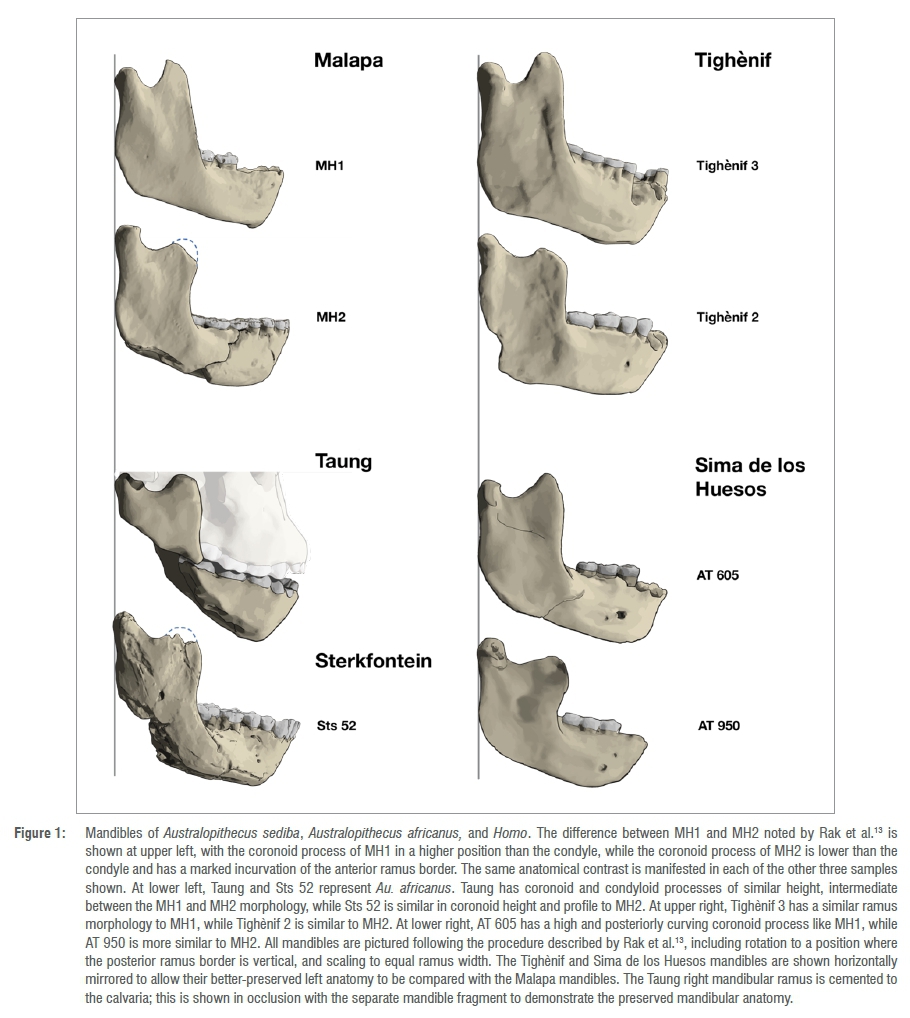
One critical omission is the geographically contiguous species Australopithecus africanus. Rak et al.13 claim that a fragment of the Sts 7 mandible embedded in breccia appears to resemble the MH1 morphology, but otherwise do not comment on this key species. We illustrate two additional mandibles in Figure 1. The holotype, from Taung, preserves the right ramus cemented to the calvaria in anatomical position. The coronoid process is of similar height to the condyle and the form of the mandibular notch is similar to that of SK 63, a mandible of similar ontogenetic age attributed to P. robustus. This is intermediate in morphology between MH1 and MH2 (Figure 1). The Sterkfontein Member 4 hominin Sts 52 has a well-preserved right mandibular ramus, with slight damage to the superior margin of the coronoid process. The outline of the preserved mandibular notch and anterior ramus border are very similar to MH2 (Figure 1). The break to the coronoid process is likewise similar, and a reasonable reconstruction yields a similar coronoid height in Sts 52 as in MH2. The Taung fossil is thought to date to between 3.0 and 2.6 million years old while Sts 52 dates to between 2.6 and 2.0 million years ago.16
Another critical omission is the sample of well-known mandibular ramus fossils attributed to Homo. In Figure 1, we picture four mandibles of Homo that were not examined by Rak et al.13 One pair includes Tighènif 2 and 3 from Tighènif, Algeria17,18, thought to date to the later Early Pleistocene between 1.4 million and 900 000 years ago19,20. The other pair includes AT-950 and AT-605 from Sima de los Huesos, Spain, which date to approximately 430 000 years ago.21-23 Both pairs exhibit the same pattern of mandibular ramus variation as the two Malapa mandibles. Tighènif 3 and AT-605 both have tall, hook-shaped coronoid processes that extend higher than the condyle, posteriorly positioned mandibular notches, and anterior ramus margins that ascend smoothly with no incurvation - all features found in MH1 that Rak et al.13 suggest are diagnostic of Australopithecus. In contrast, Tighènif 2 and AT-950 share the anatomy seen in MH2, with low coronoid height, mandibular notch position near the midpoint of the ramus, and an anterior ramus profile with an incurving base. These are the features of MH2 that Rak et al.13 suggest are diagnostic of Homo.
Rak et al.13 claim to show that 'the differences [between MH1 and MH2] are beyond what is expected in a trait's normal range of distribution in a given population' (p. 2). The three other samples that we show in Figure 1 disprove this assertion. Each sample has the mandibular ramus variation manifested at Malapa. The morphological features that Rak et al.13 describe as derived exclusively in Australopithecus actually occur widely across Homo. The form that Rak et al.13 describe as characteristic of Homo occurs not only within Au. sediba, but also within Au. africanus. The observation that hominin samples vary in these features is consistent with the results of Ritzman et al.24, who found that the morphological distance of mandibular ramus form between MH1 and MH2 is no greater than between randomly chosen pairs within modern humans, Pan, and Gorilla. The widespread variation of these ramus features makes them unsuitable to be used for taxonomic diagnosis on their own.
Palaeoanthropologists have grappled with the boundary between Homo and Australopithecus for more than 70 years. The fragmentary condition of fossils attributed to 'early Homo' is one reason for a lack of consensus about the definition of the genus. Samples that preserve more complete skeletal material, including Malapa, demonstrate that features usually found in later Homo do not assort with each other in the ways that researchers once predicted based upon fragmentary remains. Human evolution was not tidy. Hominin skeletons including MH1 and MH2 are anatomical mosaics that do not always fit a simple evolutionary tree. We are far from alone in recognising the homoplasy in the hominin fossil record and the way that it can confuse phylogenetic placement of fossils.25,26 The field must meet these challenges by considering all relevant data. A small sample cannot be better evidence than a large sample. A small fragment of anatomy cannot be better evidence than a skeleton.
Competing interests
We have no competing interests to declare.
Authors' contributions
Both authors conceived of the study, contributed data, wrote the manuscript, and edited the manuscript.
References
1. Berger LR, De Ruiter DJ, Churchill SE, Schmid P Carlson KJ, Dirks PH, et al. Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science. 2010;328(5975):195-204. https://doi.org/10.1126/science.1184944 [ Links ]
2. Pickering R, Dirks PH, Jinnah Z, De Ruiter DJ, Churchill SE, Herries AI, et al. Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo. Science. 2011;333(6048):1421-1423. https://doi.org/10.1126/science.1203697 [ Links ]
3. Williams SA, DeSilva JM, De Ruiter DJ. Australopithecus sediba Malapa at 10: Introduction to the Special Issue on Australopithecus sediba. PaleoAnthropology. 2018:49-55. [ Links ]
4. Val A, Dirks PH, Backwell LR, d'Errico F, Berger LR. Taphonomic analysis of the faunal assemblage associated with the hominins (Australopithecus sediba) from the Early Pleistocene cave deposits of Malapa, South Africa. PLoS ONE. 2015;10(6), e0126904. https://doi.org/10.1371/journal.pone.0126904 [ Links ]
5. Cameron N, Bogin B, Bolter D, Berger LR. The postcranial skeletal maturation of Australopithecus sediba. Am J Phys Anthropol. 2017;163(3):633-640. https://doi.org/10.1002/ajpa.23234 [ Links ]
6. Kibii JM, Churchill SE, Schmid P Carlson KJ, Reed ND, De Ruiter DJ, et al. A partial pelvis of Australopithecus sediba. Science. 2011;333(6048):1407-1411. https://doi.org/10.1126/science.1202521 [ Links ]
7. Churchill SE, Kibii J, Schmid P Reed ND, Berger LR. The pelvis of Australopithecus sediba. PaleoAnthropology. 2018;2018:334-356. [ Links ]
8. Irish JD, Guatelli-Steinberg D, Legge SS, De Ruiter DJ, Berger LR. Dental morphology and the phylogenetic "place" of Australopithecus sediba. Science. 2013;340(6129), Art. #1233062. https://doi.org/10.1126/science.1233062 [ Links ]
9. Zipfel B, DeSilva JM, Kidd RS, Carlson KJ, Churchill SE, Berger LR. The foot and ankle of Australopithecus sediba. Science. 2011;333(6048):1417-1420. https://doi.org/10.1126/science.1202703 [ Links ]
10. DeSilva JM, Carlson KJ, Claxton AG, Harcourt-Smith WE, McNutt EJ, Sylvester AD, et al. The anatomy of the lower limb skeleton of Australopithecus sediba. PaleoAnthropology. 2018;2018:357-105. [ Links ]
11. De Ruiter DJ, DeWitt TJ, Carlson KB, Brophy JK, Schroeder L, Ackermann RR, et al. Mandibular remains support taxonomic validity of Australopithecus sediba. Science. 2013;340(6129), Art. #1232997. https://doi.org/10.1126/science.1232997 [ Links ]
12. Henry AG, Ungar PS, Passey BH, Sponheimer M, Rossouw L, Bamford M, et al. The diet of Australopithecus sediba. Nature. 2012;487(7405):90-93. https://doi.org/10.1038/nature11185 [ Links ]
13. Rak Y Geffen E, Hylander W, Ginzburg A, Been E. One hominin taxon or two at Malapa Cave? Implications for the origins of Homo. S Afr J Sci. 2021;117(5-6), Art. #8747. https://doi.org/10.17159/sajs.2021/8747 [ Links ]
14. Wolpoff MH, Frayer DW. Unique ramus anatomy for Neandertals? Am J Phys Anthropol. 2005;128(2):245-251. https://doi.org/10.1002/ajpa.10432 [ Links ]
15. Terhune CE, Robinson CA, Ritzman TB. Ontogenetic variation in the mandibular ramus of great apes and humans. J Morphol. 2014;275(6):661-677. https://doi.org/10.1002/jmor.20246 [ Links ]
16. Herries AI, Pickering R, Adams JW, Curnoe D, Warr G, Latham AG, et al. A multi-disciplinary perspective on the age of Australopithecus in southern Africa. In: Reed K, Fleagle J, Leakey RF, editors. The paleobiology of Australopithecus. Dordrecht: Springer; 2013. p. 21-40. https://doi.org/10.1007/978-94-007-5919-0_3 [ Links ]
17. Arambourg C. Le gisement de Ternifine et l'Atlanthropus [The Ternifine deposit and the Atlanthropus]. Bulletin de la Société préhistorique de France. 1955;52(Fasc. 1/2):94-95. French. https://doi.org/10.3406/bspf.1955.3159 [ Links ]
18. Arambourg C. Le gisement de Ternifine, t. 1. Deuxième partie: l'Atlanthropus mauritanien [The Ternifine deposit, t. 1. Second part: Atlanthropus mauritanicus]. Archives de l'Institut de Paléontologie humaine, Mémoire. 1963;1963(32):37-141. French. [ Links ]
19. Geraads D. Pleistocene Carnivora (Mammalia) from Tighennif (Ternifine), Algeria. Geobios. 2016;49(6):445-458. https://doi.org/10.1016/j.geobios.2016.09.001 [ Links ]
20. Pickford M. The fossil Suidae (Mammalia, Artiodactyla) from Ternifine (Tighènif) Algeria. Münchner Geowissenschafltiche Abhandlungen. 2020;50:1-67. [ Links ]
21. Rosas A. Seventeen new mandibular specimens from the Atapuerca/ Ibeas Middle Pleistocene Hominids sample (1985-1992). J Hum Evol. 1995;28(6):533-559. https://doi.org/10.1006/jhev.1995.1041 [ Links ]
22. Rosas A, Bastir M, Alarcón JA. Tempo and mode in the Neandertal evolutionary lineage: A structuralist approach to mandible variation. Quat Sci Rev. 2019;217:62-75. https://doi.org/10.1016/j.quascirev.2019.02.025 [ Links ]
23. Demuro M, Arnold LJ, Aranburu A, Sala N, Arsuaga JL. New bracketing luminescence ages constrain the Sima de los Huesos hominin fossils (Atapuerca, Spain) to MIS 12. J Hum Evol. 2019;131:76-95. https://doi.org/10.1016/j.jhevol.2018.12.003 [ Links ]
24. Ritzman TB, Terhune CE, Gunz P, Robinson CA. Mandibular ramus shape of Australopithecus sediba suggests a single variable species. J Hum Evol. 2016;100:54-64. https://doi.org/10.1016/j.jhevol.2016.09.002 [ Links ]
25. Wood B, Harrison T. The evolutionary context of the first hominins. Nature. 2011;470(7334):347-352. https://doi.org/10.1038/nature09709 [ Links ]
26. Dembo M, Radovcic D, Garvin HM, Laird MF, Schroeder L, Scott JE, et al. The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods. J Hum Evol. 2016;97:17-26. https://doi.org/10.1016/jJjhevoL2016.04.008 [ Links ]
 Correspondence:
Correspondence:
John Hawks
Email: jhawks@wisc.edu
Received: 13 Oct. 2021
Revised: 14 Jan. 2022
Accepted: 14 Jan. 2022
Published: 29 Mar. 2022
EDITORS: Margaret Avery Jemma Finch
FUNDING: None


{kind=link}