










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.11-12 Pretoria nov./dic. 2021
http://dx.doi.org/10.17159/sajs.2021/10409
RESEARCH ARTICLE
Mechanical loading of primate fingers on vertical rock surfaces
Michael C. EverettI; Marina C. ElliottII, III; David GaynorIV; Austin C. HillI, V; Samar M. SyedaVI; Jesse CasanaI; Bernhard ZipfelVII; Jeremy M. DeSilvaI, VII; Nathaniel J. DominyI
IDepartment of Anthropology, Dartmouth College, Hanover, New Hampshire, USA
IIDepartment of Archaeology, Simon Fraser University, Burnaby, British Columbia, Canada
IIICentre for the Exploration of the Deep Human Journey, University of the Witwatersrand, Johannesburg, South Africa
IVMammal Research Institute, University of Pretoria, Pretoria, South Africa
VDepartment of Anthropology University of Pennsylvania, Philadelphia, Pennsylvania, USA
VISkeletal Biology Research Centre, School of Anthropology and Conservation, University of Kent, Canterbury, UK
VIIEvolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Mechanical loading of finger bones (phalanges) can induce angular curvature, which benefits arboreal primates by dissipating forces and economising the recruitment of muscles during climbing. The recent discovery of extremely curved phalanges in a hominin, Homo naledi, is puzzling, for it suggests life in an arboreal milieu, or, alternatively, habitual climbing on vertical rock surfaces. The importance of climbing rock walls is attested by several populations of baboons, one of which uses a 7-m vertical surface to enter and exit Dronkvlei Cave, De Hoop Nature Reserve, South Africa. This rock surface is an attractive model for estimating the probability of extreme mechanical loading on the phalanges of rock-climbing primates. Here we use three-dimensional photogrammetry to show that 82-91% of the climbable surface would generate high forces on the flexor tendon pulley system and severely load the phalanges of baboons and H. naledi. If such proportions are representative of vertical rock surfaces elsewhere, it may be sufficient to induce stress-mitigating curvature in the phalanges of primates.
SIGNIFICANCE:
• We present the first three-dimensional photogrammetric analysis of a vertical rock surface climbed by a non-human primate, the chacma baboon (Papio ursinus).
• Our results show that a large proportion of a vertical rock wall would compel crimp and slope hand positions during climbing - grips that could explain the extraordinary phalangeal curvature expressed by a Middle Pleistocene hominin, Homo naledi.
Keywords: human evolution, locomotion, climbing, suspension, bone curvature
Introduction
Bone is a dynamic tissue, and repeated mechanical loading can induce changes to its density and angular curvature. For example, loading stresses from vertical climbing and suspensory locomotion can increase the curvature of primate finger bones (phalanges) during growth and development.1-3 Greater curvature is advantageous to arboreal primates because it dissipates forces and dampens recruitment of muscles during finger flexion, thus economising energetic costs and simultaneously lowering the risk of falling. Phalangeal curvature is therefore widely viewed as a measure of arboreality among primates - it is a classic form-functional trait that informs our interpretations of behaviour and ecology in the fossil record1-7 (but see Wallace et al.8 for a counterexample).
Upsetting this orthodoxy is the hand of Homo naledi, recovered from Rising Star Cave, South Africa, in 2013-14 and described in 2015.9 The degree of phalangeal curvature is astounding, with an included angle (0) that far exceeds that of modern humans (Figure 1). Such curvature would normally and unequivocally suggest an arboreal milieu3, but there are at least two reasons to doubt such an inference for H. naledi. First, the phalanges are relatively short, resulting in humanlike hand proportions.9 No arboreal primate has phalanges that are both short and curved; it is an aberration that confounds conventional interpretation. Second, existing specimens of H. naledi are dated to the Middle Pleistocene between 335 kya and 241 kya.12 Nearby faunal assemblages of comparable age - those of Gladysvale13 and Lincoln Cave, Sterkfontein14 - favour habitat conditions that resemble those of today, i.e. a mix of acacia woodland and grassland, neither of which would incentivise a strongly arboreal ecology.15

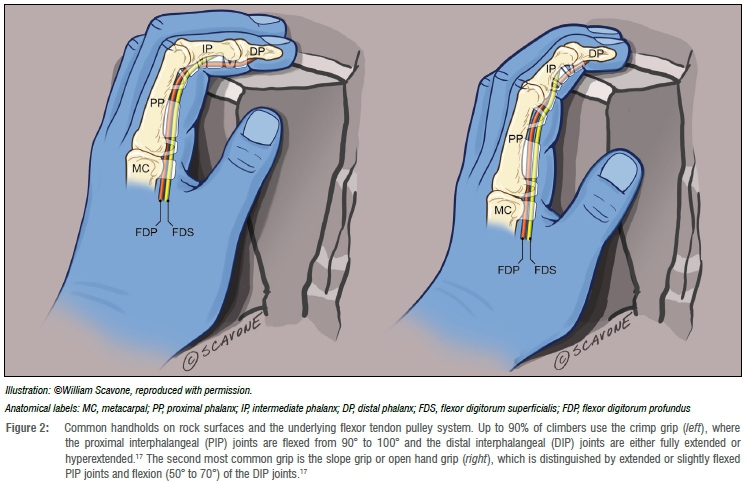
So, what was H. naledi doing with its hands? Voisin et al.16 examined the shoulder girdle of H. naledi and reported morphological traits associated with vertical climbing and suspension. But given the inferred habitat conditions, they discounted arboreal activities and argued instead for 'movement across and climbing on rocky walls'16(p.2). Voisin et al.'s hypothesis is intriguing given that rock climbing puts high forces on finger flexor tendons, especially during the 'crimp' position (Figure 2).17-20 This position puts extreme stress on the flexor tendon pulley system, a series of five annular (A1-A5) and three cruciate ligaments (C1-C3) that resist bowstringing of the tendons. In other words, the pulley system holds the tendons close to the bone, effectively converting linear force into torque that produces flexion at the metacarpophalangeal and interphalangeal joints (Supplementary figure 1).
Ruptures of A217 and other severe pulley injuries are common among human rock climbers21, and a testament to the mechanical loading of phalanges. Indeed, frequent rock climbing is known to cause geometric and cortical thickening of the phalanges22 and it may induce greater curvature, at least hypothetically. Tan et al.18 calculated the benefits of greater phalangeal curvature for reducing tendon and pulley stresses of fingers in the crimp position; however, affirming increased curvature among rock climbers is difficult because it is challenging to measure angles from radiographs of living subjects. Another model system is needed.
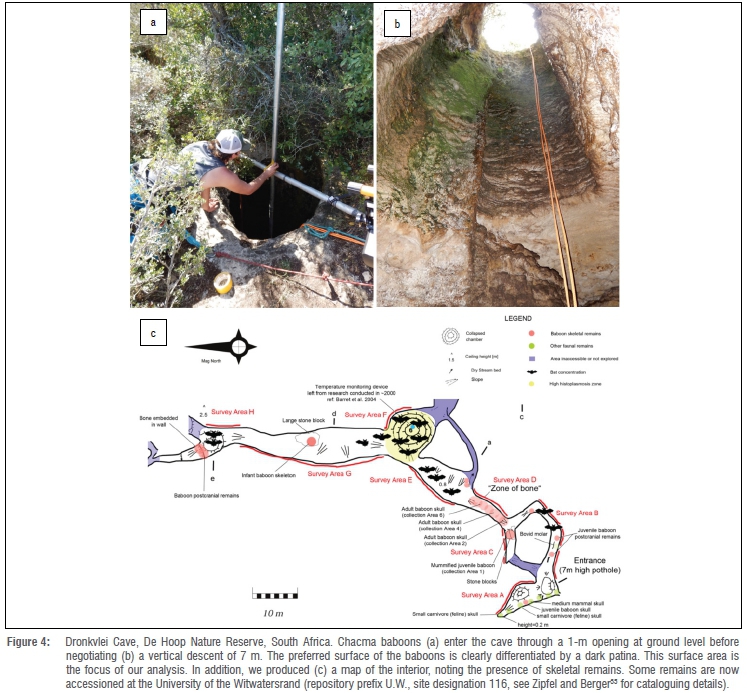
Some populations of baboons and chimpanzees enter cave systems to avoid predators and/or regulate body temperature.23-34 Some caves are only accessible by climbing sheer cliffs.28 For example, Marais23 described a group of chacma baboons (Papio ursinus) and their daily ascent to a cliffside cave opening =150 m high. His vivid account speaks to the mechanical loading of fingers: 'they had to go more than a mile hanging only by their fingers to the [cliff] ledge; their hind feet against the smooth surface'23(p.63). In some cases, baboons must negotiate vertical rock surfaces within a cave. For example, at Misgrot Cave in Thabazimbi, South Africa, baboons face a perilous 17-m vertical descent from ground level (Figure 3)32, whereas those entering Dronkvlei Cave, De Hoop Nature Reserve, South Africa, descend 7 m to access the interior (Figure 4). Baboons use both caves regularly as overnight sleeping sites; Dronkvlei is used on 28% of nights.30
Study aims and design
Dronkvlei Cave is an appealing model system for exploring the topography of a vertical rock surface and its potential to load primate fingers. Here we ask a basic question: what proportion of a vertical rock wall would, when climbed, force the distal phalanges of P. ursinus (and, hypothetically, H. naledi) into a crimp or slope grip position? It is a thought experiment that leaves formal comparative measures of phalangeal curvature in P. ursinus as a priority for future research. To answer our question, we used three-dimensional (3D) photogrammetry to detect and quantify concavities in the surface that range from a minimum graspable depth (determined as the length of the third distal phalanx) to a minimum crimpable depth (determined as the sum length of the third intermediate and distal phalanx) - i.e. depths of 11-25 mm for P. ursinus and 14-37 mm for H. naledi (Table 1).

Methods
Data acquisition
Photogrammetry is a low-cost method for extracting accurate measurements from photographs. It is well suited for producing 3D models of irregular surfaces, such as cave walls.36 To obtain photographs of the rock surface climbed by baboons, we used metal irrigation piping to construct a temporary T-shaped scaffold. The cross pipe was mounted at the cave opening and fixed to a vertical pipe that extended to the shaft floor (Figure 4a), where it was weighted to minimise lateral excursions. We hitched a digital camera to this pipe so that photography occurred from a fixed central axis, and we mounted ground control points to the rock surface. We abseiled into the cave to operate the camera, aiming for 80% overlap between serial photos in the horizontal and vertical planes.
Photogrammetry
We processed the image set (n=354) with Agisoft Metashape Pro (formerly Photoscan Pro), a common package used in archaeological research.37-39 Due to logistical constraints in the field, we did not record spatial data for each ground control point, so the model was scaled to real-world dimensions by using the targets as scale bars, and manually oriented. Manual orientation is suboptimal for any calculations that rely on the slope of the wall, but it should not affect the local depth calculations described below. After alignment, dense cloud construction, and texture application, we trimmed the 3D model to the area used for climbing (Figure 4b) and exported it as an orthophoto (Figure 5a) and digital elevation model (DEM) with an arbitrary local coordinate system. We brought the resulting geotiffs, both with sub-millimetre resolution, into ArcGIS and SAGA GIS for further processing and visualisation.
Depth processing
In ArcGIS, the DEM raster treats the surface of the rock wall as vertical elevation data. To calculate the local relative depth of each raster cell in the rock wall, we used focal statistics with a small neighborhood, and maximum elevation as the statistics type, to create a raster of the local maximum height of the wall, and then smoothed that raster again with focal statistics over a wider neighborhood, creating a smoothed idealised model of the maximum height of the local surface of the cave. We subtracted the original DEM from this idealised surface to find the local difference between each cell and the average nearest 'maximum' value. Finally, we queried this raster of local depths for cells in two ranges (11-25 mm for P ursinus and 14-37 mm for H. naledi) and used this value to calculate the total area of each range.
Results and discussion
The entrance to Dronkvlei Cave is a useful model for exploring potential stresses on the flexor tendon pulley system and phalanges of primates. We focused our analysis on the surface area climbed by baboons -representing 5.98 m2 (Figure 5a) - and we show that crimp- and slope-inducing handholds are distributed relatively evenly across a nearly vertical plane (Figure 5b). However, the total area of such holds differed between the two species.
For P. ursinus, we calculated an area of 0.494 m2 (2 514 603 cells of 0.44 mm x 0.44 mm), and for H. naledi an area of 0.353 m2 (1 795 106 cells of 0.44 mm x 0.44 mm), with an overlap of 0.282 m2 (Figure 3c). Thus, 6-8% of the total surface area, when gripped, is expected to generate high forces on the flexor tendon pulley system and severely load the phalanges of both primate species. Such an estimate is conservative, however, as a large proportion of the rock surface is smooth and unclimbable. When we subtract surface depths of <11 mm and <14 mm from the total, the total graspable surface area is reduced to 0.602 m2 and 0.390 m2, respectively, meaning that as much as 82% (P ursinus) or 91% (H. naledi) of usable handholds would produce forces that would favour greater phalangeal curvature.
The significance of our finding is a matter of speculation - even at 8% of the total surface area, the rock wall of Dronkvlei Cave is a non-trivial source of phalangeal loading when extrapolated over a lifetime. The average life expectancy of a female baboon that survives to adulthood is 12.1 years in Amboseli, Kenya, and 19.7 years in Gombe, Tanzania (with record longevities of 27 years at both sites).40 If we use these parameters to estimate an average life expectancy of =16 years for P. ursinus, and given that animals climb 12 m to enter and exit Dronkvlei Cave on 28% of days30, an individual would encounter =1.6 km of crimp- and slope-grip-inducing surfaces over its lifetime [(16 years x 365 days x 0.28) X (12 m x 0.08 m)]. It is a crude estimate of mechanical 'loading' on the flexor tendon pulley system and distal phalanges, but it invites an analysis of phalangeal curvature in this population.
More instructive perhaps are the mummified/skeletal remains of baboons in Misgrot Cave.34 These individuals climb greater distances than those at Dronkvlei (Figure 3b), but the frequency of overnight use is unknown. Still, each climbing bout carries great risk. Falling from heights as high as 17 m is likely to cause severe morbidity or outright mortality15, although the rugosity of the dolomitic surface may differ from the limestone surface of Dronkvlei Cave in crucial ways, affecting risk. Misgrot Cave is a promising site for studying the natural history of why, and the mechanics of how, baboons climb vertical rock surfaces, as well as the effects on their finger bones. Another promising study system lies in the lava tubes of Mount Suswa, Kenya, the site of an oft-cited analysis by Simons24, who described the skeletal remains of baboons (P. anubis) and leopards, sometimes commingled, at the base of vertical shafts. Remains of baboons continue to accumulate there, serving as a tourist attraction.
So, did H. naledi climb vertical rock surfaces regularly? Answering this question is beyond reach at present, but their bodies, alive or dead, negotiated at least two steep walls of dolomite to arrive in the Dinaledi Chamber of Rising Star Cave: an entry point through the roof of Postbox Chamber and a 12-m fissure ('The Chute').12 Our results suggest that climbing these surfaces would have compelled the use of crimp or slope grips to some extent, and Tan et al.'s18 model of flexor tendon pulley forces suggests that H. naledi would have enjoyed a 23% reduction in phalangeal forces compared to modern humans (Supplementary figure 2). Such findings indicate that high phalangeal curvature - whether mechanically induced or genetically mediated8 - would have conferred advantages to H. naledi during rock climbing.
A limitation of our model system approach is that H. naledi (weighing ~40 kg41) was heavier than P. ursinus is now and probably longer-lived, factors that would load its fingers to a greater extent. Yet the pedal phalanges of H. naledi express the same curvature as those of Papio, which is '[possibly] indicative of elevated pedal grasping ability'42(p.4). Thus, both species are equipped to grip vertical rock surfaces with their feet - a prudent means of distributing body mass and mitigating forces on the fingers during climbing.
We conclude by acknowledging that rock-climbing baboons are an imperfect model for interpreting the curious anatomy of H. naledi, but we would argue that detailed studies of their functional anatomy and climbing kinematics, together with systematic measures of the rock surfaces used, are promising areas of future research.
Acknowledgements
We are grateful to Sergio Almécija, Louise Barrett, Lee-Anne Benjamin, Jackson Dolphin, Elli Goudzwaard, Caitlyn Hauke, Jason Heaton, Peter Henzi, Kudakwashe Jakata, Ashley Kehoe, Tracy Kivell, Scott McGraw, Maureen O'Leary, Randall Susman, and Mary Wilde for generous practical assistance. For access to comparative baboon remains, our thanks go to Brendon Billings, School of Anatomical Sciences, University of the Witwatersrand. This research was approved by CapeNature (permit no. CN44-30-184) and funded by the Claire Garber Goodman Fund, Department of Anthropology, Dartmouth College (to M.C.E) and the Stamps Scholars Program, Dartmouth College (to M.C.E.).
Competing interests
We have no competing interests to declare.
Authors' contributions
M.C.Everett: Conceptualisation, methodology, data collection, data curation, data analysis, writing - the initial draft, writing - revisions, project leadership. M.C.Elliott: Methodology, data collection, writing - revisions. D.G.: Methodology, data collection, writing - revisions. A.C.H.: Methodology, data analysis, writing - the initial draft. S.M.S.: Data collection. J.C.: Methodology, writing - the initial draft. B.Z.: Data curation, data collection, writing - revisions. J.M.D.: Conceptualisation, student supervision, writing - the initial draft, writing - revisions. N.J.D.: Conceptualisation, student supervision, writing - the initial draft, writing - revisions.
References
1. Richmond BG. Biomechanics of phalangeal curvature. J Hum Evol. 2007;53:678-690. http://doi.org/10.1016/j.jhevol.2007.05.011 [ Links ]
2. Congdon KA. Interspecific and ontogenetic variation in proximal pedal phalangeal curvature of great apes (Gorilla gorilla, Pan troglodytes, and Pongopygmaeus). Int J Primatol. 2012;33:418-427. http://doi.org/10.1007/s10764-012-9590-7 [ Links ]
3. Patel BA, Maiolino SA. Morphological diversity in the digital rays of primate hands. In: Kivell TL, Lemelin P Richmond BG, Schmitt D, editors. The evolution of the primate hand: Anatomical, developmental, functional, and paleontological evidence. New York: Springer; 2016. p. 55-100. [ Links ]
4. Susman RL. Comparative and functional morphology of hominoid fingers. Am J Phys Anthropol. 1979;50:215-236. http://doi.org/10.1002/ajpa.1330500211 [ Links ]
5. Jungers WL, Godfrey LR, Simons EL, Chatrath PS. Phalangeal curvature and positional behavior in extinct sloth lemurs (Primates, Palaeopropithecidae). Proc Natl Acad Sci USA. 1997;94:11998-12001. http://doi.org/10.1073/pnas.94.22.11998 [ Links ]
6. Jungers WL, Godfrey LR, Simons EL, Wunderlich RE, Richmond BG, Chatrath PS. Ecomorphology and behavior of giant extinct lemurs from Madagascar. In: Plavcan JM, Kay RF, Jungers WL, Van Schaik CP editors. Reconstructing behavior in the primate fossil record. Boston, MA: Springer; 2002. p. 371411. http://doi.org/10.1007/978-1-4615-1343-8_10 [ Links ]
7. Prang TC, Ramirez K, Grabowski M, Williams SA. Ardipithecus hand provides evidence that humans and chimpanzees evolved from an ancestor with suspensory adaptations. Sci Adv. 2021;7:eabf2474. http://doi.org/10.1126/sciadv.abf2474 [ Links ]
8. Wallace IJ, Burgess ML, Patel BA. Phalangeal curvature in a chimpanzee raised like a human: Implications for inferring arboreality in fossil hominins. Proc Natl Acad Sci USA. 2020;117:11223-11225. http://doi.org/10.1073/pnas.2004371117 [ Links ]
9. Kivell TL, Deane AS, Tocheri MW, Orr CM, Schmid P Hawks J, et al. The hand of Homo naledi. Nat Commun. 2015;6:8431. http://doi.org/10.1038/ncomms9431 [ Links ]
10. Stern JT Jr, Jungers WL, Susman RL. Quantifying phalangeal curvature: An empirical comparison of alternative methods. Am J Phys Anthropol. 1995;97:1-10. http://doi.org/10.1002/ajpa.1330970102 [ Links ]
11. Domínguez-Rodrigo M, Pickering TR, Almécija S, Heaton JL, Baquedano E, Mabulla A, et al. Earliest modern human-like hand bone from a new >1.84-million-year-old site at Olduvai in Tanzania. Nat Commun. 2015;6:7987. http://doi.org/10.1038/ncomms8987 [ Links ]
12. Robbins JL, Dirks PHGM, Roberts EM, Kramers JD, Makhubela TV Hilbert-Wolf HL, et al. Providing context to the Homo naledi fossils: Constraints from flowstones on the age of sediment deposits in rising Star Cave, South Africa. Chem Geol. 2021;567, Art. #120108. http://doi.org/10.1016/j.chemgeo.2021.120108 [ Links ]
13. Lacruz, RS, Brink JS, Hancox PJ, Skinner AR, Herries A, Schmid P, et al. Palaeontology and geological context of a Middle Pleistocene faunal assemblage from the Gladysvale Cave, South Africa. Palaeontol Afr. 2002;38:99-114. http://hdl.handle.net/10539/16351 [ Links ]
14. Reynolds SC, Clarke RJ, Kuman KA. The view from the Lincoln Cave: Mid- to late Pleistocene fossil deposits from Sterkfontein hominid site, South Africa. J Hum Evol. 2007;53:260-271. http://doi.org/10.1016/j.jhevol.2007.02.004 [ Links ]
15. Kraft TS, Venkataraman VV Dominy NJ. A natural history of human tree climbing. J Hum Evol. 2014;71:105-118. http://doi.org/10.1016/j.jhevol.2014.02.002 [ Links ]
16. Voisin JL, Feuerriegel EM, Churchill SE, Berger LR. The Homo naledi shoulder girdle: An adaptation to boulder climbing. LAnthropologie 2020;124:102783. http://doi.org/10.1016/j.anthro.2020.102783 [ Links ]
17. Schweizer A. Biomechanical properties of the crimp grip position in rock climbers. J Biomech. 2001;34:217-223. http://doi.org/10.1016/S0021-9290(00)00184-6 [ Links ]
18. Tan MA, Fuss FK, Niegl G. Stress distribution at the finger pulleys during sport climbing. In: Fuss FK, Subic A, Ujihashi S, editors. The impact of technology on sport II. London: CRC Press; 2007. p. 663-669. https://doi.org/10.1201/9781439828427.ch97 [ Links ]
19. Schöffl I, Oppelt K, Jüngert J, Schweizer A, Neuhuber W, Schöffl V The influence of the crimp and slope grip position on the finger pulley system. J Biomech. 2009;42:2183-2187. http://doi.org/10.1016/j.jbiomech.2009.04.049 [ Links ]
20. Amca AM, Vigouroux L, Aritan S, Berton E. Effect of hold depth and grip technique on maximal finger forces in rock climbing. J Sports Sci. 2012;30:669-677. http://doi.org/10.1080/02640414.2012.658845 [ Links ]
21. Schöffl V Schöffl I, Frank L, Küpper T, Simon M, Lutter C. Tendon injuries in the hands in rock climbers: epidemiology, anatomy, biomechanics and treatment - an update. Muscles Ligaments Tendons J. 2020;10:233-243. http://doi.org/10.32098/mltj.02.2020.08 [ Links ]
22. Morrison AB, Schöffl VR. Physiological responses to rock climbing in young climbers. Br J Sports Med. 2007;41:852-861. https://doi.org/10.1136/bjsm.2007.034827 [ Links ]
23. Marais EN. My friends the baboons. London: Methuen; 1939. [ Links ]
24. Simons JW. The presence of leopard and a study of the food debris in the leopard lairs of the Mount Suswa caves, Kenya. Bull Cave Exploration Group E Afr. 1966;1:51-69. [ Links ]
25. Gow CE. Habitual sheltering in an extensive cave system by baboons near Bredasdorp, South Africa. S Afr J Sci. 1973;69:182. [ Links ]
26. Wells L. Baboons sheltering in caves. S Afr J Sci. 1973;69:279. [ Links ]
27. Brain CK. The hunters or the hunted? An introduction to African cave taphonomy. Chicago, IL: University of Chicago Press; 1981. [ Links ]
28. Hamilton WJ. Baboon sleeping site preferences and relationships to primate grouping patterns. Am J Primatol. 1982;3:41-53. http://doi.org/10.1002/ajp.1350030104 [ Links ]
29. McGrew WC, McKee JK, Tutin CEG. Primates in caves: Two new reports of Papio spp. J Hum Evol. 2003;44:521-526. http://doi.org/10.1016/S0047-2484(03)00042-3 [ Links ]
30. Barrett L, Gaynor D, Rendall D, Mitchell D, Henzi SP Habitual cave use and thermoregulation in chacma baboons (Papio hamadryas ursinus). J Hum Evol. 2004;46:215-222. http://doi.org/10.1016/j.jhevol.2003.11.005 [ Links ]
31. Hodgins G, Brook GA, Marais E. Bomb-spike dating of a mummified baboon in Ludwig Cave, Namibia. Int J Speleol. 2007;36:31-38. http://scholarcommons.usf.edu/ijs/vol36/iss1/3 [ Links ]
32. Egeland CP Domínguez-Rodrigo M, Pickering TR, Menter CG, Heaton JL. Hominin skeletal part abundances and claims of deliberate disposal of corpses in the Middle Pleistocene. Proc Natl Acad Sci USA. 2018;115:4601-4606. http://doi.org/10.1073/pnas.1718678115 [ Links ]
33. Boyer Ontl K, Pruetz JD. Mothers frequent caves: Lactation affects chimpanzee (Pan troglodytes verus) cave use in southeastern Senegal. Int J Primatol. 2020;41:916-935. http://doi.org/10.1007/s10764-020-00165-4 [ Links ]
34. Nel C, Bradfield J, Lombard M, Val A. Taphonomic study of a modern baboon sleeping site at Misgrot, South Africa: Implications for large-bodied primate taphonomy in karstic deposits. J Paleolith Archaeol. 2021;4:4. http://doi.org/10.1007/s41982-021-00080-x [ Links ]
35. Zipfel B, Berger LR. New Cenozoic fossil-bearing site abbreviations for collections of the University of the Witwatersrand. Palaeontol Afr. 2009;44:77-81. [ Links ]
36. González-Aguilera D, Muñoz-Nieto A, Gómez-Lahoz J, Herrero-Pascual J, Gutierrez-Alonso G. 3D digital surveying and modelling of cave geometry: Application to Paleolithic rock art. Sensors. 2009;9:1108-1127. http://doi.org/10.3390/s90201108 [ Links ]
37. Doneus M, Verhoeven G, Fera M, Briese Ch, Kucera M, Neubauer W. From deposit to point cloud - a study of low-cost computer vision approaches for the straightforward documentation of archaeological excavations. Geoinformatics FCE CTU. 2011;6:81-88. http://doi.org/10.14311/gL6.11 [ Links ]
38. De Reu J, Plets G, Verhoeven G, De Smedt P, Bats M, Cherretté B, et al. Towards a three-dimensional cost-effective registration of the archaeological heritage. J Archaeol Sci. 2013;40:1108-1121. http://doi.org/10.1016/j.jas.2012.08.040 [ Links ]
39. Olson BR, Placchetti RA, Quartermaine J, Killebrew AE. The Tel Akko Total Archaeology Project (Akko, Israel): Assessing the suitability of multi-scale 3D field recording in archaeology. J Field Archaeol. 2013;38:244-262. http://doi.org/10.1179/0093469013Z.00000000056 [ Links ]
40. Bronikowski AM, Alberts SC, Altmann J, Packer C, Carey KD, Tatar M. The aging baboon: Comparative demography in a non-human primate. Proc Natl Acad Sci USA. 2002;99:9591-9595. http://doi.org/10.1073/pnas.142675599 [ Links ]
41. Garvin HM, Elliott MC, Delezene LK, Hawks J, Churchill SE, Berger LR, et al. Body size, brain size, and sexual dimorphism in Homo naledi from the Dinaledi Chamber. J Hum Evol. 2017;111:119-138. http://doi.org/10.1016/j.jhevol.2017.06.010 [ Links ]
42. Harcourt-Smith WEH, Throckmorton Z, Congdon KA, Zipfel B, Deane AS, Drapeau MSM, et al. The foot of Homo naledi. Nat Commun. 2015;6:8432. http://doi.org/10.1038/ncomms9432 [ Links ]
 Correspondence:
Correspondence:
Michael Everett
Email: Michael.C.Everett.19@dartmouth.edu
Received: 13 Mar. 2021
Revised: 28 July 2021
Accepted: 02 Aug. 2021
Published: 29 Nov. 2021
Editors: Margaret Avery, Jemma Finch
Funding: Claire Garber Goodman Fund, Department of Anthropology, Dartmouth College; Stamps Scholars Program, Dartmouth College
Supplementary Data
The supplementary data is available in pdf: [Supplementary data]


{kind=link}
{kind=link}
{kind=link}
{kind=link}