










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.11-12 Pretoria nov./dic. 2021
http://dx.doi.org/10.17159/sajs.2021/9736
REVIEW ARTICLE
The polyphagous shot hole borer beetle: Current status of a perfect invader in South Africa
Elmar van RooyenI; Trudy PaapII, III; Wilhelm de BeerII, III; Garyn TownsendIV; Shawn FellII, III; Wilma J. NelII, III; Seamus MorganV; Martin HillIV; Allan GonzalezVI; Francois RoetsI
IDepartment of Conservation Ecology and Entomology, Stellenbosch University, Stellenbosch, South Africa
IIDepartment of Biochemistry, Genetics and Microbiology, University of Pretoria, Pretoria, South Africa
IIIForestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, Pretoria, South Africa
IVCentre for Biological Control, Department of Zoology and Entomology, Rhodes University, Makhanda, South Africa
VDepartment of Genetics, Stellenbosch University, Stellenbosch, South Africa
VISchool of Forest, Fisheries and Geomatics Sciences, University of Florida, Gainesville, Florida, USA
ABSTRACT
The polyphagous shot hole borer (PSHB) beetle is a recent invader in South Africa. Together with its fungal symbiont, Fusarium euwallaceae, it can rapidly kill highly susceptible host plants. Its impact is most profound in urban areas, but it has also been found infesting important forestry, agricultural crop and native species. Since its first detection in 2012, PSHB has spread to all but one province in the country. The beetle-fungus complex has several biological traits that enhance its anthropogenically mediated dispersal, establishment and survival in novel environments - factors that have likely facilitated its rapid spread across the country. We review the history of the PSHB invasion in South Africa, its taxonomic status and the reasons for its rapid spread. We highlight its potential impact and challenges for its management. Finally, we provide an updated distribution map and list of confirmed host plants in South Africa. Of the 130 plant species identified as hosts, 48 of these (19 indigenous and 29 introduced) are reproductive hosts able to maintain breeding PSHB populations. These reproductive hosts may succumb to beetle infestations and act as 'pest-amplifiers'. The economic impact on urban forests, plantation forestry and agricultural crops may be severe, but the ecological impact of PSHB invasion in native ecosystems should not be underestimated.
SIGNIFICANCE:
• We provide an updated host list and distribution map for South Africa of the globally significant tree pest, the polyphagous shot hole borer (PSHB, Euwallacea fornicatus). The South African PSHB invasion represents the largest outbreak of this pest in its global invaded range. PSHB was confirmed to infest 130 plant species in urban, agricultural, and native ecosystems in South Africa, including 44 previously unreported hosts. Impact in South Africa is in its infancy but will likely be substantial to local economies and ecosystems. Mitigation has proven difficult, but numerous research projects have been initiated throughout the country.
Keywords: distribution, Euwallacea fornicatus, Fusarium euwallaceae, host range, invasion biology
Study species
Ambrosia beetles in the tribe Xyleborini (Coleoptera: Curculionidae: Scolytinae) are emerging as a group of globally important invasive insect pests.1Euwallacea, a genus within the Xyleborini, contains 45 species, including four lineages in the E. fornicatus sensu lato species complex: E. fornicatior, E. perbrevis, E. kuroshio and E. fornicatus. The taxonomy of the E. fornicatus species complex has been described as one of the most challenging in scolytine systematics2; however, the species complex has recently been resolved, with the name E. fornicatus now formally linked to the common name, polyphagous shot hole borer (PSHB) (Figure 1)24. Diagnostic characters separating these species have been described, and while the four clades on average differ morphologically, notable overlap in morphological characters precludes complete delimitation.2,4 The most reliable means of identification is therefore via DNA sequence comparisons2,4 or high-resolution melt assay of the 28S ribosomal subunit5.

PSHB originates from Southeast Asia3,6, and, like other ambrosia beetles, has a mutualistic association with fungi. Three fungal symbionts have been described from PSHB and infected plant hosts: Fusarium euwallaceae, Graphium euwallaceae and Paracremoniumpembeum.79These fungal symbionts are introduced into the galleries constructed by the beetles in host trees, via mandibular mycangia - specialised internal organs evolved for fungal transport.7-9 Most notable of these three fungi is the primary symbiont Fusarium euwallaceae. In addition to being the main food source for the beetle and its larvae, the fungus invades the vascular tissue of the tree, blocking the transport of water and nutrients, and resulting in Fusarium dieback.7-9 The majority of ambrosia beetles typically infest stressed, dead or dying trees; however, species within the E. fornicatus species complex are amongst the very few ambrosia beetles capable of infesting healthy plants10, and therefore present an important phytosanitary threat in their invaded range.
History of invasion
PSHB was first detected outside its native range in 2003 in urban California, USA, reported as the tea shot hole borer.11 In 2009, it was detected on avocado in Israel (as Euwallacea sp. nr fornicatus), but anecdotal evidence suggests that it was introduced as early as 2004.12 It is now known to infest avocado trees and many other woody plants in urban and agricultural areas in these countries.12,13 Even though exotic Euwallacea has been known from Hawaii since 191014, E. fornicatus was only confirmed among populations of E. perbrevis in 20205,15. PSHB was also recently (April 2020) detected in a greenhouse in Italy; the pest status is currently: present, under eradication.16
In 2017, as part of a plant sentinel project, PSHB was found infesting plane trees (Platanus x acerifolia) in a botanical garden in the KwaZulu-Natal Province, South Africa.17 However, a DNA sequence of PSHB was obtained from a specimen in Durban in 2012 as part of the Barcode of Life project (BOLD: ACC9773, ETKC270-13).3 This location is ca. 50 km southeast of the 2017 collection site, indicating a much earlier introduction into the country. Since then, the beetle has been found in eight of the nine South African provinces, reaching as far as 1000 km from the first detection locality, making it the largest current outbreak of this pest globally.
Factors driving invasion by PSHB in South Africa
Introduction and dispersal pathways
Central to the understanding of the likelihood of establishment of species beyond their native range are the characters that enhance their dispersal, i.e. departure, transport, and arrival in non-native environments.18 For PSHB, chances for departure are high due to their small size (females are 1.8-2.5 mm long while males are only 1.5-1.67 mm long) and cryptic colouration (dark to light brown).19 Therefore, they are easily missed during routine inspections when hitchhiking on goods and biological material such as nursery stock, even when not colonising the material.20 Their cryptic life cycle adds considerably to their successful departure as contaminants that colonise wood products.21 Ambrosia beetles typically infest host trees by constructing galleries deep within the vascular tissues.22 They are therefore easily missed in untreated wooden articles and packaging materials.23 Potential transport vectors for PSHB include aeroplanes, trains, trucks, cars, and boats. PSHB was most likely introduced into South Africa via ships as the earliest known record for it is close to two major ports.3 Subsequent spread throughout the country was probably aided by the movement of infested wood (e.g. firewood).
As hitchhikers, characters such as resistance to drastic temperatures, starvation, dehydration, or exposure to toxins enhance survival during the transport phase.18,24 As there is no evidence that PSHB is particularly resistant to any of these conditions, it likely survives during transport as contaminants due to their colonisation of wooden products.25 Wood acts as a buffer against rapid temperature fluctuations and is therefore a good insulator. Additionally, many wood types are not particularly porous, preventing chemical treatments from penetrating far enough to reach the beetle colony. Finally, moisture content is often fairly high, benefitting fungal growth and beetle survival26, but adults of PSHB are often found in fairly dry wood, where its second nutritional symbiont Graphium euwallaceae thrives8. As contaminants of wood, individuals may be resistant to phytosanitary treatments27 and are difficult to manage through application of a single phytosanitary measure23. Their small size and cryptic colouration also aid in preventing detection during inspections.28 In South Africa, there is an increased risk of accidental introductions due to limited phytosanitary inspections - a consequence of a lack of trained personnel and funding.29
Reproduction and propagule pressure
High propagule pressure can enhance the chances of dispersal at all three levels (departure, transport, and arrival).3032 Studies rearing PSHB on artificial diets have provided valuable data on beetle biology, phenology, sex ratios and temperature requirements.25,33 Development rates for PSHB are temperature dependent, with fmin, fmax and t determined to be 13.34 °C, 33.08 °C and 27.51 °C, respectively, with the number of degree days required for development from egg to adult estimated to be 398 ± 52.23.33 Fungal growth is also influenced by temperature, with optimum growth of F. euwallaceae occurring around 27 °C (Freeman S 2020, personal communication). Fungal growth rate is likely to play a role in brood size and may explain the observation of reduced brood sizes at higher temperatures.33
A single foundress female lays an average of 32.4 eggs, with the first adults developing in as little as 22 days at 24 °C.25 For PSHB, outbreeding is not required for reproduction; it has a haplodiploid mating system in which haploid males develop from unfertilised eggs.25 If a mated female initiates a colony, both male and female offspring may be produced. Sex ratios are heavily female biased, with only ca. 7% male beetles.25 These male offspring can mate with female offspring within the galleries, resulting in mated female progeny that can leave the natal gallery and start new colonies. However, a virgin female can construct a gallery by laying haploid eggs that will result in all-male offspring.25 She can then mate with her male offspring to produce diploid eggs.25 The result is that a single female beetle, whether mated or not, can start a new infestation with no indication that the population will suffer from inbreeding.3,34 For PSHB, successful establishment is therefore possible even with low propagule pressure.4
Colonies contain multiple overlapping generations and a colony from a single foundress can produce up to 57 reproductive female adults within 6-7 weeks.25 Infested wood generally contains multiple colonies from numerous foundress females and population sizes can be tremendously large and self-sustaining over relatively long periods of time. For example, a single piece of infested wood weighing ca. 500 g collected in South Africa produced 44 mature females and five males after drying. These adult females are able to produce ca. 1425 offspring (ca. 32.4 eggs per female) within 22 days at 24 °C if all are able to successfully construct galleries.25 It is clear that propagule pressure for PSHB is high, enhancing its chances of dispersal and establishment.35 It has also been shown that adult beetles and larvae are able to survive in cut wood, with females continuing to lay viable eggs.19 Studies from California have shown that beetles can continue to emerge for several months after felling of infested trees.19
Host range
PSHB is known to have a particularly broad host range, in which both the fungus and beetle are able to establish.3638 This increases the chances of a suitable host being encountered, thus increasing the chances of successful establishment. To obtain the PSHB host range for South Africa, observations made by researchers during field investigations and reports made by the public to the Forestry and Agricultural Biotechnology Institute, University of Pretoria, and Stellenbosch University were collated. Only taxa from which the primary fungal symbiont, F. euwallaceae, or a beetle individual were successfully isolated and identified using DNA sequence fingerprinting were included. Reproductive hosts include only those hosts that have been shown to be suitable for PSHB reproduction at least once (i.e. those with galleries containing eggs, larvae or multiple adults). Non-reproductive hosts include those from which either PSHB or F. euwallaceae was isolated, but no evidence of PSHB reproduction was found. In South Africa, signs of PSHB infestations have been confirmed from 130 tree species of which 48 are suitable for reproduction (i.e. reproductive hosts) and 82 can sustain fungal growth (i.e. non-reproductive hosts) (Table 1). This wide fungal host range ensures availability of suitable hosts near the introduction point. When beetle populations are very high, the insects appear to assess newly encountered hosts almost indiscriminately for fungal establishment13,38, explaining the high numbers of non-reproductive hosts. Such 'attacks' may also lead to severe decline in tree health due to Fusarium wilt13. Fusarium euwallaceae may also help PSHB to overcome tree defences and enhance successful colonisation of a host, even those that are only marginally suitable for beetle development.39
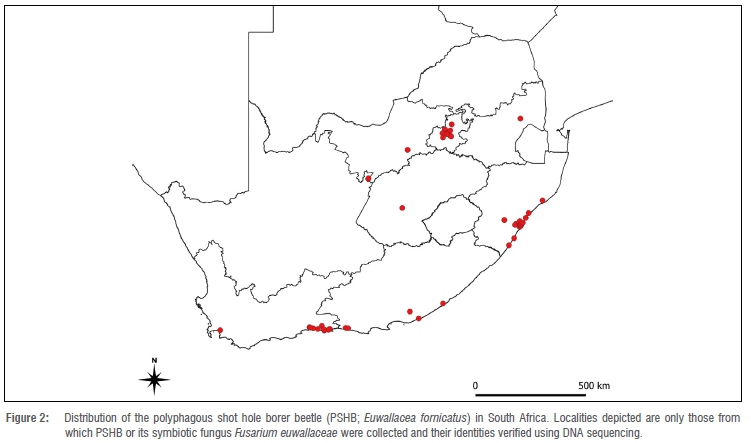
Distribution of PSHB in South Africa
As with the determination of the PSHB host range in South Africa, the PSHB distribution was mapped by collating confirmed (by DNA sequence fingerprinting) reports based on field observations made by researchers and reports made by the public. In South Africa, PSHB is widespread, and has a continuous distribution in both KwaZulu-Natal (between Durban and Richards Bay) and the southern Cape (between George and Plettenberg Bay) (Figure 2). This indicates a relatively long history in these areas where initial populations expanded from the points of introduction. Unlike males, females can fly, but most disperse only short distances (ca. 30 m from point of release40). This may seem unremarkable, but factors like wind can have tremendous influence on the number of dispersing individuals, the direction of dispersal and the dispersal distance.40 In Somerset West (Western Cape Province), E. fornicatus dispersed at least 3 km from the putative point of introduction in only 2 months, against prevailing winds. Therefore, when suitable hosts are present, PSHB can expand its range rapidly without direct human assistance. However, distances between closest confirmed PSHB populations in South Africa can be as far as 500 km (Figure 2). This can only be achieved when dispersal is aided by humans and human-mediated dispersal remains the most substantial form of longdistance transport and establishment of new colonies.
PSHB has a wide range of climatic conditions in which it can persist, increasing its chances of finding sites open for establishment. It is native to equatorial climates and has successfully established in temperate (California and Israel) and tropical (Hawaii) regions. Currently occupied climate zones, as defined by Köppen-Geiger and updated by Peel et al.41, include Af, Am, Aw, BSk, Csa, Csb, Cfa, Cfb and Cwa. Within South Africa, PSHB has established in at least six climatic zones: BSh, BSk, Cfa, Cfb, Csb and Cwb. In addition, Csa and Cwa are also present in South Africa, making much of the country climatically suitable for PSHB. While the minimum temperature threshold for PSHB reproduction has been determined to be 13.34 °C, beetles are able to persist at lower temperatures for extended periods.33 A cold tolerance study found significant mortality rates among colonies exposed to -5 °C and -1 °C; however, no mortality was observed for colonies exposed to 0 °C, 1 °C or 5 °C.25 In addition, PSHB galleries are often located deep within wood, where they are protected from extreme temperatures. Finally, microclimates exist in urban and agricultural settings, which may provide a conducive climate in an otherwise unsuitable area.
In South Africa, PSHB is currently mostly confined to urban environments, likely due to greater invasibility and anthropogenically mediated dispersal. Also, urban areas are far less biologically diverse than natural environments and offer less of a biological buffer.42 Typically, ambrosia beetles favour trees that are stressed in their natural environments.43 Even in novel environments this is often the case. Anthropogenic activities such as urbanisation, gardening, trampling, and pruning can cause tree stress.44 As a result, urban areas have high invasibility.42,45 Natural disturbances such as drought, storms, fire, and other pests can also produce added stress to trees in both urban and natural settings.46 Combined, these factors promote the spread and status of PSHB, but none has yet been comprehensively evaluated. Although PSHB is unlike many other ambrosia beetle species, in that it can easily infest healthy hosts10,47, if tree stress is still a contributing factor to epidemiology, a natural or anthropogenic disturbance in future could have catastrophic consequences.
Impact of PSHB
The first indication of PSHB in invaded habitats is benign. It tends to infest healthy hosts where 0.85-mm diameter holes penetrate through the bark directly into the sapwood (Figure 3a). Lesions may be visible on the outside of the tree (Figure 3b), but more commonly a brown to pinkish stain, resulting from colonisation by F. euwallaceae, can be seen spreading from the gallery into the vascular tissues below the bark (Figure 3c). Different hosts respond to infestation in different ways, thus complicating initial diagnosis. External signs of infestations include wet patches and gum exudates (tree resin flows), sugar fountains, and accumulating frass (mixture of sawdust and excreta) or noodles (compacted frass) (Figures 3d-g). Successful colonisation of reproductive hosts often results in wilting and death of the infested branch or the entire tree (Figure 3h).12,36 By this stage, beetle populations are well established. Detection in natural systems is very challenging due to a greater tree diversity, fewer people, and larger areas to survey.
The most notable agricultural impact of PSHB invasion has been on the avocado industries of Israel and California, USA. In Israel, it was detected in orchards in 2009, and by 2016 it had spread to nearly all avocado-growing areas in the country.48 Symptoms such as stem and branch breakages, wilting, dieback and death are common.48 Initial studies from Israel and California reported a high risk of branch dieback and mortality, raising concern about the potential of PSHB to severely impact avocado production worldwide.7,12,49 More recent observations, however, suggest impacts have not been as severe as anticipated, and that sanitation practices, including the removal of infested branches and trees, may offer a cost-effective management approach.38,48 Coleman et al.50 emphasise though, that their survey data represent only one time point and that further surveys are needed to confirm potential impacts to production. Caution should be taken in extrapolating this to the situation in South Africa due to differences in climates, cultivation practices and the relative importance of different cultivars for production. South Africa currently has around 17 500 ha avocado orchards, producing around 170 000 tons (or around ZAR700 million worth) of avocado annually. However, to date, no PSHB infestations have been found in commercial avocado orchards (only in backyard avocado trees). The only agricultural crops thus far affected by PSHB infestation in the country appear to be pecan and macadamia, but the effect on these crops appears limited, most likely as they are not reproductive hosts.
PSHB has also been found associated with numerous other agricultural crops such as cherry, apple, citrus, peach, guava, olive, grape vine and prune (Table 1). Whether any of these can act as reproductive hosts is still unclear as all reports are from urban settings. Even so, when grown near reproductive hosts, there is a strong possibility of infection by F. euwallaceae. Therefore, current research in South Africa is aimed at establishing whether these crops are susceptible to Fusarium wilt. Besides agriculturally important tree species, PSHB have been observed on Acacia mearnsii, a commercially grown forestry species in South Africa, but to date, no trees in commercial plantations have been reported to be infested. Based on observations on Acacia spp. from plantations in southeast Asia, there are concerns surrounding the threat posed to the South African forestry industry.51
In its global invaded range, the most notable impacts of PSHB are on urban trees, with hundreds of different species susceptible to beetle infestation and Fusarium wilt13,38,48 (Table 1). In the USA, nearly 300 tree species have been reported to be susceptible to infestation by PSHB, with 55 of these suitable for reproduction.38 PSHB has caused injury and/or mortality to thousands of ornamental trees in urban forests in southern California.38 In South Africa, visual surveys undertaken in invaded urban areas of Johannesburg (Gauteng Province), Knysna, George and Somerset West (Western Cape Province) showed that nearly all Quercus robur (English oak), Acer negundo (box elder) and other maples (Acer spp.) will die when infested by PSHB (Z.W.d.B., T.P, F.R., unpublished data). Currently about 50% of London plane trees (Platanus x acerifolia) infested with PSHB in these areas are in decline and likely to die.
Unfortunately, data on the species and numbers of trees planted in South African cities are lacking, and, for many of the reproductive host tree species planted in urban environments, no data are available on the percentage of trees that will die. However, a recent system dynamics model to estimate the potential economic impact of PSHB in South African urban environments could be substantial, including impacts on stormwater run-off, losses to ecosystem services and carbon dioxide sequestration, and reduced shade contributing to remediation of the urban heat island effect and pollution reduction.52 There is increasing evidence linking widespread death of trees in urban environments following pest and pathogen invasions, and increased human mortality related to cardiovascular and respiratory tract illness.53 This may be particularly relevant in large cities with an urban/industrial interface where trees play an important role in pollution remediation. As trees succumb, falling trees and branches may impact human safety directly by causing traffic accidents and damaging vehicles and infrastructure such as buildings, fences, and power lines.
Negative economic impacts are incurred with the removal of infested trees and their replacement. However, tree removal has additional negative effects, such as a possible reduction in property values. In South Africa this has already resulted in at least one court case where PSHB infestations were not reported to potential buyers. The cost of tree removal in Somerset West amounted to just over ZAR7000/tree in 2019. With more than 600 trees earmarked for removal in this town alone, the City of Cape Town requested a stipend of ZAR3 million in 2020 to monitor and remove infested trees (Ramabulana P 2019, personal communication). Services provided by these infested trees (e.g. carbon sequestering, providing shade, aesthetics, and emotional health) are also severely negatively affected by their removal. The replacement cost for trees that have already been removed in Somerset West is estimated to be between ZAR4.5 million and ZAR5.8 million, depending on tree species and labour costs (E.v.R., unpublished data). Therefore, by far the largest proportion of the social cost due to PSHB invasion in South Africa will be incurred in urban areas.52
Most worrying perhaps is the ability of PSHB to infest and kill native trees in its invaded ranges. Of the reproductive host species recorded in South Africa, 19 are indigenous (Table 1), and severe ecological consequences may result after invasion of natural environments, as was seen with the destruction of endemic willow forests in the Tijuana River Valley.54 Approximately 120 000 (30%) native willows (Salix spp.) were killed following infestation by E. kuroshio, a close relative of PSHB.55 Loss of shade resulted in increased river temperatures, which negatively impacted aquatic organisms. Recovery from the loss of large established trees will take time and encroachment by invasive alien plants has prevented full recovery of some stands.56 The invasion of PSHB into South Africa's natural forests is still in its infancy, and there is no empirical data yet on the numbers of trees expected to succumb. Long-term monitoring plots have, however, been established in natural forests in Knysna and George, with some preliminary data available from ongoing surveys in these locations. In the ca. 3000 km2 surveyed, 2195 trees were assessed for PSHB presence. Of these, 217 individuals (ca. 10%) were infested and 191 (8.7%) showed signs of beetle reproduction, which may lead to dieback and death (E.v.R., G.T., unpublished data). In South Africa, the loss of even a single species from an ecosystem can be catastrophic. For example, Virgilia oroboides is a native reproductive host for PSHB. It is an important pioneer forest species in the southern Cape that protects forests from severe climatic fluctuations and fire and houses a large number of native organisms.57,58 Elimination of this single species could have irreversible consequences for native forest integrity.
Mitigation
Experiences from California, Israel, and South Africa indicate that the management of the PSHB is particularly challenging. To summarise, miniscule size presents detection challenges, and the haplodiploid and sib-mating system means the introduction of a single female can lead to the establishment of a new infestation. As the beetle feeds on symbiotic fungi, it has low tree host specificity. It is able to persist in many climatic zones and climate may not be critical for its establishment. The reproductive cycle is fast under optimum conditions, and if temperatures fall outside the suitable range, larvae and beetles can survive for some time and continue development should temperatures become suitable again. Spread is difficult to manage due to the ease of movement of infested wood and green waste. Despite the challenges, there are several mitigation strategies that may aid in reducing the impact of PSHB.
Biological control is generally difficult for ambrosia beetles owing to their cryptic habits and inbreeding success.48 There is no biocontrol agent available against PSHB. However, Californian researchers have been investigating PSHB in its natural range to identify potential natural enemies (Stouthamer R 2021, personal communication). In South Africa, investigations into biocontrol options are being initiated.
Control of low levels of PSHB infestation may be achievable by direct injection of insecticides and fungicides. This not only maximises pesticide impact but reduces environmental contamination. Control of PSHB in more heavily infested trees seems unattainable even with the direct application of chemicals.59 Chemicals that show the most promise include combinations of insecticide and fungicide treatments, like emamectin benzoate or bifenthrin combined with propiconazole.59-61 These controls are currently being investigated in South Africa. A report by the California Forest Pest Council62, however, highlights that chemical treatment of infested trees in Orange County currently requires repeated applications (up to three times per year) using a combination of insecticides and fungicides and various application methods. The cost of chemical treatment is therefore expected to be high, and even if proven effective, their application in natural settings will not be feasible. Additionally, the application of commercial chemicals for prolonged periods could result in phytotoxicity.63-65 In South Africa, no insecticides or fungicides that have been evaluated with scientific rigour for efficacy in PSHB management have been registered for use.
As there is no effective means to control pre-existing beetle infestations within a tree, the best alternative is to reduce the propagule number by physical removal of highly infested reproductive host trees. This reduces propagule pressure and potential of spread to new localities. This strategy has been used to reduce PSHB impact in infested avocado orchards.38,48 Infested material needs to be properly treated to reduce beetle survival. Currently, best protocols dictate chipping of the wood to pieces smaller than 5 cm followed by solarisation under plastic sheeting for 4-6 weeks at 55 °C.19,66 Movement of infested wood and wood chips should be avoided.32,67 Minimising the spread of invasive reproductive host plant species is crucial, as these may create pathways for autonomous spread.68
Conclusions
PSHB has proven to be one of the most difficult tree pests to manage. This is largely due to its ability to colonise a massive variety of tree species and its ease of anthropogenically mediated long-distance dispersal. In the absence of a rapidly implemented nationally coordinated strategy, and without strong stakeholder and public engagement, PSHB has easily spread throughout the country.69 Thus far, it has invaded numerous major cities, and, because South Africa is climatically suitable, PSHB is expected to rapidly spread to new areas. Impacts of the invasion are currently only in their infancy and major social and ecological costs are expected in the near future.52 Continued monitoring of the spread of the beetle is essential if we are to mitigate its impact. Mitigation will prove difficult, but is of the utmost importance. Even halting the movement of infested wood and wood products will be problematic. Despite the informal urban firewood trade being a widespread and important energy and income security source for many poor urban residents, its prevention is seen as the most effective way to curb long-distance spread. One benefit of PSHB invasion is an increase in public awareness of invasive organisms and their potential impact, at least in heavily affected areas. This increase in public awareness has also provided an opportunity in terms of public education and, combined with the creation of a multidisciplinary and multi-institutional Polyphagous Shot Hole Borer Research Network in South Africa, will help considerably in continued efforts in research, monitoring and control of invasive species.
Acknowledgements
We thank the Natural Resource Management Programme of the South African Department of Forestry, Fisheries and the Environment (DFFE) for funding, noting that this publication does not necessarily represent the views or opinions of DFFE or its employees. We acknowledge the Tree Protection Cooperative Programme (TPCP) and the Forestry and Agricultural Biotechnology Institute (FABI) of the University of Pretoria for funding and infrastructure support. Comments from anonymous reviewers helped to improve our manuscript.
Competing interests
We declare that there are no competing interests.
Authors' contributions
E.v.R.: Conceptualisation; methodology; data collection; validation; data curation; writing - the initial draft; writing - revisions. T.P: Data collection; validation; data curation; writing - revisions. W.d.B.: Conceptualisation; methodology; data collection; validation; data curation; student supervision; project management; and funding acquisition. G.T.: Data collection; validation; writing - revisions. S.F.: Data collection; validation; writing - revisions. W.J.N.: Data collection; validation; writing - revisions. S.M. Data collection; validation; writing - revisions. M.H.: Conceptualisation; methodology; data collection; validation; data curation; writing - revisions; project management; and funding acquisition. A.G.: Methodology; data collection; validation; data curation; writing - revisions. F.R.: Conceptualisation; methodology; data collection; validation; data curation; writing - revisions; student supervision; project leadership; project management; and funding acquisition.
References
1. Hulcr J, Stelinski LL. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu Rev Entomol. 2017;62:285-303. https://doi.org/10.1146/annurev-ento-031616-035105 [ Links ]
2. Smith SM, Gomez DF, Beaver RA, Hulcr J, Cognato AI. Reassessment of the species in the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) complex after the rediscovery of the "lost" type specimen. Insects. 2019;10(9):1-11. https://doi.org/10.3390/insects10090261 [ Links ]
3. Stouthamer R, Rugman-Jones P Thu PQ, Eskalen A, Thibault T, Hulcr J, et al. Tracing the origin of a cryptic invader: Phylogeography of the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) species complex. Agric For Entomol. 2017;19(4):366-375. https://doi.org/10.1111/afe.12215 [ Links ]
4. Gomez DF, Skelton J, Steininger MS, Stouthamer R, Rugman-Jones P, Sittichaya W, et al. Species delineation within the Euwallacea fornicatus (Coleoptera: Curculionidae) complex revealed by morphometric and phylogenetic analyses. Insect Syst Divers. 2018;2(6):1-11. https://doi.org/10.1093/isd/ixy018 [ Links ]
5. Rugman-Jones PF, Au M, Ebrahimi V Eskalen A, Gillett CPDT, Honsberger D, et al. One becomes two: Second species of the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) species complex is established on two Hawaiian Islands. PeerJ. 2020;8, e9987. https://doi.org/10.7717/peerj.9987 [ Links ]
6. Storer CG, Breinholt JW, Hulcr J. Wallacellus is Euwallacea: Molecular phylogenetics settles generic relationships (Coleoptera: Curculionidae: Scolytinae: Xyleborini). Zootaxa. 2015;3974(3):391-400. https://doi.org/10.11646/zootaxa.3974.3.6 [ Links ]
7. Freeman S, Sharon M, Maymon M, Mendel Z, Protasov A, Aoki T, et al. Fusarium euwallaceae sp. nov. - a symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia. 2013;105(6):1595-1606. https://doi.org/10.3852/13-066 [ Links ]
8. Freeman S, Sharon M, Dori-Bachash M, Maymon M, Belausov E, Maoz Y et al. Symbiotic association of three fungal species throughout the life cycle of the ambrosia beetle Euwallacea nr. fornicatus. Symbiosis. 2016;68(1-3):115-128. https://doi.org/10.1007/s13199-015-0356-9 [ Links ]
9. Lynch SC, Twizeyimana M, Mayorquin JS, Wang DH, Na F, Kayim M, et al. Identification, pathogenicity and abundance of Paracremonium pembeum sp. nov. and Graphium euwallaceae sp. nov. - two newly discovered mycangial associates of the polyphagous shot hole borer (Euwallacea sp.) in California. Mycologia. 2016;108(2):313-329. https://doi.org/10.3852/15-063 [ Links ]
10. Gomez DF, Lin W, Gao L, Li Y New host plant records for the Euwallacea fornicatus (Eichhoff) species complex (Coleoptera: Curculionidae: Scolytinae) across its natural and introduced distribution. J Asia Pac Entomol. 2019;22(1):338-340. https://doi.org/10.1016/j.aspen.2019.01.013 [ Links ]
11. Rabaglia RJ, Dole SA, Cognato AI. Review of American Xyleborina (Coleoptera: Curculionidae: Scolytinae) occurring north of Mexico, with an illustrated key. Ann Entomol Soc Am. 2006;99(6):1034-1056. https://doi.org/10.1603/0013-8746(2006)99[1034:ROAXCC]2.0.CO;2 [ Links ]
12. Mendel Z, Protasov A, Sharon M, Zveibil A. An Asian ambrosia beetle Euwallacea fornicatus and its novel symbiotic fungus Fusarium sp. pose a serious threat to the Israeli avocado industry. Phytoparasitica. 2012;40:235-238. https://doi.org/10.1007/s12600-012-0223-7 [ Links ]
13. Eskalen A, Stouthamer R, Lynch SC, Rugman-Jones PF, Twizeyimana M, Gonzalez A, et al. Host range of Fusarium dieback and its ambrosia beetle (Coleoptera: Scolytinae) vector in southern California. Plant Dis. 2013;97(7):938-951. https://doi.org/10.1094/pdis-11-12-1026-re [ Links ]
14. Swezey OH. Some injurious insects in Samoa which do not occur in the Hawaiian Islands. Proc Hawaiian Entomol Soc. 1941;11(1):127-130. http://hdl.handle.net/10125/16047 [ Links ]
15. Mitchell A, Maddox C. Bark beetles (Coleoptera: Curculionidae: Scolytinae) of importance to the Australian macadamia industry: An integrative taxonomic approach to species diagnostics. Aust J Entomol. 2010;49(2):104-113. https://doi.org/10.1111/j.1440-6055.2010.00746.x [ Links ]
16. EPPO. First report of Euwallacea fornicatus in Italy [document on the Internet]. c2020 [cited 2021 Feb 25]. Available from: https://gd.eppo.int/reporting/article-6772 [ Links ]
17. Paap T, De Beer ZW, Migliorini D, Nel WJ, Wingfield MJ. The polyphagous shot hole borer (PSHB) and its fungal symbiont Fusarium euwallaceae: A new invasion in South Africa. Australas Plant Pathol. 2018;47(2):231-237. https://doi.org/10.1007/s13313-018-0545-0 [ Links ]
18. Gippet JM, Liebhold AM, Fenn-Moltu G, Bertelsmeier C. Human-mediated dispersal in insects. Curr Opin Insect Sci. 2019;35:96-102. https://doi.org/10.1016/j.cois.2019.07.005 [ Links ]
19. Jones ME, Paine TD. Effect of chipping and solarization on emergence and boring activity of a recently introduced ambrosia beetle (Euwallacea sp., Coleoptera: Curculionidae: Scolytinae) in southern California. J Econ Entomol. 2015;108(4):1852-1859. https://doi.org/10.1093/jee/tov169 [ Links ]
20. Liebhold AM, Brockerhoff EG, Garrett LJ, Parke JL, Britton KO. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front Ecol Environ. 2012;10(3):135-143. https://doi.org/10.1890/110198 [ Links ]
21. Meurisse N, Rassati D, Hurley BP, Brockerhoff EG, Haack RA. Common pathways by which non-native forest insects move internationally and domestically. J Pest Sci. 2019;92(1):13-27. https://doi.org/10.1007/s10340-018-0990-0 [ Links ]
22. Hulcr J, Dunn RR. The sudden emergence of pathogenicity in insect-fungus symbioses threatens naive forest ecosystems. Proc R Soc B Biol Sci. 2011;278(1720):2866-2873. https://doi.org/10.1098/rspb.2011.1130 [ Links ]
23. Allen E, Noseworthy M, Ormsby M. Phytosanitary measures to reduce the movement of forest pests with the international trade of wood products. Biol Invasions. 2017;19(11):3365-3376. https://doi.org/10.1007/s10530-017-1515-0 [ Links ]
24. Renault D, Laparie M, McCauley SJ, Bonte D. Environmental adaptations, ecological filtering, and dispersal central to insect invasions. Annu Rev Entomol. 2018;63(1):345-368. https://doi.org/10.1146/annurev-ento-020117-043315 [ Links ]
25. Cooperband MF, Stouthamer R, Carrillo D, Eskalen A, Thibault T, Cossé AA, et al. Biology of two members of the Euwallacea fornicatus species complex (Coleoptera: Curculionidae: Scolytinae), recently invasive in the U.S.A., reared on an ambrosia beetle artificial diet. Agric For Entomol. 2016;18(3):223-237. https://doi.org/10.1111/afe.12155 [ Links ]
26. Huang J, Kautz M, Trowbridge AM, Hammerbacher A, Raffa KF, Adams HD, et al. Tree defence and bark beetles in a drying world: Carbon partitioning, functioning and modelling. New Phytol. 2020;225(1):26-36. https://doi.org/10.1111/nph.16173 [ Links ]
27. Whattam M, Clover G, Firko M, Kalaris T. The biosecurity continuum and trade: Border operations. In: Gordh G, McKirdy S, editors. The handbook of plant biosecurity. Dordrecht: Springer; 2014. p. 149-188. https://doi.org/10.1007/978-94-007-7365-3_6 [ Links ]
28. Eschen R, Rigaux L, Sukovata L, Vettraino AM, Marzano M, Grégoire JC. Phytosanitary inspection of woody plants for planting at European Union entry points: A practical enquiry. Biol Invasions. 2015;17(8):2403-2413. https://doi.org/10.1007/s10530-015-0883-6 [ Links ]
29. Parliament of the Republic of South Africa. Plant health (phytosanitary) Bill. 2017 p. 1-24. [ Links ]
30. Haack RA, Petrice TR, Wiedenhoeft AC. Incidence of bark and woodboring insects in firewood: A survey at Michigan's Mackinac bridge. J Econ Entomol. 2010;103(5):1682-1692. https://doi.org/10.1603/ec10041 [ Links ]
31. Liebhold AM, Brockerhoff EG, Kimberley M. Depletion of heterogeneous source species pools predicts future invasion rates. J Appl Ecol. 2017;54(6):1968-1977. https://doi.org/10.1111/1365-2664.12895 [ Links ]
32. Grousset F, Grégoire JC, Jactel H, Battisti A, Beloglavec AB, Hrasovec B, et al. The risk of bark and ambrosia beetles associated with imported non-coniferous wood and potential horizontal phytosanitary measures. Forests. 2020;11(3):1-17. https://doi.org/10.3390/f11030342 [ Links ]
33. Umeda C, Paine T. Temperature can limit the invasion range of the ambrosia beetle Euwallacea nr. fornicatus. Agric For Entomol. 2019;21(1):1-7. https://doi.org/10.1111/afe.12297 [ Links ]
34. Peer K, Taborsky M. Outbreeding depression, but no inbreeding depression in haplodiploid ambrosia beetles with regular sibling mating. Evolution. 2005;59(2):317-323. https://doi.org/10.1111/j.0014-3820.2005.tb00992.x [ Links ]
35. Simberloff D. The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst. 2009;40(1):81-102. https://doi.org/10.1146/annurev.ecolsys.110308.120304 [ Links ]
36. Dodge C, Carrillo J, Eskalen A, Stouthamer R. Evidence for symbiont promiscuity in two invasive ambrosia beetles (Coleoptera: Scolytinae: Euwallacea spp.). Paper presented at: Annual Meeting of the Entomological Society of America; 2017 November 5-8; Denver, Colorado, USA. p. 165. [ Links ]
37. Freeman S, Miller G, Protasov A, Maymon M, Elazar M, David-Schwartz R, et al. Aposymbiotic interactions of three ambrosia beetle fungi with avocado trees. Fungal Ecol. 2019;39:117-130. https://doi.org/10.1016/J.FUNECO.2018.11.007 [ Links ]
38. Lynch SC, Eskalen A, Gilbert GS. Host evolutionary relationships explain tree mortality caused by a generalist pest-pathogen complex. Evol Appl. 2021;14(4):1083-1094. https://doi.org/10.1111/eva.13182 [ Links ]
39. Norris DM, Baker JM. A minimal nutritional substrate required by Fusarium solani to fulfill its mutualistic relationship with Xyleborus ferrugineus. Ann Entomol Soc Am. 1968;61(6):1473-1475. https://doi.org/10.1093/aesa/61.6.1473 [ Links ]
40. Owens D, Seo M, Montgomery WS, Rivera MJ, Stelinski LL, Kendra PE. Dispersal behaviour of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae) in avocado groves and estimation of lure sampling range. Agric For Entomol. 2019;21(2):199-208. https://doi.org/10.1111/afe.12321 [ Links ]
41. Peel MC, Finlayson BL, McMahon TA. Updated world map of the Köppen-Geiger climate classification. Hydrol Earth Syst Sci Discuss. 2007;4(2):439-473. https://doi.org/10.5194/hess-11-1633-2007 [ Links ]
42. Potgieter LJ, Douwes E, Gaertner M, Measey J, Paap T, Richardson DM. Biological invasions in South Africa's urban ecosystems: Patterns, processes, impacts, and management. In: Van Wilgen B, Measey J, Richardson DM, Wilson JR, Zengeya TA, editors. Biological Invasions in South Africa. New York: Springer International Publishing; 2020. p. 275-309. https://doi.org/10.1007/978-3-030-32394-3 [ Links ]
43. Ploetz RC, Hulcr J, Wingfield MJ, De Beer ZWZ Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013;97(7):856-872. https://doi.org/10.1094/PDIS-01-13-0056-FE [ Links ]
44. Paap T, Burgess TI, Wingfield MJ. Urban trees: Bridge-heads for forest pest invasions and sentinels for early detection. Biol Invasions. 2017;19(12):3515-3526. https://doi.org/10.1007/s10530-017-1595-x [ Links ]
45. Richardson DM, Pysek P Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog Phys Geogr. 2006;30:409-431. https://doi.org/10.1191/0309133306pp490pr [ Links ]
46. Sukopp H, Starfinger U. Disturbance in urban ecosystems. In: Walker LR, editor. Ecosystems of disturbed ground. Amsterdam: Elsevier; 1999. p. 397-412. [ Links ]
47. Kühnholz S, Borden JH, Uzunovic A. Secondary ambrosia beetles in apparently healthy trees: Adaptations, potential causes and suggested research. Integr Pest Manag Rev. 2003;6(3-4):209-219. https://doi.org/10.1023/A:1025702930580 [ Links ]
48. Mendel Z, Protasov A, Maoz Y, Maymon M, Miller G, Elazar M, et al. The role of Euwallacea nr. fornicatus (Coleoptera: Scolytinae) in the wilt syndrome of avocado trees in Israel. Phytoparasitica. 2017;45(3):341-359. https://doi.org/10.1007/s12600-017-0598-6 [ Links ]
49. Eskalen A, Gonzalez A, Wang DH, Twizeyimana M, Mayorquin JS, Lynch SC. First report of a Fusarium sp. and its vector tea shot hole borer (Euwallacea fornicatus) causing fusarium dieback on avocado in California. Plant Dis. 2012;96(7):1070. https://doi.org/10.1094/PDIS-03-12-0276-PDN [ Links ]
50. Coleman TW, Poloni AL, Chen Y Thu PQ, Li Q, Sun J, et al. Hardwood injury and mortality associated with two shot hole borers, Euwallacea spp., in the invaded region of southern California, USA, and the native region of Southeast Asia. Ann For Sci. 2019;76(61):1-18. https://doi.org/10.1007/s13595-019-0847-6 [ Links ]
51. Hulcr J, Black A, Prior K, Chen CY, Li HF. Studies of ambrosia beetles (Coleoptera: Curculionidae) in their native ranges help predict invasion impact. Florida Entomol. 2017;100(2):257-261. https://doi.org/10.1653/024.100.0219 [ Links ]
52. De Wit MP, Crookes DJ, Blignaut JN, De Beer ZW, Paap T, Roets F, et al. Invasion of the polyphagous shot hole borer beetle in South Africa: A preliminary assessment of the economic impacts [document on the Internet]. c2021 [cited 2021 Jun 29]. Available from: https://doi.org/10.21203/rs.3.rs-220132/v1 [ Links ]
53. Donovan GH, Butry DT, Michael YL, Prestemon JP, Liebhold AM, Gatziolis D, et al. The relationship between trees and human health. Am J Prev Med. 2013;44(2):139-145. https://doi.org/10.1016/j.amepre.2012.09.066 [ Links ]
54. Boland JM. The impact of an invasive ambrosia beetle on the riparian habitats of the Tijuana River Valley, California. PeerJ. 2016;4(6), e2141. https://doi.org/10.7717/peerj.2141 [ Links ]
55. Boland JM, Woodward DL. Impacts of the invasive shot hole borer (Euwallacea kuroshio) are linked to sewage pollution in southern California: The Enriched Tree Hypothesis. PeerJ. 2019;7, e6812. https://doi.org/10.7717/peerj.6812 [ Links ]
56. Boland JM, Uyeda KA. The ecology and management of the Kuroshio Shot Hole Borer in the Tijuana River Valley [document on the Internet]. c2020 [cited 2021 Mar 25]. Available from: https://trnerr.org/wp-content/uploads/2020/05/KSHB-TRValley2020.pdf [ Links ]
57. Machingambi NM, Roux J, Dreyer LL, Roets F. Bark and ambrosia beetles (Curculionidae: Scolytinae), their phoretic mites (Acari) and associated Geosmithia species (Ascomycota: Hypocreales) from Virgilia trees in South Africa. Fungal Biol. 2014;118(5-6):472-483. https://doi.org/10.1016/j.funbio.2014.03.006 [ Links ]
58. Coetsee C, Wigley BJ. Virgilia divaricata may facilitate forest expansion in the afrotemperate forests of the southern Cape, South Africa. Koedoe. 2013;55(1), Art. #1128. https://doi.org/10.4102/koedoe.v55i1.1128 [ Links ]
59. Mayorquin JS, Carrillo JD, Twizeyimana M, Peacock BB, Sugino KY, Na F, et al. Chemical management of invasive shot hole borer and Fusarium dieback in California sycamore (Platanus racemosa) in southern California. Plant Dis. 2018;102(7):1307-1315. https://doi.org/10.1094/PDIS-10-17-1569-RE [ Links ]
60. Eatough Jones M, Paine TD. Potential pesticides for control of a recently introduced ambrosia beetle (Euwallacea sp.) in southern California. J Pest Sci. 2018;91(1):237-246. https://doi.org/10.1007/s10340-017-0866-8 [ Links ]
61. Grosman DM, Eskalen A, Brownie C. Evaluation of emamectin benzoate and propiconazole for management of a new invasive shot hole borer (Euwallacea nr. fornicatus, Coleoptera: Curculionidae) and symbiotic fungi in California sycamores. J Econ Entomol. 2019;112(3):1267-1273. https://doi.org/10.1093/jee/toy423 [ Links ]
62. California Forest Pest Council. California Forest Pest Conditions [document on the Internet]. c2021 [cited 2021 Feb 21]. Available from: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fseprd896758.pdf [ Links ]
63. Jones ME, Kabashima J, Eskalen A, Dimson M, Mayorquin JS, Carrillo JD, et al. Evaluations of insecticides and fungicides for reducing attack rates of a new invasive ambrosia beetle (Euwallacea sp., Coleoptera: Curculionidae: Scolytinae) in infested landscape trees in California. J Econ Entomol. 2017;110(4):1611-1618. https://doi.org/10.1093/jee/tox163 [ Links ]
64. Bonilla-Landa I, De la Cruz OL, Sanchéz-Rangel D, Ortíz-Castro R, Rodriguez-Haas B, Barrera-Méndez F, et al. Design, synthesis and biological evaluation of novel fungicides for the management of Fusarium dieback disease. J Mex Chem Soc. 2018;62(3):86-98. https://doi.org/10.29356/jmcs.v62i3.531 [ Links ]
65. Van Der Werf HMG. Assessing the impact of pesticides on the environment. Agric Ecosyst Environ. 1996;60(2-3):81-96. https://doi.org/10.1016/S0167-8809(96)01096-1 [ Links ]
66. Chen Y Coleman TW, Poloni AL, Nelson L, Seybold SJ. Reproduction and control of the invasive polyphagous shot hole borer, Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae), in three species of hardwoods: Effective sanitation through felling and chipping. Environ Entomol. 2020;49(5):1155-1163. https://doi.org/10.1093/ee/nvaa103 [ Links ]
67. Haack RA. Intercepted Scolytidae (Coleoptera) at US ports of entry: 1985-2000. Integr Pest Manag Rev. 2001;6:254-282. https://doi.org/10.1023/A:1025715200538 [ Links ]
68. Van der Colff D, Dreyer LL, Valentine A, Roets F. Invasive plant species may serve as a biological corridor for the invertebrate fauna of naturally isolated hosts. J Insect Conserv. 2015;19(5):863-875. https://doi.org/10.1007/s10841-015-9804-3 [ Links ]
69. Paap T, Wingfleld MJ, De Beer ZW, Roets F. Lessons from a major pest invasion: The polyphagous shot hole borer in South Africa. S Afr J Sci. 2020;116(11/12), Art. #8757. https://doi.org/10.17159/sajs.2020/8757 [ Links ]
 Correspondence:
Correspondence:
Francois Roets
Email: fr@sun.ac.za
Received: 22 Feb. 2021
Revised: 22 June 2021
Accepted: 28 June 2021
Published: 29 Nov. 2021
Editors: Teresa Coutinho, Salmina Mokgehle
Funding: South African Department of Forestry, Fisheries and the Environment, University of Pretoria


{kind=link}
{kind=link}
{kind=link}