










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.5-6 Pretoria may./jun. 2021
http://dx.doi.org/10.17159/sajs.2021/9145
RESEARCH ARTICLE
Darting towards Storm Shelter: A minute dinosaur trackway from southern Africa
Emese M. Bordy
Department of Geological Sciences, University of Cape Town, Cape Town, South Africa
ABSTRACT
Theropod dinosaurs are considered the main terrestrial carnivores in the Jurassic and Cretaceous. Their rise to dominance has been linked to, among others, body size changes in their early history, especially across the Triassic-Jurassic boundary. However, to qualitatively assess such temporal trends, robust skeletal and trace fossil data sets are needed globally. The richly fossiliferous southern African continental rock record in the main Karoo Basin offers an unparalleled perspective for such investigations. Herein, by documenting a newly discovered Early Jurassic trackway of very small, functionally tridactyl tracks near Storm Shelter (Eastern Cape) in South Africa, the track record can be expanded. Based on ichnological measurements at the ichnosite and digital 3D models, the footprint dimensions (length, width, splay), locomotor parameters (step length, stride, speed), and body size estimates of the trackmaker are presented. In comparison to other similar tracks, these footprints are not only the smallest Grallator-like tracks in the Clarens Formation, but also the most elongated dinosaur footprints in southern Africa to date. The tracks also show that the small-bodied bipedal trackmaker dashed across the wet sediment surface at an estimated running speed of -12.5 km/h. During the dash, either as a predator or as a prey, the trackmaker's small feet sunk hallux-deep into the sediment. The tracking surface is overgrown by fossilised microbial mats, which likely enhanced the footprint preservation. Based on track morphometrics and the regional dinosaur skeletal record, the trackmakers are attributed to Megapnosaurus rhodesiensis (formerly Syntarsus rhodesiensis), a small-to-medium-sized, early theropod common in southern Africa.
SIGNIFICANCE:
• A newly discovered Early Jurassic theropod trackway in South Africa contains not only the smallest tracks in the Clarens Formation, but also the most elongated dinosaur footprints in southern Africa to date.
• The tracks show that the small bipedal trackmaker dashed across the wet sediment surface at an estimated running speed of -12.5 km per hour.
• During the run, the trackmaker's feet sunk so deeply into the sediment that even the forwards-directed halluces were impressed.
Keywords: tridactyl footprints, Grallator-like tracks, small theropods, southern Gondwana, Jurassic Clarens Formation
Introduction
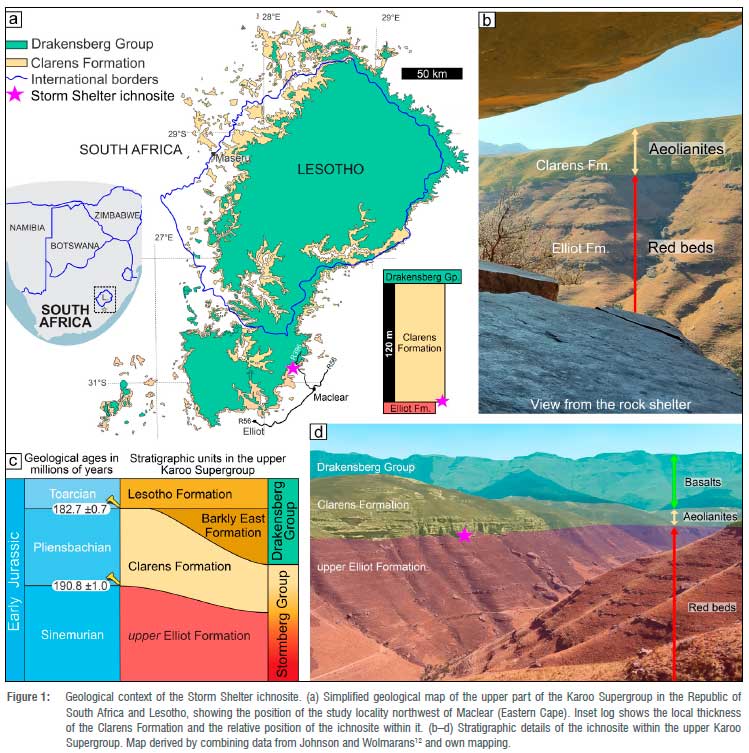
Mounting evidence for body size changes in theropods1-3, a class of carnivorous dinosaurs, during the dawn of the dinosaurs is increasingly placing the southern African fossil record into the focus of global palaeontological studies. Late Triassic to Early Jurassic dinosaur footprints, for which southern Gondwana, especially southern Africa, is an excellent archive, are particularly useful proxies for such biogeological investigations. In particular, the Lower Jurassic of southern Africa (i.e. the upper Stormberg Group; Figure 1) is key in this regard as: (1) it was deposited in a semi-arid continental ecosystem of rivers, lakes and deserts; (2) it is richly fossiliferous, and (3) it provides clues on how life recovered after the global biodiversity crisis event that occurred -200 million years ago.4,5
This study reports on the smallest dinosaur footprints, forming a single trackway, in the Clarens Formation (uppermost Stormberg Group; Figure 1) of South Africa and shows that these tracks are also, to date, the most elongated dinosaur footprints from the Early Jurassic of southern Africa. These tiny, slender tracks in the lower Clarens Formation are part of the Lower Jurassic footprint assemblage, which in southern Africa is contained in the upper Elliot Formation (Hettangian-Sinemurian) and in the comformably overlying Clarens Formation (Sinemurian-Pliensbachian).4 These two rock units also contain the two largest (footprint lengths: 55-57 cm)6-9 and the smallest (footprint length: -6.5 cm)10 tridactyl tracks on record thus far, all reported from Lesotho. Because the current tracks are part of the ichnofauna of the lower Clarens Formation, which is likely Sinemurian4, they enrich the Early Jurassic track record locally and globally, and thus can contribute to the evaluation of the temporal patterns in the footprint record, which, in turn, can add to the ongoing debate on dinosaur body size changes across the Triassic-Jurassic boundary1-3,11.
Geological background
The dinosaur footprints investigated in this study were discovered north of Maclear in the Eastern Cape of South Africa (Figure 1) by a local resident, Mrs Adele Moore, and her field party in 2014 along a footpath leading to the Storm Shelter archaeological site.13 Found at the same altitude as the rock art site, the very fine-grained sandstone slab, containing the single trackway of five well-impressed dinosaur footprints, is part of the lowermost Clarens Formation (Figure 1). Here, at Storm Shelter, and throughout the main Karoo Basin (South Africa and Lesotho), the basal Clarens Formation, which is likely Sinemurian4,14 in age, is characterised by tabular, fine- to medium-grained sandstone beds and subordinate mudstones. Suites of sedimentary structures (e.g. horizontal lamination, cross-bedding, ripple cross-lamination, desiccation cracks; Supplementary figure 1) in these rocks point to deposition in decelerating powerful traction currents and stagnant water.15,16 Overall, these rocks indicate that the ancient wet aeolian ecosystem was prone to intermittent flash flooding and drying episodes.15,16 While being less fossiliferous than the conformably underlying Hettangian to Sinemurian upper Elliot Formation, the palaeontological record of the Clarens Formation is diverse, both in vertebrate skeletal remains and trace fossils4,6,14,1718, particularly in footprints, and most notably in dinosaur tracks, as recently summarised by Mukaddam et al.19
Methods
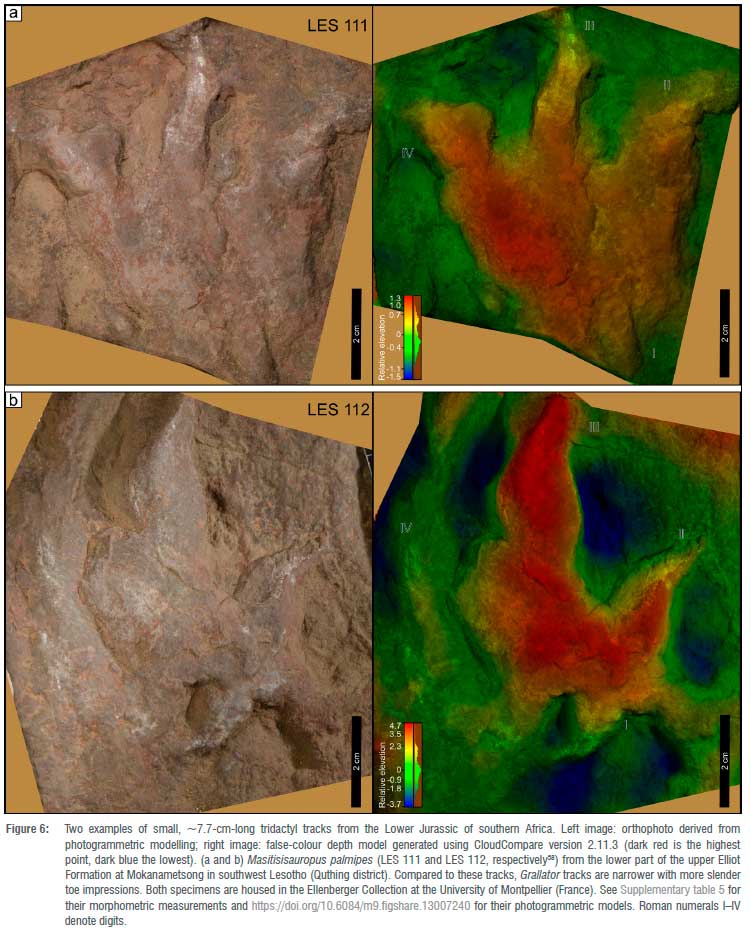
The footprints investigated in this study were subjected to morphometric measurements as shown in Figure 2. Moreover, for comparative purposes, four additional footprints collected from Lesotho, two Grallator-like (LES 283 and 288) and two /Inomoepus-like (LES 111 and 112) tracks from the Clarens and upper Elliot Formations, respectively, were also briefly analysed for this study. These four Lesotho tracks were collected by Dr Paul Ellenberger pre-1970, and are now housed in the Ellenberger Collection at the University of Montpellier (France).
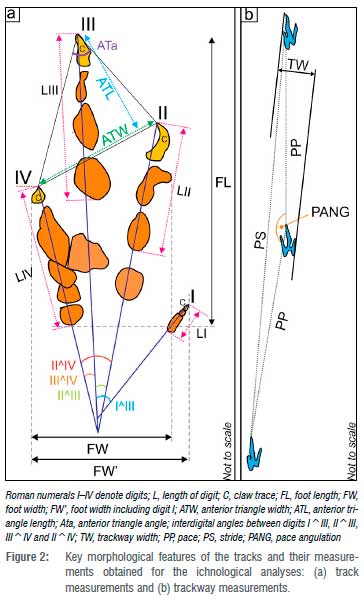
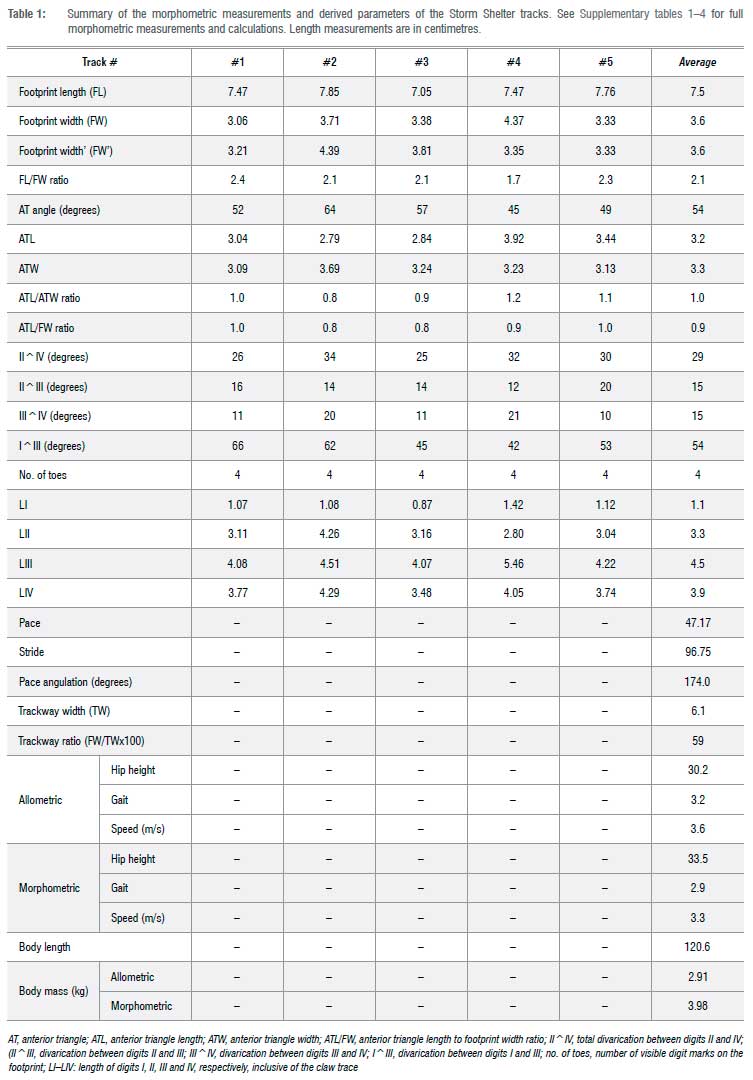
The ichnological morphometric measurements and the standard proxies derived from them, which estimate the trackmaker's hip height, gait, body length and speed, are summarised in Table 1 and detailed in Supplementary tables 1-5 (with references). The relevant modus operandi is explained elsewhere5,8,9,10,19 and in references therein. The track measurements were taken physically in the field using a calliper. After photographing each track with a Nikon D5100 digital camera (focal length 50 mm), individual photogrammetric 3D models and false-colour depth maps were generated with Agisoft Metashape Professional (version 1.6.4) and CloudCompare (v.2.11.0) software packages, respectively. Following the standard protocol of Falkingham et al.20, the photogrammetric 3D models and relevant raw data are publicly available online via this open access data repository link: https://doi.org/10.6084/m9.figshare.13007240
Results
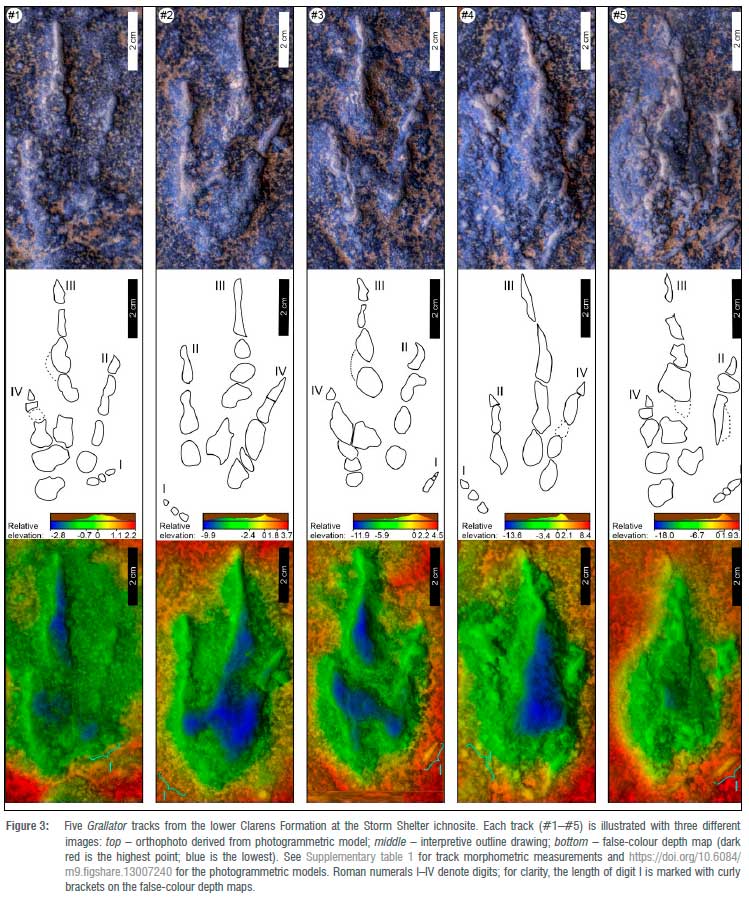
The Storm Shelter ichnosite preserves a single trackway of five, very small, narrow, elongated and strongly mesaxonic footprints (Figure 3, Table 1, Supplementary table 1) with the following main parameters (average values): length 7.5 cm, width 3.6 cm, length/width ratio >2.1. The ratios of the anterior triangle21 length to width and the anterior triangle length to track width are 1 and 0.9, respectively. Manus tracks were not observed.
These elongate, digitigrade tracks preserve evidence for claw marks at the tips of the three well-impressed digits and one lightly impressed digit, indicative of a hindfoot that was functionally tridactyl and featured a well-developed hallux (digit I). The slender digit impressions only sporadically show discrete, oval digital pads, most likely due to extensive sediment collapse within the tracks, which would also suggest that these are likely penetrative tracks22,23 (i.e. the foot penetrated the substrate much more deeply than is apparent from the track outlines on the tracking surface). The penetrative nature of the tracks is further supported by the very narrow distal portion of digit III. Relative to digits II and IV, digit III is better impressed and extends beyond digits II and IV, which are subequal. Both the apex of the anterior triangle and the divarication between digits II and IV form acute angles of 54° and 29°, respectively. On average, digit I is ~1 cm long and its long axis encloses an acute angle of 54° with the long axis of digit III. The variability of the position of the digit I impression relative to the other digits (Figure 3, Table 1) indicates that the hallux did not have any functional use for locomotion or stability.
The single trackway that these five pes tracks form is narrow (~6.1 cm), and relative to the footprint length of 7.5 cm, preserves long strides of nearly 1 m (Figure 4, Table 1, Supplementary table 2). The long axis of digit III is essentially parallel to the trackway axis. The average pace is 47.6 cm with an angulation of 172°, and the track width to trackway width ratio is 59%, which define a narrow-gauge trackway. Along the trackway, the morphological quality of the tracks is uniform, although well-developed wrinkle structures, a type of microbially induced sedimentary structure, are present both inside and outside the tracks on the tracking surface (Figure 4 and Supplementary figure 1), and interfere with the outlines of some of the tracks (e.g. see the posterior of track #1 in Figure 3). The microbially induced sedimentary structures and the digit I impression demonstrate that the tracks are not underprints, but rather primary, penetrative footprints impressed directly and deeply on the tracking surface.
Because of the incomplete/missing digital pad impressions, some ill-defined imprint walls, and superimposition of microbial structures, all five Storm Shelter tracks rank low, around 1.5 on the recently proposed24,25, four-point (0-1-2-3) track-grading scale. Because of the suboptimal morphological quality of the digital pad impressions, only a very tentative phalangeal formula of 1(?2)-2-3(?4)-4-0 (not counting the claw traces) is inferred from the collective assessment of the five tracks.
Using standard ichnological formulae26,27 (detailed in Supplementary tables 3 and 4), the trackmaker's hip height, body length, body mass, gait and speed were estimated as summarised in Table 1. Based on these parameters, which are essentially derived from the average foot length and stride values, the trackmaker's hip height was ~32 cm, its body length was ~120 cm and it weighed ~3-4 kg (Table 1, Supplementary table 3). The body size of this digitigrade, obligate bipedal trackmaker thus is comparable to small-sized, long-legged, ground-dwelling extant birds, which fall into a size class between a very tall domestic rooster and a bustard or a wild turkey (hip height 0.39 m and body mass of ~5-7 kg; cf.28). The track data are insufficient to determine whether the small trackmaker was a juvenile or a small-sized but fully grown adult.
The trackmaker's allometric and morphometric gait values are 3.2 and 2.9, respectively, indicating a constant 'running' gait26,27 across the sediment surface (Table 1, Supplementary tables 3 and 4). The allometric and morphometric speed estimations show that the trackmaker ran at an average speed of 3.45 m/s (i.e. ~12.42 km/h). The relatively rapid locomotion of the trackmaker is also suggested by the narrow trackway, as during fast movement, running bipedal animals place their feet closer to the midline (cf.29). The stride/footprint length ratio for the Storm Shelter trackway is 12.9 and thus is only slightly greater than the median value30 for trackways in this size category (i.e. foot length of 5-10 cm) from the early Mesozoic and attributed to theropods. This suggests that the trackmaker was moving in a typical manner for a small theropod.
Discussion
The morphological characteristics describing the Storm Shelter footprint proportions (Table 1) closely resemble those of Grallator-like tracks; however, due to the moderate to low quality of these tracks, their ichnotaxonomic treatment is not warranted here. Grallator is an ichnogenus that refers to globally occurring tracks of bipedal theropod dinosaurs of early Mesozoic age. Grallator tracks are small (<15 cm long), elongate with a footprint length/width ratio of >2, narrow digit divarication angles (10-30°) of the slender, pointy toes and a great anterior projection of digit III (i.e. mesaxonic tracks; e.g.21,26,31,32). Moreover, Grallator tracks are digitigrade, often preserve claw traces and occasionally also the impression of digit I28, and thus the ichnogenus refers to functionally tridactyl tracks.
In the early Mesozoic ichnological record of the main Karoo Basin of southern Africa, tracks referable to Grallator are not rare.610,33-37 Being only ~1 cm longer than the smallest Grallator tracks in southern Africa (described10 from the lower part of the upper Elliot Formation in Lesotho), the Storm Shelter tracks, with their average footprint length of 7.5 cm, are among the smallest Grallator-like tracks in the Lower Jurassic of this region, and certainly the smallest of all dinosaur tracks in the Clarens Formation to date.
In the Clarens Formation, only four tracks that are morphologically similar to Grallator were mentioned by Ellenberger6 from Lesotho: three tracks of similar size (~10 cm long) from zone B5 (his figures 127-129) and one from zone B6 (no illustration). In spite of efforts to relocate these tracks in the field, only the latter track (G. molapoi - Leribe-Molapo) as well as G. (Paragrallator) matsiengensis (Matsieng, his figure 128) are available for direct study as specimens LES 288 (holotype) and LES 283 (the only replica of the now lost holotype), respectively, in the Ellenberger Collection at the University of Montpellier (France). Because these ichnotaxa are partially preserved, severely obscured by extramorphological features and lacking information regarding the foot anatomy of the trackmaker, they can be considered nomina nuda, but being the only tracks identified as 'Grallator' from the Clarens Formation, the digital 3D models of these tracks are documented here (Figure 5, Supplementary table 5) for sake of completeness. Morphometric measurements show that these 'Grallator' tracks are >33% longer (average length: 11.3 cm) than the Storm Shelter tracks. The only other footprints in the Clarens Formation that have been likened to Grallator are large, ~40-cm-long tridactyl tracks reported33 from the eastern Free State (Uniondale) in South Africa. However, these large, broad tracks, due to their key track morphological parameters (e.g. length-to-width ratio of 1.2; total divarication of 71°), are not considered here to be attributable to Grallator (see 8,21,32,35 and references therein for a more detailed discussion on the ichnotaxonomic considerations of these tracks).
Very small tridactyl tracks from Lesotho have been reported5,6,10 from the upper Elliot Formation (Hettangian-Sinemurian, zone B1 of Ellenberger). However, with the exception of the minute Grallator tracks10 in the lower part of the upper Elliot Formation at Lephoto in the Roma valley, compared to which the Storm Shelter tracks are similar in length but more elongated, all other very small tridactyl tracks (footprint length <9 cm) from the Elliot Formation seem to be consistent with ichnological parameters of either Trisauropodiscus-like or //nomoepus-like tracks (Figure 6, Supplementary table 1; also see5,10). Compared to these older, likely pre-Sinemurian4, tridactyl tracks (Figure 6), the Storm Shelter tracks are typically more elongated with narrow digit divarication angles (10-30°) and a greater anterior projection of digit III (i.e. more mesaxonic; see Supplementary table 1). All in all, these comparisons show that, thus far, the Storm Shelter tracks are among the most elongated (i.e. high length/width ratio) Grallator-like tracks reported from the Lower Jurassic of southern Africa, and the smallest in the Clarens Formation.
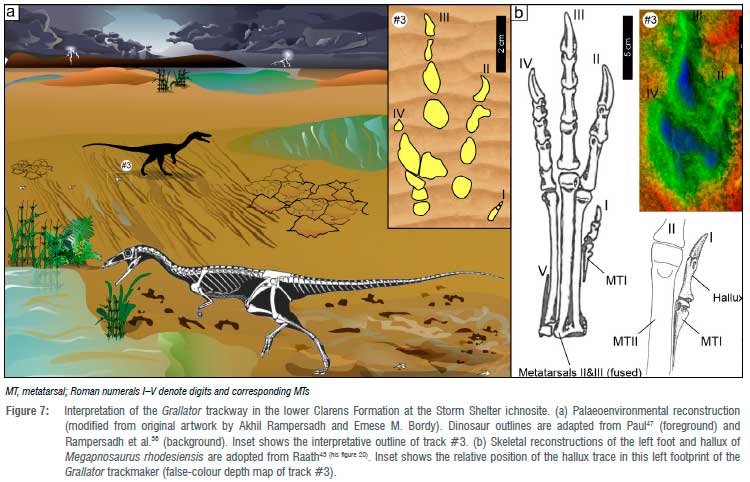
Although Grallator tracks rarely preserve28 impressions of digit I, remarkably, all Grallator-like tracks at Storm Shelter are associated with hallux (digit I) traces (Figure 3). Moreover, unlike in typical Grallator tracks26,31,39, the impression of the hallux is not directed medially or to the rear, but forward. Figure 3 shows how these ~1-cm-long impressions point in the forward direction, at ~54° (average) away from the long axis of digit III. The hallux impressions are not only important for identifying the trackmaker (see below) but also for determining that the trackmaker's feet sunk hallux-deep into the wet, soft sediment while the animal ran across the surface. A pliable, squidgy tracking surface also explains why digits II, III and IV in these tracks lack well-defined digital pad impressions, imprint walls, etc. (Figure 3), and thus limit the tracks' ichnotaxonomic assessment and conclusive attribution to any specific trackmaker. The wrinkle structures (Figure 4, Supplementary figure 1), which are linked to fossilised microbial mats and are abundant on this tracking surface, were possibly present already at the time of trackmaking and thus enhanced the footprint generation and overall preservation. Microbially induced sedimentary structures are known to be associated with dinosaur tracks in the southern African10,34,35 and global40-42 ichnofossil record. Likely, these sediment-binding biofilms developed as localised algal blooms flourished in shallow, stagnant pools of water that were generated in ephemeral downpours and that evaporated over time (Figure 7). Although the host Clarens Formation at Storm Shelter does preserve desiccation cracks (Supplementary figure 1), there are none preserved on the tracking surface itself, which possibly remained wet, or at least moist, until it was buried by the next layer of flash flood generated sediment (Figure 7).
Grallator is commonly attributed to early theropod dinosaurs that were obligatory bipeds with long, thin toes, including well-developed halluces (e.g.21,26,31,32). Based on the morphometric parameters of the Storm Shelter tracks (Figure 3, Table 1, Supplementary table 1) and the regional Early Jurassic bone fossil record, a likely trackmaker candidate is Megapnosaurus rhodesiensis (formerly Syntarsus rhodesiensis and Coelophysis rhodesiensis39,40). This early theropod is a coelophysoid dinosaur (Figure 7) that is common in the same stratigraphic interval (i.e. Lower Jurassic) of southern Africa, especially in Zimbabwe (e.g.43-45). A similar coelophysoid dinosaur, Coelophysis bauri, is often assumed to have made the early Mesozoic Grallator tracks in North America (e.g.26,31,32,46-48). Both of these early theropods were small-to-medium-sized, lightly built, agile, carnivorous bipeds and are often found in richly fossiliferous strata where entire animal populations are jumbled together as partially intact or complete skeletons that also include articulated foot bones.43 Foot reconstructions show that these early theropods had four pedal digits and a pes phalangeal formula (inclusive of unguals) of 2:3:4:5:0.43 Reconstructions also show that digit I (the hallux; Figure 7b) did not touch the ground during the steeply digitigrade locomotion of the animal (Figure 7a; e.g.43,48,49). Moreover, the well-formed hallux and associated foot bones of Megapnosaurus rhodesiensis preserve anatomical features that indicate that the hallux was forwards directed26,43, alongside digit II (Figure 7b). Raath43(p.93) remarked that the hallux 'retained a specialised function in life. Its use as a grooming accessory seems quite feasible'. The congruence in the morphological features of this foot skeleton and the tracks (including the non-reversed hallux configuration) is used here to suggest that the Storm Shelter footprints likely belong to Megapnosaurus rhodesiensis. It is noteworthy that in other early Mesozoic Grallator tracks26,31,39,48, the orientation of the hallux impression is reversed (posterior directed) in contrast to the original anatomical configuration. This backward-directed hallux mark resulted from the way the forward-directed hallux interacted with the sediment during the track-generating process as demonstrated by Gatesy et al.48 In the Storm Shelter tracks, however, the hallux impression closely corresponds to the typical anatomical (forward) orientation of the digit I in coelophysoid dinosaurs43,48, and this makes the Storm Shelter tracks unique among Grallator-like tracks. It is probable that this anterior-directed hallux impression resulted from the fortuitous combination of the microbially influenced substrate consistency that allowed the penetrative tracks to form, the trackmaker's tiny body proportions and the fast speed of motion.
Theropods were the main terrestrial carnivores in the post-Triassic part of the Mesozoic. Their smaller bodied varieties, like the trackmaker of the Storm Shelter tracks, most probably took on the dual role of predator and prey, and thus had good reason to leave behind tracks indicative of a running gait. The ability to run and occasionally sprint at speeds up to 12.5 km/h, even in the smallest individuals, must have been a great advantage for these agile, highly successful predators43 that had to adapt to an increasingly harsh, desert ecosystem, prone to flash flooding and dry spells in the Early Jurassic of southern Africa4 (Figure 7).
The minute, elongated Storm Shelter tracks together with the region's smallest10 and two largest6-9 tridactyl tracks are taken as evidence, not only of the abundance but also the size diversity of the Early Jurassic theropod dinosaurs in southern Africa. Given the regional abundance and diversity of the footprint record, the dinosaur tracks of southern Africa, in all shapes and sizes, remain an important proxy for meaningfully assessing concepts on macroevolutionary changes in dinosaur body size during the early Mesozoic.1-3,50 The true potential of this rich ichnological record is only achievable if the collected, but largely undescribed materials in various museum collections, as well as new discoveries like this one at the Storm Shelter, are quantified and integrated with the global early Mesozoic ichnological and osteological fossil records.
Acknowledgements
I acknowledge Mrs Adele Moore, not only for the discovery of the site in 2014 (in the company of Ms Julie Moore and Mr Neels van der Westhuizen), but also for granting and arranging access to the site and providing companionship during my field work in 2020. I also acknowledge curator Ms Suzanne Jiquel for access to the Ellenberger Collection at the Universitè de Montpellier (France). I am thankful to Fabien Knoll and Raquel López-Antonanzas, for many insightful discussions on the Early Jurassic palaeontology of southern Africa and for being wonderful hosts during my study visit in Montpellier. Jonah Choiniere broadened my mind about the evolution of theropod halluces. Finally, I thank Robert Nagy and Lilla Bordy-Nagy for their field assistance, feedback and overall support during this family (COVID-19 lockdown) project. This work was supported by the South African National Research Foundation (Competitive Programme for Rated Researches grant number 113394) and DST-NRF Centre of Excellence in Palaeosciences (grant number CoE PAL 2020-2021). The funders had no role in study design, data collection or any other aspect of the research. I received no financial benefit from this research.
Competing interests
I declare that there are no competing interests.
Data availability
The ichnological photogrammetric data that support the findings of this study are openly available in Figshare at https://doi.org/10.6084/m9.figshare.13007240, and include photographs used in the photogrammetric models, and the cleaned and aligned 3D models of the tracks figured in this paper (Figures 3, 5, 6).
References
1. Benson RB, Campione NE, Carrano MT, Mannion PD, Sullivan C, Upchurch P et al. Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage. PLoS Biol. 2014;12(5), e1001853. https://doi.org/10.1371/journal.pbio.1001853 [ Links ]
2. Griffin CT, Nesbitt SJ. Does the maximum body size of theropods increase across the Triassic-Jurassic boundary? Integrating ontogeny, phylogeny, and body size. Anat Rec. 2020;303(4):1158-1169. https://doi.org/10.1002/ar.24130 [ Links ]
3. Marsh AD, Rowe TB. A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona. J Paleontol. 2020;94(S78):1-3. https://doi.org/10.1017/jpa.2020.14 [ Links ]
4. Bordy EM, Abrahams M, Sharman GR, Viglietti PA, Benson RB, McPhee BW, et al. A chronostratigraphic framework for the upper Stormberg Group: Implications for the Triassic-Jurassic boundary in southern Africa. Earth-Sci Rev. 2020;203, Art. #103120. https://doi.org/10.1016/j.earscirev.2020.103120 [ Links ]
5. Bordy EM, Rampersadh A, Abrahams M, Lockley MG, Head HV. Tracking the Pliensbachian-Toarcian Karoo firewalkers: Trackways of quadruped and biped dinosaurs and mammaliaforms. PLoS ONE. 2020;15(1), e0226847. https://doi.org/10.1371/journal.pone.0226847 [ Links ]
6. Ellenberger P Les niveaux paléontologiques de première apparition des mammifères primordiaux en Afrique du Sud et leur ichnologie: établissement de zones stratigraphiques détaillées dans le Stormberg du Lesotho (Afrique du Sud) (Trias supérieur ä Jurassique) [The palaeontological levels of first appearance of primordial mammals in South Africa and their ichnology: Establishment of detailed stratigraphic zones in the Stormberg of Lesotho (South Africa) (Upper Triassic to Jurassic)]. In: Haughton SH, editor. Proceedings of the 2nd IUGS Symposium on Gondwana Stratigraphy and Palaeontology. Pretoria: Council for Scientific and Industrial Research; 1970. p. 343-370. French. [ Links ]
7. Ellenberger P. Contribution ä la classification des Piste de vertébrés du Trias: les types du Stormberg d'Afrique du Sud (I) [Contribution to the classification of the Triassic Vertebrate Track: The Stormberg types of South Africa (I)]. Montpellier: Palaeovertebrata, Memoire Extraordinaire; 1972. French. [ Links ]
8. Sciscio L, Bordy EM, Abrahams M, Knoll F, McPhee BW. The first megatheropod tracks from the Lower Jurassic upper Elliot Formation, Karoo Basin, Lesotho. PLoS ONE. 2017;12(10), e0185941. https://doi.org/10.1371/journal.pone.0185941 [ Links ]
9. Abrahams M, Sciscio L, Reid M, Bordy EM. Large tridactyl dinosaur tracks from the Early Jurassic of southern Gondwana-uppermost Elliot Formation, Upper Moyeni, Lesotho. Ann Soc Geol Pol. 2020;90(1):1-26. https://doi.org/10.14241/asgp.2020.0 [ Links ]
10. Abrahams M, Bordy EM, Sciscio L, Knoll F. Scampering, trotting, walking tridactyl bipedal dinosaurs in southern Africa: Ichnological account of a Lower Jurassic palaeosurface (upper Elliot Formation, Roma Valley) in Lesotho. Hist Biol. 2017;29(7):958-975. https://doi.org/10.1080/08912963.2016.1267164 [ Links ]
11. Lucas SG, Tanner LH. The missing mass extinction at the Triassic-Jurassic boundary. In: Tanner LH, editor. The Late Triassic World: Earth in a time of transition. Cham: Springer International Publishing; 2018. p. 721-825. [ Links ]
12. Johnson M, Wolmarans L. Simplified geological map of the Republic of South Africa and Kingdoms of Lesotho and Swaziland. Pretoria: Council for Geoscience; 2008. https://www.geoscience.org.za/images/DownloadableMaterial/RSA_Geology.pdf [ Links ]
13. Blundell G, Lewis-Williams D. Storm Shelter: An important new rock art find in South Africa. S Afr J Sci. 2001;97(1-2):43-46. https://hdl.handle.net/10520/EJC97281 [ Links ]
14. Knoll F. The tetrapod fauna of the Upper Elliot and Clarens formations in the main Karoo Basin (South Africa and Lesotho). Bull Soc Géol Fr. 2005;176(1):81-91. https://doi.org/10.2113/176.1.81 [ Links ]
15. Beukes NJ. Stratigraphy and sedimentology of the Cave Sandstone stage, Karoo System. In: Haughton SH, editor. Proceedings of the 2nd IUGS Symposium on Gondwana Stratigraphy and Palaeontology. Pretoria: Council for Scientific and Industrial Research; 1970. p. 321-341. [ Links ]
16. Bordy EM, Head HV. Lithostratigraphy of the Clarens Formation (Stormberg Group, Karoo Supergroup), South Africa. S Afr J Geol. 2018;121(1):119-130. https://doi.org/10.25131/sajg.121.0009 [ Links ]
17. Kitching JW, Raath MA. Fossils from the Elliot and Clarens Formations (Karoo Sequence) of the northeastern Cape, Orange Free State and Lesotho, and a suggested biozonation based on tetrapods. Palaeontol Afr. 1984;25:111-125. [ Links ]
18. Viglietti PA, McPhee BW, Bordy EM, Sciscio L, Barrett PM, Benson RBJ, et al. Biostratigraphy of the Massospondylus Assemblage Zone (Stormberg Group, Karoo Supergroup), South Africa. S Afr J Geol. 2020;123(2):249-262. https://doi.org/10.25131/sajg.123.0018 [ Links ]
19. Mukaddam R, Bordy EM, Lockley MG, Chapelle KEJ. Reviving Kalosauropus, an Early Jurassic sauropodomorph track from southern Africa (Lesotho). Hist Biol. 2020; Art. #1834542. http://dx.doi.org/10.1080/08912963.2020.1834542 [ Links ]
20. Falkingham PL, Bates KT, Avanzini M, Bennett M, Bordy EM, Breithaupt BH, et al. A standard protocol for documenting modern and fossil ichnological data. Palaeontology. 2018;61(4):469-480. https://doi.org/10.1111/pala.12373 [ Links ]
21. Lockley MG. New perspectives on morphological variation in tridactyl footprints: Clues to widespread convergence in developmental dynamics. Geol Q. 2009;53(4):415-432. [ Links ]
22. Gatesy SM, Falkingham PL. Hitchcock's Leptodactyli, penetrative tracks, and dinosaur footprint diversity. J Vertebr Paleontol. 2020;40(3), e1781142. https://doi.org/10.1080/02724634.2020.1781142 [ Links ]
23. Falkingham PL, Turner ML, Gatesy SM. Constructing and testing hypotheses of dinosaur foot motions from fossil tracks using digitization and simulation. Palaeontology. 2020;63(6):865-880. https://doi.org/10.1111/pala.12502 [ Links ]
24. Belvedere M, Farlow JO. A numerical scale for quantifying the quality of preservation of vertebrate tracks. Dinosaur tracks: the next steps. In: Falkingham PL, Marty D, Richter A, editors. Dinosaur tracks: The next steps. Bloomington, IN: Indiana University Press; 2016. p. 92-98. [ Links ]
25. Marchetti L, Belvedere M, Voigt S, Klein H, Castanera D, Díaz-Martínez I, et al. Defining the morphological quality of fossil footprints. Problems and principles of preservation in tetrapod ichnology with examples from the Palaeozoic to the present. Earth-Sci Rev. 2019;193:109-145. https://doi.org/10.1016/j.earscirev.2019.04.008 [ Links ]
26. Thulborn T. Dinosaur tracks. London: Chapman and Hall; 1990. [ Links ]
27. Weems RE. Locomotor speeds and patterns of running behavior in non-maniraptoriform theropod dinosaurs. Bull N M Mus Nat Hist Sci. 2006;37:379-389. [ Links ]
28. Gatesy SM, Biewener AA. Bipedal locomotion: Effects of speed, size and limb posture in birds and humans. J Zool. 1991;224(1):127-147. https://doi.org/10.1111/j.1469-7998.1991.tb04794.x [ Links ]
29. Bishop PJ, Clemente CJ, Weems RE, Graham DF, Lamas LP Hutchinson JR, et al. Using step width to compare locomotor biomechanics between extinct, non-avian theropod dinosaurs and modern obligate bipeds. J R Soc Interface. 2017;14(132), Art. #20170276. https://doi.org/10.1098/rsif.2017.0276 [ Links ]
30. Farlow JO, Coroian D, Currie PJ. Noah's ravens: Interpreting the makers of tridactyl dinosaur footprints. Bloomington, IN: Indiana University Press; 2018. [ Links ]
31. Olsen PE, Smith JB, McDonald NG. Type material of the type species of the classic theropod footprint genera Eubrontes, Anchisauripus, and Grallator (Early Jurassic, Hartford and Deerfield basins, Connecticut and Massachusetts, USA). J Vertebr Paleontol. 1998;18(3):586-601. https://doi.org/10.1080/02724634.1998.10011086 [ Links ]
32. Lucas SG, Klein HE, Lockley MG, Spielmann JA, Gierlinski GD, Hunt AP et al. Triassic-Jurassic stratigraphic distribution of the theropod footprint ichnogenus Eubrontes. Bull N M Mus Nat Hist Sci. 2006;37:86-93. [ Links ]
33. Raath MA, Yates AM. Preliminary report of a large theropod dinosaur trackway in Clarens Formation sandstone (Early Jurassic) in the Paul Roux district, northeastern Free State, South Africa. Palaeontol Afr. 2005;41:101-104. [ Links ]
34. Wilson JA, Marsicano CA, Smith RM. Dynamic locomotor capabilities revealed by early dinosaur trackmakers from Southern Africa. PLoS ONE. 2009;4(10), e7331. https://doi.org/10.1371/journal.pone.0007331 [ Links ]
35. Sciscio L, Bordy EM, Reid M, Abrahams M. Sedimentology and ichnology of the Mafube dinosaur track site (Lower Jurassic, eastern Free State, South Africa): A report on footprint preservation and palaeoenvironment. PeerJ. 2016;4, e2285. https://doi.org/10.7717/peerj.2285 [ Links ]
36. Rampersadh A, Bordy EM, Sciscio L, Abrahams M. Dinosaur behaviour in an Early Jurassic palaeoecosystem - uppermost Elliot Formation, Ha Nohana, Lesotho. Ann Soc Geol Pol. 2018;88:163-179. https://doi.org/10.14241/asgp.2018.010 [ Links ]
37. Abrahams M, Bordy EM, Knoll F. Hidden for one hundred years: A diverse theropod ichnoassemblage and cross-sectional tracks from the historic Early Jurassic Tsikoane ichnosite (Clarens Formation, northern Lesotho, southern Africa). Hist Biol. 2020:1-6. https://doi.org/10.1080/08912963.2020.1810681 [ Links ]
38. Ellenberger P. Contribution ä la classification des pistes de vertébrés du Trias: les types du Stormberg d'Afrique du Sud (II) (Hème partie: le Stormberg Superieur-I. Le biome de la zone B/1 ou niveau de Moyeni: ses biocénoses) [Contribution to the classification of the tracks of Triassic vertebrates: The types of the Stormberg of South Africa (II) (Hème part: the Stormberg Superior - I. The biome of zone B / 1 or level of Moyeni: its biocenoses). Montpellier: Palaeovertebrata, Memoire Extraordinaire; 1974. French. [ Links ]
39. Weems RE. Evidence for bipedal prosauropods as the likely Eubrontes track-makers. Ichnos. 2019;26(3):187-215. https://doi.org/10.1080/10420940.2018.1532902 [ Links ]
40. Razzolini NL, Belvedere M, Marty D, Paratte G, Lovis C, Cattin M, et al. Megalosauripus transjuranicus ichnosp. nov. A new Late Jurassic theropod ichnotaxon from NW Switzerland and implications for tridactyl dinosaur ichnology and ichnotaxomy. PLoS ONE. 2017;12(7), e0180289. https://doi.org/10.1371/journal.pone.0180289 [ Links ]
41. Castanera D, Belvedere M, Marty D, Paratte G, Lapaire-Cattin M, Lovis C, et al. A walk in the maze: Variation in Late Jurassic tridactyl dinosaur tracks from the Swiss Jura Mountains (NW Switzerland). PeerJ. 2018;6, e4579. https://doi.org/10.7717/peerj.4579 [ Links ]
42. Marty D, Belvedere M, Razzolini NL, Lockley MG, Paratte G, Cattin M, et al. The tracks of giant theropods (Jurabrontes curtedulensis ichnogen. & ichnosp. nov.) from the Late Jurassic of NW Switzerland: Palaeoecological & palaeogeographical implications. Hist Biol. 2018;30(7):928-956. https://doi.org/10.1080/08912963.2017.1324438 [ Links ]
43. Raath MA. The anatomy of the Triassic theropod Syntarsus rhodesiensis (Saurischia, Podokesauridae) and a consideration of its biology [unpublished PhD thesis]. Grahamstown: Rhodes University; 1978. http://hdl.handle.net/10962/d1002051 [ Links ]
44. Bristowe A, Raath MA. A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus. Palaeontol Afr. 2004;40:31-41. [ Links ]
45. Barta DE, Nesbitt SJ, Norell MA. The evolution of the manus of early theropod dinosaurs is characterized by high inter- and intraspecific variation. J Anat. 2018;232(1):80-104. https://doi.org/10.1111/joa.12719 [ Links ]
46. Colbert EH. The Triassic dinosaur Coelophysis. Bulletin no. 57. Flagstaff, AZ: Museum of Northern Arizona; 1989. [ Links ]
47. Paul GS. Are Syntarsus and the Whitaker quarry theropod the same genus? Bull N M Mus Nat Hist Sci. 1993;3:397-102. [ Links ]
48. Gatesy SM, Middleton KM, Jenkins Jr FA, Shubin NH. Three-dimensional preservation of foot movements in Triassic theropod dinosaurs. Nature. 1999;399(6732):141-144. https://doi.org/10.1038/20167 [ Links ]
49. Hattori S. Evolution of the hallux in non-avian theropod dinosaurs. J Vertebr Paleontol. 2016;36(4), e1116995. https://doi.org/10.1080/02724634.2016.1116995 [ Links ]
50. Cashmore DD, Butler RJ. Skeletal completeness of the non-avian theropod dinosaur fossil record. Palaeontology. 2019;62(6):951-981. https://doi.org/10.1111/pala.12436 [ Links ]
 Correspondence:
Correspondence:
Emese Bordy
Email: emese.bordy@uct.ac.za
Received: 01 Nov. 2020
Revised: 09 Dec. 2020
Accepted: 11 Dec. 2020
Published: 28 May 2021
Editor: Margaret Avery
Funding: South African National Research Foundation (grant no. 113394] and DST-NRF Centre of Excellence in Palaeosciences (grant no. CoE PAL 2020)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}