










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.3-4 Pretoria mar./abr. 2021
http://dx.doi.org/10.17159/sajs.2021/8705
RESEARCH ARTICLE
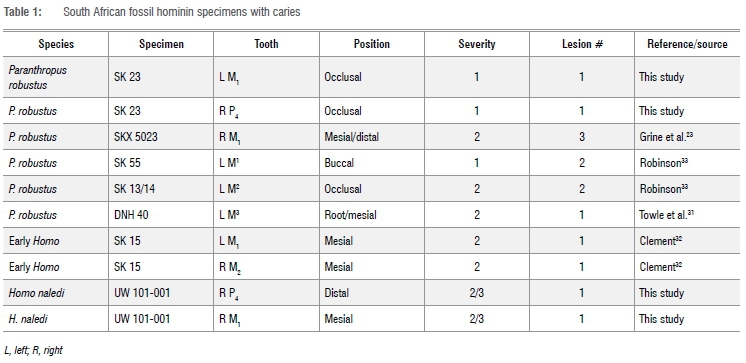
Dental caries in South African fossil hominins
Ian TowleI; Joel D. IrishII, III; Isabelle De GrooteIV; Christianne FernéeV, VI; Carolina LochI
ISir John Walsh Research Institute, Faculty of Dentistry, University of Otago, Dunedin, New Zealand
IIResearch Centre in Evolutionary Anthropology and Palaeoecology, Liverpool John Moores University, Liverpool, United Kingdom
IIIEvolutionary Studies Institute and Centre for Excellence in PaleoSciences, University of the Witwatersrand, Johannesburg, South Africa
IVDepartment of Archaeology, Ghent University, Ghent, Belgium
VDepartment of Anthropology and Archaeology, University of Bristol, Bristol, United Kingdom
VIDepartment of Archaeology, University of Southampton, Southampton, United Kingdom
ABSTRACT
Once considered rare in fossil hominins, caries has recently been reported in several hominin species, requiring a new assessment of this condition during human evolution. Caries prevalence and location on the teeth of South African fossil hominins were observed and compared with published data from other hominin samples. leeth were viewed macroscopically, with lesion position and severity noted and described. For all South African fossil hominin specimens studied to date, a total of 10 carious teeth (14 lesions), including 4 described for the first time here, have been observed. lhese carious teeth were found in a minimum of seven individuals, including five Paranthropus robustus, one early Homo, and one Homo naledi. All 14 lesions affected posterior teeth. The results suggest cariogenic biofilms and foods may have been present in the oral environment of a wide variety of hominins. Caries prevalence in studied fossil hominins is similar to those in pre-agricultural human groups, in which 1-5% of teeth are typically affected.
SIGNIFICANCE:
• This study adds to the growing evidence that dental caries was present throughout the course of human evolution. Caries prevalence in the fossil species studied is similar to those in non-agricultural human groups, with 1-5% of teeth displaying cavities.
• Differences in prevalence and position of dental caries can provide insight into dietary aspects of past populations. South African fossil hominins display lesions on their posterior teeth, suggesting sugary foods were consumed.
Keywords: dental pathology, Homo naledi, Paranthropus robustus, diet, cariogenic bacteria
Introduction
A range of different intraoral bacteria may have been involved in caries formation in the past, including one of the most common forms today, Streptococcus mutans. S. mutans is generally associated with agriculture and population growth. Postindustrial populations, in particular, evidence a marked increase in cariogenic bacteria, as well as less diverse microbiotic ecosystems.1,2 Recent studies suggest the entire microbiota has to be considered in caries research, with a variety of species acting collectively in creating carious lesions.3 Types and proportions of bacteria today differ substantially between active lesions in different locations (e.g. white spot enamel caries vs. deep dentine lesions), as well as between individuals.4 When acids from these bacterial consortiums cannot be neutralised by saliva, lesions form.5 Some foods are especially cariogenic, such as those with high levels of refined carbohydrates and sugars6-8, as well as fruits, honey, and some nuts and seeds.9,10 Tough and fibrous foods are linked with low caries prevalence, as they create a more alkaline oral environment related to substantial saliva production.7,8,11 Diets rich in meat and certain plants have also been associated with low caries prevalence.10,11
Environmental and genetic influences are also important to consider.12,13 It is not yet clear how genetic differences between populations influence caries prevalence13, but interspecific differences in dental morphology will affect lesion position and severity, as is evident when comparing distantly related groups14. Nevertheless, the tooth types, tissues, and tooth areas affected by caries can give further insight into diet and food processing behaviours of a population.10,15,16 Caries lesions have been widely reported in archaeological and fossil samples, with prevalence varying substantially in agricultural groups.10,17-19 Extant nonhuman primates are also reported to have caries, with captive individuals typically more affected than their wild counterparts.20 It is often suggested that caries was scarce in pre-agricultural hominins.21,22 Yet, evidence for such lesions in pre-agricultural groups is growing.23-31
In light of this evidence, we reanalysed South African fossil hominin material and Homo naledi recorded for the first time for the presence of caries. Comparisons of position and prevalence were then made with hominin samples from the literature. Ten carious lesions have already been recorded in South African hominins, including two in a mandible from early Homo, SK 15.32 The others are attributed to Paranthropus robustus, with three lesions on SKX 502323, two on SK 5 533, two on SK 13/1433, and one on DNH 40 from Drimolen31. The aim of this study was to compare the prevalence and characteristics of caries in South African fossil hominins. We hypothesised that there would be differences in caries prevalence and position in the dentition among hominid groups, given substantial dietary differences between great apes, including hominins.34,35
Materials and methods
The South African material that was analysed included specimens assigned to early Homo, Homo naledi, Australopithecus sediba, Australopithecus africanus and Paranthropus robustus, all of which are curated at the University of the Witwatersrand and Ditsong National Museum of Natural History. Only complete teeth were included in the analysis, with each tooth examined under a 10x hand lens for initial lesion classification. A carious lesion was recorded as present when clear enamel cavitation was evident; colour changes alone were not considered diagnostic. Severity was also recorded on a scale of 1 to 4, following Connell and Rauxloh36: (1) enamel destruction only; (2) dentine involvement but pulp chamber not exposed; (3) dentine destruction with pulp chamber exposed; and (4) gross destruction with the crown mostly affected. Lastly, location was scored as distal, buccal, occlusal, lingual, mesial, root, or a combination of these.
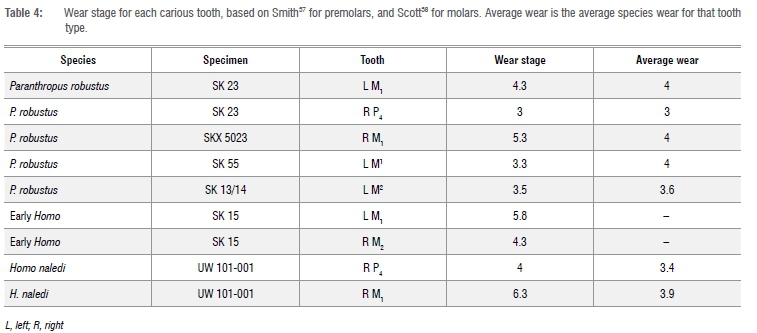
Tooth wear scores were recorded to provide some insight into the interaction with caries, as well as potential information on the role of individual age on lesion formation. Wear was recorded following Smith37 for incisors, canines and premolars, and following Scott38 for molars, as this method allows for more precise inferences on intracrown variability. Molars were split into four quadrants, with each given a wear score of 1 to 10; all other teeth were graded using a single value of 1 to 8. An average for the four molar quadrants was calculated. Like in Meinl et al.16 and Larsen39, no corrective methods were implemented and rates were reported as the per cent of carious teeth among the total number of teeth examined. Comparisons are made between teeth rather than individuals to allow insight into variation across the dentition, but also to maximise data, with fossil samples often represented by loose teeth only. This approach comes with a caveat, as an individual with one carious lesion is more likely to display multiple lesions due to a common oral environment. Thus, a well-preserved individual with multiple lesions would increase caries prevalence, although this potential limitation does not significantly affect the present results.
Micro-computed tomography (CT) scans of specific teeth were taken to clarify if a lesion had been carious. In a clinical setting, scans can differentiate between normal enamel and dentine; areas affected by caries display lower density, which is often identifiable.40 Such techniques can aid in visualisation of a lesion's extent, even if the cavity on the surface is ambiguous.41 Scans were provided by the Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology. They were obtained with a BIR ACTIS 225/300 (settings 100 kV, 100 mA, 0.25 brass filter) or a SkyScan 1172 (settings 100 kV, 94 mA, 2.0 mm aluminum and copper filter) micro-CT scanner. The isometric voxel sizes from these scans range between 15 μηι and 50 μηι (Skinner M 2018, personal communicaton).
Results
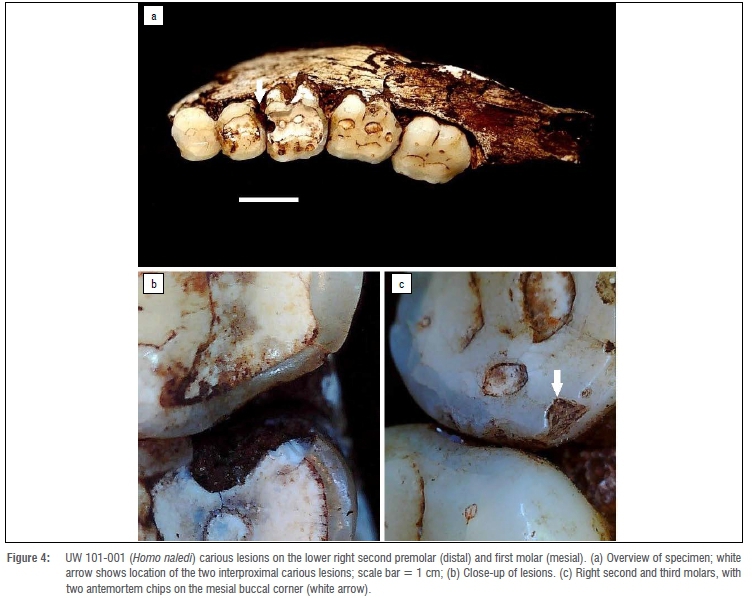
We added four new cases to the six carious teeth already described in the literature for South African hominins (Table 1). Of these four, two are from P. robustus and two are from one H. naledi individual. In total, a minimum of 14 lesions have been diagnosed in all South African material described (Table 1). These include five P. robustus, one H. naledi, and one early Homo individual. None was found in the deciduous teeth, or any tooth from A. africanus and A. sediba. The 14 lesions were observed on 10 teeth from seven individuals. Figures 1-3 highlight lesions previously reported, with more in-depth descriptions of the newly described cases below.
Caries on H. naledi specimen UW 101-001 appear the most severe of all yet described (Figure 4). They seem to have been active for an extended period given penetration deep into the dentine. No difference in crown wear in the antimeres suggests the lesions may not have affected normal mastication. Lesions formed interproximally between the right P4 and adjacent M1, so likely originated from the same microenvironment. Both likely started on the interproximal surface, and enlarged to affect the M1 occlusal surface, with both crown and root surfaces affected. Due to postmortem sediment present on the surface, it is unclear how deep into the dentine these lesions penetrated, or whether they reached the pulp chamber. A micro-CT scan of this specimen was not available at the time of study. Substantial wear and antemortem enamel fractures are also associated with the posterior teeth of this individual.
At least two teeth from P. robustus mandible SK 23 had occlusal caries. The left first molar and right second premolar exhibited larger and darker fissures on their occlusal surface than those present in their antimeres (Figure 5a). However, due to the presence of postmortem matrix in these depressions (Figure 5b), it was difficult to confirm the lesion extent through macroscopic observation. Micro-CT scans could not be used to assess caries presence, as taphonomic processes make the resolution of density differences in the enamel poor. However, enamel under the depression on the premolar occlusal surface appeared to have patches of less dense material, potentially supporting a diagnosis of occlusal caries (Figure 5d, e). There was also a low-density line extending down from the lesion, which was likely a postmortem artefact based on similar features elsewhere in the tooth. The difference in macroscopic appearance compared with its antimere (Figure 5a), large size of the depressions (Figure 5b), and potential demineralisation of enamel (Figure 5d,e), suggest the presence of a carious lesion on the occlusal fissures. These same features were associated with the lesion in the left M1 of SK23.
Caries prevalence for each sample is listed in Tables 2-3. Homo naledi and P. robustus have similar prevalence (1.36% and 1.75% of all permanent teeth analysed, respectively). Early Homo had the highest prevalence of all (4.55%). Australopithecus sediba and A. africanus had no detectable caries lesions, and the former was not included in the Tables due to small sample size. Four of seven carious teeth displayed more occlusal wear than the average for that tooth type (Table 4). Due to small sample size, comparisons for early Homo wear (SK 15) were not possible. However, most carious teeth were near the species wear mean scores (Table 4) and had much of the crown remaining.
Discussion
The results suggest caries may have been more common in past populations than originally thought. In particular, caries is relatively common in several of the South African fossil hominin species, suggesting this pathology may be a common feature of hominins. In addition to macroscopic observations, X-ray imaging, exploration with dental probes and colouration changes are also commonly used to confirm caries lesions in the clinical setting. In archaeological and palaeoanthropological studies, these approaches are often difficult due to postmortem changes (e.g. tooth colouration or structure), or unavailability of diagnostic equipment. Additionally, lesions within interproximal areas may not be observable if teeth remain in situ within the mandible or maxilla. Only a small sample of teeth (n=5) were studied via micro-CT here. A comparative micro-CT study is needed to identify lesions with more accuracy, particularly to account for intersample differences in morphology and taphonomy. Preservation and sample bias are also important considerations, with diagnosis without clear large cavitation not possible in many fossil samples due to taphonomic changes to dental tissues. Therefore, caries prevalence in fossil hominins should be viewed with caution; nevertheless, presence and position can provide useful information concerning diet and behaviour in individuals and groups.
Given the results here and elsewhere23-31, cariogenic bacteria seemed to be prevalent in a number of hominin species. This scenario is supported by research suggesting that a variety of bacteria contribute to a cariogenic microbiota; these same species are often part of the normal oral microbiome in non-pathological situations.3,4 As noted by Sheiham and James42, diet is the primary factor in caries formation and if enough cariogenic foods were consumed regularly by past groups, they would likely develop carious lesions. Differences in microbiota among hominin groups may have influenced caries prevalence, with the threshold required to create a lesion (e.g. regularity of sugar intake) likely varying depending on the prevailing oral microbiota.22,43
High prevalence of occlusal caries is associated with low attrition, while interproximal lesions are linked with high attrition.44 However, interproximal caries may also be related to calculus, as a result of plaque deposition in these areas45, although this link has not been extensively researched. There is no evidence for calculus-associated caries in South African hominins, although postmortem loss of calculus may have occurred. Enamel hypoplasia may also influence caries formation, acting as a site for lesion development because weakened enamel is more vulnerable to acid solubility.8,46 Enamel hypoplasia likely influenced caries formation in P. robustus SK 55 and SK 13/14, with substantial hypoplastic pitting present; pitting is a common feature of this species.47
Other pathologies and wear can also create an environment in which caries is more likely to form, e.g. in response to unusual occlusion patterns.14 It is noteworthy that several fossil teeth with interproximal caries also have substantial cupped occlusal wear and enamel chipping, either above the lesion or on interproximal surfaces of adjacent teeth (SKX 5023, DNH 40 and UW 101-001). Occlusal forces may therefore have been contributing factors, either directly through weakening dental tissues or indirectly through creating an interproximal gap where cariogenic bacteria and foods could accumulate.48
The mastication of hard items (e.g. grit) over time can lead to fatigue of the enamel49,50; as such, certain regions of the crown can become susceptible to pathology. In hominins, the interproximal regions of molars seem susceptible to different types of wear and pathologies, including chipping and non-carious cervical lesions (i.e. 'toothpick' or interproximal grooves). Therefore, higher interproximal stresses may lead to tissue fatigue, which could ultimately mean these regions are more prone to pathology and wear. In humans, caries tends to be more common in posterior teeth, with occlusal surfaces commonly affected.10,17-19,51,52 This pattern is observed in the present hominin sample, with posterior teeth affected more commonly. However, in contrast to recent humans, most lesions were not on occlusal surfaces; this difference likely reflects differences in occlusal wear, in that surface fissures and pits were worn away before lesions could form.
Many human samples from the last 50 000 years show caries frequencies similar to those observed in this study.15,27,39 With the development of agriculture, this frequency varied, and some populations had substantially greater caries rates.10,17-19 It seems that a low to moderate occurrence of caries was common in these hominin populations, typically affecting 1-5% of teeth - as previously suggested for hunter-gatherer groups.53 Lower or higher caries frequencies are usually associated with specific diets or behaviours. For example, high caries prevalence (i.e. >5% of teeth) is associated with agricultural diets10,18,19,51,52, and hunter-gatherer groups reliant on cariogenic foods9,54-56. Low prevalence of caries (i.e. <1% of teeth) is often associated with a diet high in marine foods/ terrestrial meat.15,27,39
It is unlikely that the oral microbiome composition was solely responsible for the lack of carious lesions in A. africanus. Instead, it may relate to dietary differences, or due to sampling bias. The large lesions on P. robustus, H. naledi and early Homo teeth suggest cariogenic foods were consumed. Most fossils assigned to the genus Homo also exhibit caries, including Neanderthals, early Homo, H. erectus, and Pleistocene age H. sapiens25-30,57,58Thus, caries was likely common throughout the evolutionary history of the genus, and may relate to the consumption of cariogenic foods such as tubers, nuts, plants or fruit.9,54-56
A recent study of caries in the African sub-continent through time found that hunter-gatherer groups had a caries prevalence similar to that of the South African fossil hominins analysed here.59 This includes groups relevant to the present study because of geographic proximity, i.e. prehistoric and historic Khoesan peoples. The diet of these hunter-gatherers would have been diverse, but tubers rich in starch have been suggested as one possible contributor to caries occurrence.44 Similarly, honey is consumed in many recent hunter-gatherer groups, thus it may have been present in the diet of early Homo and Paranthropus.11Cooking has also been proposed as a factor influencing caries prevalence25, but there is currently insufficient evidence to conclude if the hominins in this study mastered the use of fire for cooking food. Further ecological/ environmental comparisons may allow insight into the dietary items and food processing techniques facilitating the development of carious lesions. Caries has been present throughout human evolution, with fossil hominins from South Africa affected by this disease. Caries rates observed here are similar to those of pre-agricultural human groups, in which 1-5% of teeth are typically affected.
Acknowledgements
We thank L. Berger and B. Zipfel from the University of the Witwatersrand, and S. Potze from the Ditsong Museum of South Africa for access to their collections. CT scans were provided by the Ditsong Museum, and we thank M. Skinner for kindly providing technical assistance. Research was supported by a studentship to I.T. from Liverpool John Moores University, and a Sir Thomas Kay Sidey Postdoctoral fellowship from the University of Otago.
Competing interests
We declare that there are no competing interests.
Authors' contributions
I.T.: Formulation or evolution of overarching research goals and aims; application of statistical, mathematical, or computational techniques to analyse or synthesise study data. I.T., J.D.I.: Development or design of methodology. I.T., J.D.I., C.F., I.D.G., C.L.: Conducting a research and investigation process; preparation and creation of the published work, specifically writing the initial draft (including substantive translation). I.T., C.F.: Morphological or laboratory assessment of biological samples. J.D.I., I.D.G.: Supervision of student and oversight of study. J.D.I., C.L.: Management and coordination responsibility for the research activity planning and execution. C.F., C.L.: Development or design of methodology.
References
1. Adler CJ, Dobney K, Weyrich LS, Kaidonis J, Walker AW, Haak W, et al. Sequencing ancient calcified dental plaque shows changes in oral microbiota with dietary shifts of the Neolithic and Industrial revolutions. Nat Genet. 2013;45(4):450-455. https://doi.org/10.1038/ng.2536 [ Links ]
2. Cornejo OE, Lefébure T, Pavinski Bitar PD, Lang P Richards VP Eilertson K, et al. Evolutionary and population genomics of the cavity causing bacteria Streptococcus mutans. Mol Biol Evol. 2013;30(4):881-893. https://doi.org/10.1093/molbev/mss278 [ Links ]
3. Simón-Soro A, Mira A. Solving the etiology of dental caries. Trends Microbiol. 2015;23(2):76-82. https://doi.org/10.1016/j.tim.2014.10.010 [ Links ]
4. Aas JA, Griffen AL, Dardis SR, Lee AM, Olsen I, Dewhirst FE, et al. Bacteria of dental caries in primary and permanent teeth in children and young adults. J Clin Microbiol. 2008;46(4):1407-1417. https://doi.org/10.1128/JCM.01410-07 [ Links ]
5. Gussy MG, Waters EG, Walsh O, Kilpatrick NM. Early childhood caries: Current evidence for aetiology and prevention. J Paediatr Child Health. 2006;42(1-2):37-43. https://doi.org/10.1111/j.1440-1754.2006.00777.x [ Links ]
6. Clarkson BH, Krell D, Wefel JS, Crall J, Feagin FF. In vitro caries-like lesion production by Streptococcus mutans and Actinomyces viscosus using sucrose and starch. J Dent Res. 1987;66(3):795-798. https://doi.org/10.1177/00220345870660031801 [ Links ]
7. Prowse TL, Saunders SR, Schwarcz HP, Garnsey P, Macchiarelli R, Bondioli L. Isotopic and dental evidence for infant and young child feeding practices in an imperial Roman skeletal sample. Am J Phys Anthropol. 2008;137(3):294-308. https://doi.org/10.1002/ajpa.20870 [ Links ]
8. Rohnbogner A, Lewis M. Dental caries as a measure of diet, health, and difference in non-adults from urban and rural Roman Britain. Dent Anthropol J. 2016;29(1):16-31. https://doi.org/10.26575/daj.v29i1.32 [ Links ]
9. Humphrey LT, De Groote I, Morales J, Barton N, Collcutt S, Ramsey CB, et al. Earliest evidence for caries and exploitation of starchy plant foods in Pleistocene hunter-gatherers from Morocco. Proc Natl Acad Sci USA. 2014;111(3):954-959. https://doi.org/10.1073/pnas.1318176111 [ Links ]
10. Novak M. Dental health and diet in early medieval Ireland. Arch Oral Biol. 2015;60(9):1299-1309. https://doi.org/10.1016/j.archoralbio.2015.06.004 [ Links ]
11. Moynihan P. Foods and factors that protect against dental caries. Nutr Bull. 2000;25(4):281-286. https://doi.org/10.1046/j.1467-3010.2000.00033.x [ Links ]
12. Slade GD, Sanders AE, Do L, Roberts-Thomson K, Spencer AJ. Effects of fluoridated drinking water on dental caries in Australian adults. J Dent Res. 2013;92(4):376-382. https://doi.org/10.1177/0022034513481190 [ Links ]
13. Haworth S, Shungin D, Van Der Tas JT, Vucic S, Medina-Gomez C, Yakimov V, et al. Consortium-based genome-wide meta-analysis for childhood dental caries traits. Hum Mol Genet. 2018;27(17):3113-3127. https://doi.org/10.1093/hmg/ddy237 [ Links ]
14. Calcagno JM, Gibson KR. Selective compromise: Evolutionary trends and mechanisms in hominid tooth size. In: Kelley MA, Larsen CS, editors. Advances in dental anthropology. New York: Wiley-Liss; 1991. p. 59-76. [ Links ]
15. Kelley MA. Contrasting patterns of dental disease in five early northern Chilean groups. In: Kelley MA, Larsen CS, editors. Advances in dental anthropology. New York: Wiley-Liss; 1991. p. 59-76. [ Links ]
16. Meinl A, Rottensteiner GM, Huber CD, Tangl S, Watzak G, Watzek G. Caries frequency and distribution in an early medieval Avar population from Austria. Oral Dis. 2010;16(1):108-116. https://doi.org/10.1111/j.1601-0825.2009.01624.x [ Links ]
17. Watt ME, Lunt DA, Gilmour WH. Caries prevalence in the permanent dentition of a mediaeval population from the south-west of Scotland. Arch Oral Biol. 1997;42(9):601-620. https://doi.org/10.1016/s0003-9969(97)00061-7 [ Links ]
18. Srejic MD. Dental paleopathology in a Serbian medieval population. Anthropol Anz. 2001;1:113-122. https://doi.org/10.1127/anthranz/59/2001/113 [ Links ]
19. Slaus M, Bedic Z, Rajic Sikanjic P Vodanovic M, Domic Kunic A. Dental health at the transition from the Late Antique to the early Medieval period on Croatia's eastern Adriatic coast. Int J Osteoarchaeol. 2011;21(5):577-590. https://doi.org/10.1002/oa.1163 [ Links ]
20. Crovella S, Ardito G. Frequencies of oral pathologies in a sample of 767 non-human primates. Primates. 1994;35(2):225-230. https://doi.org/10.1007/bf02382058 [ Links ]
21. Tillier AM, Arensburg B, Rak Y Vandermeersch B. Middle Palaeolithic dental caries: New evidence from Kebara (Mount Carmel, Israel). J Hum Evol. 1995;29(2):189-192. https://doi.org/10.1006/jhev.1995.1055 [ Links ]
22. Adler CJ, Browne GV Sukumar S, Hughes T. Evolution of the oral microbiome and dental caries. Curr Oral Health Rep. 2017;1;4(3):264-269. https://doi.org/10.1007/s40496-017-0151-1 [ Links ]
23. Grine FE, Gwinnett AJ, Oaks JH. Early hominid dental pathology: Interproximal caries in 1.5 million-year-old Paranthropus robustus from Swartkrans. Arch Oral Biol. 1990;35(5):381-386. https://doi.org/10.1016/0003-9969(90)90185-d [ Links ]
24. Trinkaus E, Smith RJ, Lebel S. Dental caries in the Aubesier 5 Neandertal primary molar. J Archaeol Sci. 2000;27(11):1017-1021. https://doi.org/10.1006/jasc.1999.0512 [ Links ]
25. Lanfranco LP Eggers S. Caries through time: An anthropological overview. In: Li M-Y editor. Contemporary approach to dental caries. Shangai: Intech Open; 2012. https://doi.org/10.5772/38059 [ Links ]
26. Lacy SA, Wu XJ, Jin CZ, Qin DG, Cai YJ, Trinkaus E. Dentoalveolar paleopathology of the early modern humans from Zhirendong, South China. Int J Paleopathol. 2012;2(1):10-18. https://doi.org/10.1016/j.ijpp.2012.06.003 [ Links ]
27. Lacy SA. Oral health and its implications in late Pleistocene Western Eurasian humans. St. Louis: Washington University; 2014. [ Links ]
28. Liu W, Martinón-Torres M, Cai YJ, Xing S, Tong HW, Pei SW, et al. The earliest unequivocally modern humans in southern China. Nature. 2015;526(7575):696-699. https://doi.org/10.1038/nature15696 [ Links ]
29. Arnaud J, Benazzi S, Romandini M, Livraghi A, Panetta D, Salvadori PA, et al. Neanderthal deciduous human molar with incipient carious infection from the Middle Palaeolithic De Nadale cave, Italy. Am J Phys Anthropol. 2017;162(2):370-376. https://doi.org/10.1002/ajpa.23111 [ Links ]
30. Margvelashvili A, Zollikofer CP Lordkipanidze D, Tafforeau IP Ponce de León MS. Comparative analysis of dentognathic pathologies in the Dmanisi mandibles. Am J Phys Anthropol. 2016;160(2):229-253. https://doi.org/10.1002/ajpa.22966 [ Links ]
31. Towle I, Riga A, Irish JD, Dori I, Menter C, Moggi-Cecchi J. Root caries on a Paranthropus robustus third molar from Drimolen. Am J Phys Anthropol. 2019;170(2):319-323. https://doi.org/10.1002/ajpa.23891 [ Links ]
32. Clement AJ. Caries in the South African apeman: Some examples of undoubted pathological authenticity believed to be 800,000 years old. Brit Dent J. 1956;101:4-7. [ Links ]
33. Robinson JT. Some hominid features of the apeman dentition. J Dent Assoc Series Africa. 1952;7:102-107. [ Links ]
34. Kupczik K, Toro-Ibacache V Macho GA. On the relationship between maxillary molar root shape and jaw kinematics in Australopithecus africanus and Paranthropus robustus. R Soc Open Sci. 2018;5(8):180825. https://doi.org/10.1098/rsos.180825 [ Links ]
35. Towle I, Irish JD, De Groote I. Behavioral inferences from the high levels of dental chipping in Homo naledi. Am J Phys Anthropol. 2017;164(1):184-192. https://doi.org/10.1002/ajpa.23250 [ Links ]
36. Connell B, Rauxloh P. A rapid method for recording human skeletal data. London: Museum of London; 2003. [ Links ]
37. Smith BH. Patterns of molar wear in hunter-gatherers and agriculturalists. Am J Phys Anthropol. 1984;63(1):39-56. https://doi.org/10.1002/ajpa.1330630107 [ Links ]
38. Scott EC. Dental wear scoring technique. Am J Phys Anthropol. 1979;51(2):213-217. https://doi.org/10.1002/ajpa.1330510208 [ Links ]
39. Larsen CS. Dental caries evidence for dietary change: An archaeological context. Adv Dent Anthropol. 1991:179-202. [ Links ]
40. Swain MV Xue J. State of the art of micro-CT applications in dental research. Int J Oral Sci. 2009;1(4):177-188. https://doi.org/10.4248/IJOS09031 [ Links ]
41. Rossi M, Casali F, Romani D, Bondioli L, Macchiarelli R, Rook L. MicroCT scan in paleobiology: Application to the study of dental tissues. Nucl Instrum Methods Phys Res B. 2004;213:747-750. https://doi.org/10.1016/S0168-583X(03)01697-5 [ Links ]
42. Sheiham A, James WP. Diet and dental caries: The pivotal role of free sugars reemphasized. J Dent Res. 2015;94(10):1341-1347. https://doi.org/10.1177/0022034515590377 [ Links ]
43. Simon-Soro A, Tomás I, Cabrera-Rubio R, Catalan MD, Nyvad B, Mira A. Microbial geography of the oral cavity. J Dent Res. 2013;92(7):616-621. https://doi.org/10.1177/0022034513488119 [ Links ]
44. Hillson S. The current state of dental decay. In: Irish JD, Nelson GC, editors. Technique and application in dental anthropology. Cambridge: Cambridge University Press; 2008. p. 53-111. [ Links ]
45. Tomczyk J, Szostek K, Komarnitki I, Mankowska-Pliszka H, Zalewska M. Dental caries and chemical analyses in reconstruction of diet, health and hygienic behaviour in the Middle Euphrates valley (Syria). Arch Oral Biol. 2013;58(6):740-751. https://doi.org/10.1016/j.archoralbio.2012.12.014 [ Links ]
46. Hong L, Levy SM, Warren JJ, Broffitt B. Association between enamel hypoplasia and dental caries in primary second molars: A cohort study. Caries Res. 2009;43(5):345-353. https://doi.org/10.1159/000231571 [ Links ]
47. Towle I, Irish JD. A probable genetic origin for pitting enamel hypoplasia on the molars of Paranthropus robustus. J Hum Evol. 2019;129:54-61. https://doi.org/10.1016/j.jhevol.2019.01.002 [ Links ]
48. Hinton RJ. Differences in interproximal and occlusal tooth wear among prehistoric Tennessee Indians: Implications for masticatory function. Am J Phys Anthropol. 1982;57(1):103-115. https://doi.org/10.1002/ajpa.1330570111 [ Links ]
49. Gao SS, An BB, Yahyazadehfar M, Zhang D, Arola DD. Contact fatigue of human enamel: Experiments, mechanisms and modeling. J Mech Behav Biomed Mater. 2016;60:438-450. https://doi.org/10.1016/j.jmbbm.2016.02.030 [ Links ]
50. Sanchez-Gonzalez E, Pinilla-Cienfuegos E, Borrero-Lopez O, Rodríguez-Rojas F, Guiberteau F. Contact damage of human dental enamel under cyclic axial loading with abrasive particles. J Mech Behav Biomed Mater. 2020;102:103512. https://doi.org/10.1016/j.jmbbm.2019.103512 [ Links ]
51. Varrela TM. Prevalence and distribution of dental caries in a late medieval population in Finland. Arch Oral Biol. 1991;36(8):553-559. https://doi.org/10.1016/0003-9969(91)90104-3 [ Links ]
52. Vodanovic M, Hrvoje B, Mario S, Zeljko D. The frequency and distribution of caries in the mediaeval population of Bijelo Brdo in Croatia (10th-11th century). Arch Oral Biol. 2005;50(7):669-680. https://doi.org/10.1016/j.archoralbio.2004.11.014 [ Links ]
53. Turner CG. Dental anthropological indications of agriculture among the Jomon people of central Japan. X. Peopling of the Pacific. Am J Phys Anthropol. 1979;51(4):619-635. https://doi.org/10.1002/ajpa.1330510413 [ Links ]
54. Walker PL, Hewlett BS. Dental health diet and social status among Central African foragers and farmers. Am Anthropol. 1990;92(2):383-398. https://doi.org/10.1525/aa.1990.92.2.02a00080 [ Links ]
55. Sealy JC, Patrick MK, Morris AG, Alder D. Diet and dental caries among later stone age inhabitants of the Cape Province, South Africa. Am J Phys Anthropol. 1992;88(2):123-134. https://doi.org/10.1002/ajpa.1330880202 [ Links ]
56. Nelson GC, Lukacs JR, Yule P. Dates, caries, and early tooth loss during the Iron Age of Oman. Am J Phys Anthropol. 1999;108(3):333-343. https://doi.org/10.1002/(sici)1096-8644(199903)108:3<333::aid-ajpa8>3.0.co;2-# [ Links ]
57. Brothwell DR. The macroscopic dental pathology of some earlier human populations. In: Dental anthropology. Pergamon Press; 1963. p. 271-288. [ Links ]
58. Clement AJ. The antiquity of caries. Brit Dent J. 1958;104:115-123. [ Links ]
59. Carter F, Irish JD. A sub-continent of caries: Prevalence and severity in early Holocene through recent Africans. Dent Anthropol. 2019;32(2):22-29. https://doi.org/10.26575/daj.v32i2.285 [ Links ]
 Correspondence:
Correspondence:
Ian Towle
Email: ian.towle@otago.ac.nz
Received: 28 July 2020
Revised: 01 Oct. 2020
Accepted: 08 Oct. 2020
Published: 29 Mar. 2021
Editor: Margaret Avery
Funding: Liverpool John Moores University, Sir lhomas Kay Sidey Postdoctoral Fellowship (University of Otago)


{kind=link}
{kind=link}
{kind=link}
{kind=link}