










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.3-4 Pretoria mar./abr. 2021
http://dx.doi.org/10.17159/sajs.2021/7666
RESEARCH ARTICLE
Foliar fungi of the enigmatic desert plant Welwitschia mirabilis show little adaptation to their unique host plant
Martin KemlerI, II, III; Michael J. WingfieldI, II; Don A. CowanI, IV; Bernard SlippersI, II
IDepartment of Biochemistry, Genetics and Microbiology, University of Pretoria, Pretoria, South Africa
IIForestry and Agricultura Biotechnology Institute (FABI). University of Pretoria, Pretoria. South Africa
IIIAG Geobotanik. Ruhr University Bochum, Bochum. Germany
IVCentre for Microbial Ecology anc Genomics, University of Pretoria, Pretoria, South Africa
ABSTRACT
Foliar fungi, especially endophytic fungi, constitute an important part of the microbiome of plants. Yet little is known about the composition of these communities. In this study, we isolated fungi from leaf tissues of the desert plant Welwitschia mirabilis to determine the culturable diversity of the foliar fungal community. The isolated fungal taxa, which grouped into 17 distinct lineages, were identified by sequencing elongation factor 1 alpha, beta-tubulin 1, beta-tubulin 2 and the internal transcribed spacer region. The culturable community was mainly composed of cosmopolitan fungal genera despite the unique taxonomie position of the plant and its geographic isolation. To test for endemism in two of the common fungal genera, Altemaria and Aureobasidium, we built haplotype networks using a global data set. Even this broad data set showed little evidence for specialisation within this unique host or its geographical location. The data suggest that the culturable members of communities of leaf-associated fungi in habitats with little plant coverage, such as the Namib Desert, are mainly established by long-distance aerially distributed fungal inocula and few of these taxa co-evolve with the host within the habitat.
SIGNIFICANCE:
The culturable members of fungal communities associated with an ecological and evolutionary isolated plant have not co-speciated with their hosts, but to a large extent are composed of globally distributed fungal species.
Harsh environmental conditions and the geographic isolation of host plants seem to favour ubiquitous fungal species over more specialist fungal species.
Keywords: Welwitschia mirabilis, Namib Desert, foliar fungi, Altemaria, Aureobasidium
Introduction
Fungi and plants have a long history of co-evolution and plant-fungal interactions are thought to have been essential in the establishment of plants in terrestrial environments. For example, fossil records of interaction structures of arbuscular mycorrhizal fungi and Embryophyta have been recovered from the Rhynie cherts and date back to more than 400 Ma1, possibly as old as 475 Ma2. Structures of endophytic fungi (i.e. microorganisms that in part or during their whole life cycle colonise plant tissues without visible symptoms3) in prostrate axes have been described from as early as 400 Ma.4 Similarly, leaf-associated endophytes are known from the 300 Ma Carboniferous era.5 These studies show that, despite the unknown interaction type between host and fungus, aboveground organs of plants, the so-called phyllosphere, have served as a suitable habitat for fungi for a long time.
Extant phyllosphere-associated fungi including endophytes and epiphytes (i.e. fungi that grow on the surface of plants) mostly show no visible impact on their host under favourable conditions or even enhance plant performance. Some studies have provided evidence that these fungi affect plant physiology. Some epiphytic and endophytic yeasts have been shown to promote plant growth67, whereas some endophytes become pathogenic when the host plant experiences abiotic or biotic stress8,9. Despite our limited knowledge of their function, culture-dependent and culture-independent methods have identified species-rich fungal communities in the phyllosphere that add substantially to the hyperdiversity of fungi.10-14 How these communities become established and the factors that influence them are understood only for a limited number of plant-fungus systems. In the case of endophytes, the greatest impact on the structure of communities results from geographic location and host plant.15 It is also thought that the evolutionary history of the host and the climate16 play additional roles. Most likely similar mechanisms apply for epiphytes.17,18
Plant species in deserts harbour a greater proportion of fungal endophytes with cosmopolitan distribution than the same plant species under less extreme conditions.15 This observation is interesting in the light of two aspects. Firstly, many plant-fungus interactions are characterised by local adaptation to the host population19,20, which is reflected in the genetic sub-structuring of the fungal population21. Secondly, many fungi show clear biogeographic patterns when an appropriate species concept that incorporates molecular data is applied.22 These findings imply that geographic structure should exist in the distribution of cosmopolitan phyllosphere fungi. However, especially culture-dependent studies have recovered many fungal species that seem not necessarily restricted to plant interactions and that have been found in other habitats as well.17,23, 24 Such low host association could counterbalance restrictions to gene flow and lead to low genetic population structure. However, to the best of our knowledge, no studies of phyllosphere-associated fungi have focused on the possibility of biogeographic patterns of cosmopolitan species.
The Namib Desert is one of the oldest and driest deserts on the planet.25 Like all desert ecosystems, it imposes severe constraints on living organisms and is characterised by the stochasticity of nutrient and water supply, as well as continuous varying UV radiation and extreme temperature changes.26 The coastal (western) Namib Desert is unusual in that it experiences frequent fog events with a decreasing gradient from the coast towards the Great Escarpment. It also experiences infrequent and sparse rainfall events with a decreasing gradient from the Great Escarpment towards the coast.25
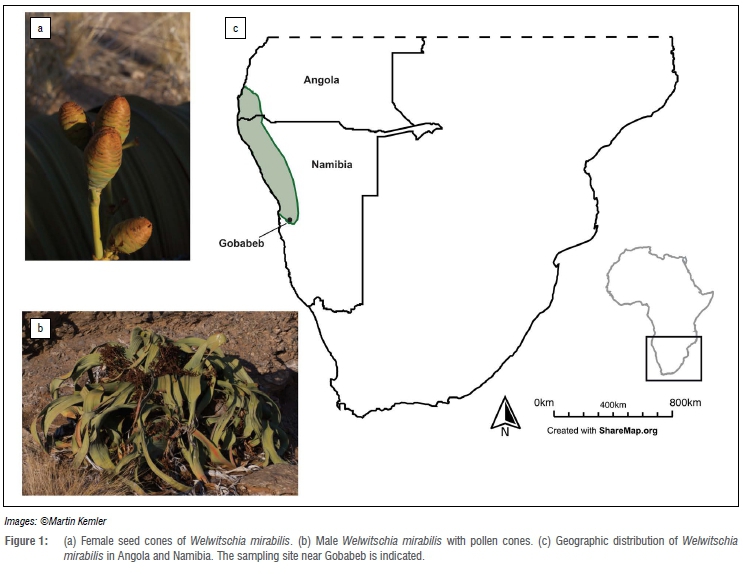
Welwitschia mirabilis Hook, the only living member of the family Welwitschiaceae, is a unique dioecious plant that occurs natively only in the Namib Desert and adjacent savannah ecosystems in Namibia and Angola (Figure 1).27 It is part of an old lineage of land plants, the Gnetophyta, that diverged from the Pinaceae between 121.8 Ma and 309.5 Ma ago.28 Welwitschiaceae fossils have been found from the Lower Cretaceous, 112-114 Ma ago.29 Although these long-lived plants (estimates reach over 2000 years27) have been studied extensively30, little is known regarding their associated mycobiome. A few studies have concentrated on the effect of Aspergillus on seed mortality31,32 and the level of mycorrhizal interactions33, but to date the diversity of phyllosphere fungi associated with Welwitschia is unknown.
The uniqueness of the phylogenetic position of Welwitschia, as well as its remote habitat in one of the oldest and hottest deserts, provides an ideal opportunity to compare factors that might influence the composition of its fungal communities. As a first goal we isolated fungi to assess the diversity and uniqueness of potential endophytes colonising Welwitschia plants at a species level. The second goal was to consider the biogeographic structure of intraspecific diversity for some of the common cosmopolitan fungi that were isolated.
Materials and methods
Samples from Welwitischia mirabilis were recovered under the auspices of the Namibian Ministry of Environment and Tourism permit, number 1522/2011. Sections of leaves (about 20 cm2) were obtained from two individuals in the Welwitschia Wash near Gobabeb (23°36.54' S; 1510.07' E) in the Central Namib Desert in May 2011. Only leaf tissue from young sections near the stem could be obtained, as the plants showed symptoms of intense grazing and lacked any old growth leaf parts. Leaves were washed with ddH20 and subsequently shaken for 1 h in phosphate-buffered saline34 to remove superficially attached spores. Leaves were not surface sterilised, as our interest was in all fungi associated with this desert plant, including endophytic and epiphytic fungi. The leaves were cut into pieces of approximately 4 χ 4 mm and plated on malt extract agar at room temperature (~22 °C). Emerging mycelia were transferred to fresh malt extract agar media to obtain pure cultures.
DNA from pure fungal cultures was isolated using either a salt extraction method described by Aljanabi and Martinez35 or the Qiagen DNeasy® Plant Mini Kit (Qiagen, Hilden, Germany). The fungal barcode region, namely the rDNA ÍTS, as well as parts of elongation factor 1 alpha (EF1a), beta-tubulin 1 (BTUB1) and beta-tubulin 2 (BTUB2) genes for selected samples were amplified (Supplementary table 1). The ITS rRNA gene region was amplified using ITS1F36 and ITS437; EFT a was amplified using EF1f and EF1r; BTUB1 was amplified using BT1a and BT1b38, and BTUB2 was amplified using BT2a and BT2b38. Amplicons were sequenced using the Big Dye™ Terminator Cycle Sequencing Kit V3.1 (Applied Biosystems, Foster City, CA, USA) on an ABI PRISM® 3100 Genetic Analyser (Applied Biosystems, Foster City, CA, USA).
Sequences were checked for quality, trimmed and, where applicable, a consensus sequence was built from forward and reverse strands using CLC Genomics Workbench v4.0.3 (CLC bio, Aarhus, Denmark). Phylogenetic analyses were used to assess the species composition. Phylogenetic trees were inferred for ITS, EF1a and BTUB1 individually, as well as by applying a super matrix approach of a concatenated data set for these DNA regions. For the individual gene trees, sequences were compared against the NCBI nucleotide database using BLASTn39 and the five hits with greatest similarity were downloaded. Alignments were generated using MAFFT v7.154b4041 under the auto option, which chooses the most appropriate alignment strategy for a given data set automatically. The aligned sequences were pasted into the GBIocks web interface42 to remove ambiguous sites, thus allowing smaller final blocks and gaps in the alignment. Species names for the specimens isolated in this study were assigned based on the single tree. For the super matrix approach, selected NCBI sequences of the single gene trees that clustered close to the sequences retrieved in this study were chosen. Sequences were aligned as described for the individual genes and then concatenated using SequenceMatrix43, whereby missing sequences were coded as missing data. Maximum likelihood phylogenies and bootstrap with 1000 replicates were inferred using RAxML 8.0.2544 applied via the raxmlGUI45 using the GAMMAGTR and the rapid bootstrap option.
To consider the possible effects of geographic distribution and adaptation to W. mirabilis, haplotype networks were reconstructed for two of the dominant fungal taxa {Aureobasidium and Alternaria) with a broader phylogenetic sampling from the NCBI database. In the case of Aureobasidium, individual haplotype networks were constructed for ITS and BTUB2, whereas for Alternaria they were constructed for ITS and BTUB1. After aligning the sequences in MAFFT v7.154b, the sequences were collapsed into haplotypes using Map in the SNAP workbench46,47, thus excluding indels and infinite site violations. The pegas package48 in R v3.0.049 was then used to construct minimum spanning trees from these haplotypes.
Results
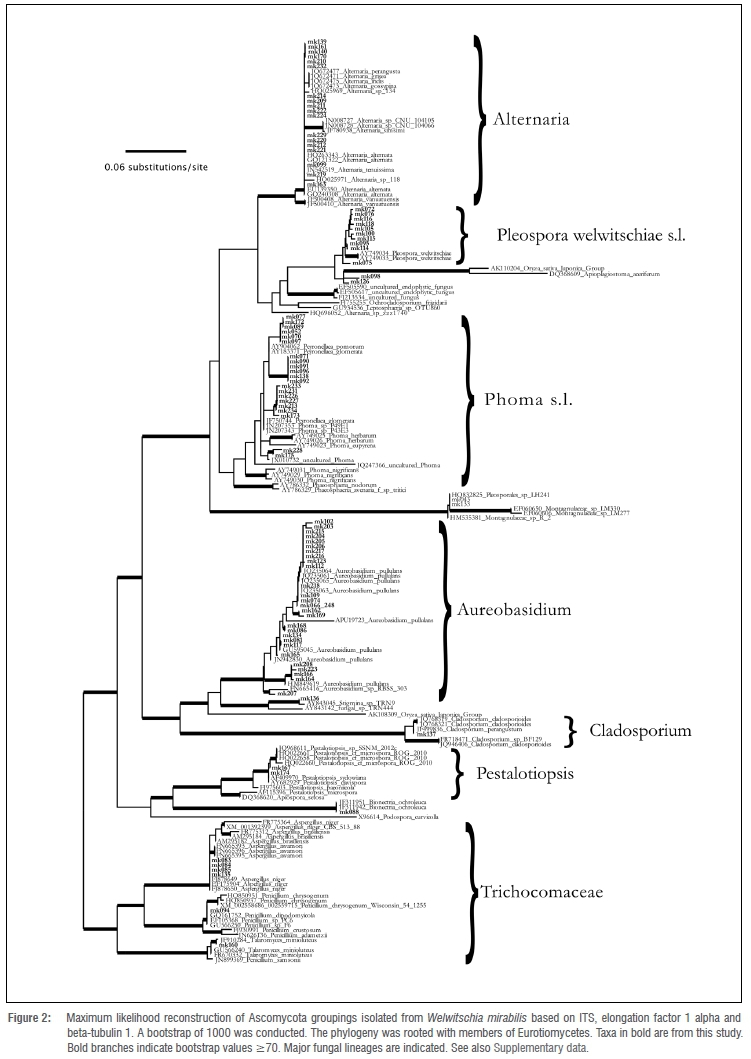
The foliar fungi associated with the leaves of W. mirabilis that could be isolated and grown in culture belonged to either the Ascomycota or Basidiomycota (Supplementary table 1). The latter group was represented by two isolates (Rhodotorula and Cryptococcus) and these were not considered in further analyses. Within the Ascomycota, specimens belonging to the Capnodiales, Dothideales, Eurotiales, Hypocreales, Pleosporales or Xylariales were recovered and the most common taxa were Alternaria, Aspergillus, Aureobasidium, Didymellaceae and Pleospora welwitschiae s.l. Taxa with few isolates recovered belonged to Bionectria, Cladosporium, Montagnulaceae, Penicillium, Pestalotiopsis, Stigmina and Talaromyces (Figure 2). In total, 17 species or clades that might represent species were identified in the Ascomycota. The majority of taxa clustered together with fungal sequences obtained by other studies from other geographic localities. We inferred only three lineages that did not contain sequences from other studies. However, even these lineages clustered closely to sequences from fungi deposited at NCBI (Figure 2). In the Didymellaceae, we recovered one lineage that showed some genetic divergence from its sister taxon Didymella pinodella and another lineage that showed phylogenetic affinity to an uncultured Phoma sequence. One lineage clustered together with Apioplagiostoma aceriferum and a sequence from Oryza sativa (a potential fungal contamination), but with a larger genetic distance and low statistical support. Fungal specimens isolated from Welwitschia that fell within the genera Alternaria and Aureobasidium did not cluster according to their origin but were intermingled with sequences from other geographic origins. The sequences obtained in this study were deposited in GenBank under the accession numbers KT150524-KT150716 (Supplementary table 1).
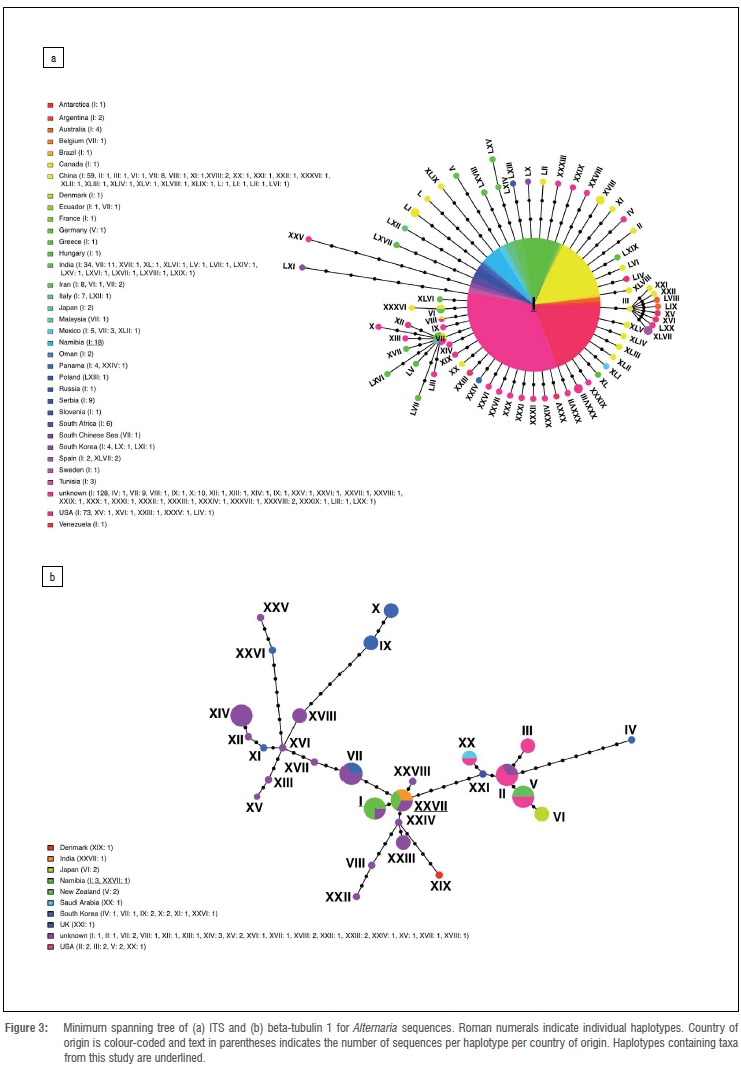
Sequences downloaded from NCBI for the more thorough analyses of geographic clustering of the two most dominant taxa included: for Alternaria, 493 ITS and 122 BTUB1 sequences, and for Aureobasidium, 376 ITS and 50 BTUB2 sequences. The analysis of the ITS sequences for both genera inferred that the dominant haplotypes contained most of the sequences from many different geographic locations (Figure 3a and 4a). The analysis of the BTUB1 region in Alternaria and the analysis of the BTUB2 region in Aureobasidium showed more haplotype diversity. However, there was no clear correlation between geographic location and the occurrence of fungal groupings (Figure 3b and 4b).
Discussion
This study, although restricted to a few leaf samples because of the high protective state of W. mirabilis, revealed a rich fungal diversity of foliar fungi associated with the leaves of this plant. Of the 17 taxa identified, only Pleospora welwitschiae is a species known to be specifically associated with W. mirabilis. Only three other lineages, which according to our analyses could belong to as-yet unknown taxa, might potentially represent endemic or even host-specific lineages. The other taxa clustered closely with sequences of fungi from different geographic origins and appear to have widespread geographic distributions. To understand geographic sub-structuring and potential adaptation we explored haplotype patterns in more detail for the dominant cosmopolitan genera Aureobasidium50and Alternaria51.
In the phylogenetic analysis, only the best matching sequences from NCBI were used for identification purposes, as is common for studies of this nature. However, low taxon sampling in phylogenetic analyses has a limited capacity to show any geographic clustering within a species. Many plant-associated fungi show geographic patterns due to local adaptations to a host population and biogeographic patterns in fungi are more prevalent than previously assumed.2252,53 We thus hypothesised that for adaptation to the locally restricted host plant to occur, some geographic sub-structure must exist in the common fungal taxa associated with W. mirabilis.
In order to identify the existence of geographic sub-structure in the two most frequently isolated fungal taxa isolated from W. mirabilis, haplotype networks using larger sequence data sets with publicly available data were reconstructed. Using these larger data sets, we observed no geographic clustering of our samples of Alternaria and Aureobasidium amongst global collections of closely related sequences (Figures 3 and 4). These results indicate common gene flow between these fungi on W. mirabilis and populations represented by downloaded sequences. They most likely also reflect frequent introduction of these fungi into the Namib Desert. Although grazers (e.g. springbok, donkeys and horses30) could also distribute phyllosphere fungi, given the general isolation of the environments in which W. mirabilis grows, with usually limited human traffic, spores of the commonly isolated and globally distributed fungi are expected to be mostly aerially dispersed. Supporting this view, studies of the biological content of aerosols have shown that fungal biomass can be up to 45% of the total weight of coarse aerosol particles and contain some of the fungi recovered in the present study.54
Isolation of cosmopolitan foliar fungal genera has previously been reported in other plant species15,17,23,24, but seems even more pronounced in individuals in extreme climatic conditions15. Studies of endophytes from desert plants, including other gymnosperms, have shown more diverse communities than in plants of the same species from less extreme environments, but cosmopolitan taxa such as Alternaria, Aspergillus, Aureobasidium, Cladosporium and Phoma constituted the majority of isolated fungi.15,55,56 In Deschampsia antarctica, one of the two angiosperms that grow on the Antarctic continent, endophyte diversity was generally low and the most common endophyte isolated was an Alternaria sp.57 When our results are considered together with these previous studies, it appears that the pattern of high diversity but low uniqueness of plant-associated microbes might be consistent across diverse and isolated extremely dry environments (both cold and hot).
The reason for the high level of occurrence of cosmopolitan and mostly saprobic fungi in harsh environments is not known. It has, however, been hypothesised that infecting any given host in such an environment, even a less optimal host, is preferable to prolonged exposure to extreme temperatures, desiccation or UV radiation.15 An alternative hypothesis, substantiated by analyses in our study, could be that these fungi, due to their cosmopolitan distribution, are not host specific. In this case, they would be outcompeted in environments where specialised fungi occur, thereby decreasing their abundance. In harsh environments where lack of plant cover reduces overall propagule number, as well as opportunities to infect plants, the adaptations of these opportunistic colonists for survival in aerosols and in harsh environments might provide them with a competitive advantage. Some of the common taxa isolated in this study are well adapted to live under desert conditions.
Many of these taxa (e.g. Alternaria58,59, Aspergillus60, Aureobasidium61,62) have been shown to produce melanins, a group of polymers that fungi produce as protection against harsh environmental conditions.63 In cases where the fungi enter an endophytic life stage, the production of melanins might be less relevant. However, during long-distance dispersal, arrival, epiphytic growth, and infection in an extreme environment, melanins could be the difference between success and failure of establishment.
In this study, we used a culture-dependent approach to identify fungi. Studies comparing culture-dependent vs. culture-independent next-generation sequencing studies point to the fact that there could be a significant difference in fungi between these approaches.64 Nevertheless, this culture-based approach remains relevant for the categories of taxa which have been isolated. Future studies, possibly using culture-independent tools and next-generation sequencing, would therefore be especially interesting, particularly with an increased sample size to include gradients from extremely dry to more humid environments. This would make it possible to characterise changes in the patterns common to geographically unique fungal phyllosphere communities. Such studies should be complemented by local air sampling to assess the propagule hypothesis raised in this study.
Acknowledgements
We thank the Department of Science and Technology (DSI)-National Research Foundation (NRF) Centre of Excellence in Tree Health Biotechnology (CTHB) for funding to conduct this study. M.K. thanks the University of Pretoria for postdoctoral funding. We also acknowledge the important improvements suggested by the anonymous reviewers.
Competing interests
We declare that there are no competing interests.
Authors' contributions
D.A.C, B.S. and M.J.W. conceived the study and provided funding. M.K. and D.A.C. conducted the sampling of material. M.K. conducted sample analysis, data analysis and wrote the initial draft. B.S., D.A.C, M.K. and M.J.W. revised the manuscript and wrote the final draft. All authors approved the final draft.
References
1. Remy W, Taylor TN, Hass H, Kerp H. Four hundred-million-year-old vesicular arbuscular mycorrhizae. Proc Natl Acad Sei USA. 1994;91:11841-11843. https://doi.org/10.1073/pnas.91.25.11841 [ Links ]
2. Bidartondo Ml, Read DJ, Trappe JM, Merckx V Ligrone R, Duckett JG. The dawn of symbiosis between plants and fungi. Biol Lett. 2011;7:574-577. https://doi.org/10.1098/rsbl.2010.1203 [ Links ]
3. Wilson D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos. 1995;73:274-276. http://dx.doi.org/10.2307/3545919 [ Links ]
4. Krings M, Taylor TN, Hass H, Kerp H, Dotzier N, Hermsen EJ. Fungal endophytes in a 400-million-yr-old land plant: Infection pathways, spatial distribution, and host responses. New Phytol. 2007;174:648-657. https://doi.org/10.1111/J.1469-8137.2007.02008.x [ Links ]
5. Krings M, Taylor TN, Dotzier N, Galtier J. Fungal remains in cordaite (Cordaitales) leaves from the Upper Pennsylvanian of central France. Bull Geosci. 2011;86:777-784. https://doi.org/10.3140/bull.geosci.1278 [ Links ]
6. Fu SF, Sun PF, Lu HYWei JY Xiao HS, FangWT, etal. Plant growth-promoting traits of yeasts isolated from the phyllosphere and rhizosphere of Drosera spatulata Lab. Fungal Biol. 2016;120:433-448. https://doi.org/10.1016/j.funbio.2015.12.006 [ Links ]
7. Nassar AH, El-Tarabily KA, Sivasithamparam K. Promotion of plant growth by an auxin-producing isolate of the yeast Williopsis saturnus endophytic in maize (lea mays L.) roots. Biol Fertil Soils. 2005;42:97-108. https://doi.org/10.1007/s00374-005-0008-y [ Links ]
8. Sieber TN. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol Rev. 2007;21:75-89. https://doi.org/10.1016/j.fbr.2007.05.004 [ Links ]
9. Slippers B, Wingfield MJ. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol Rev. 2007;21:90-106. https://doi.org/10.1016/j.fbr.2007.06.002 [ Links ]
10. Arnold AE, Maynard Z, Gilbert GS, Coley PD, Kursar TA. Are tropical fungal endophytes hyperdiverse? Ecol Lett. 2000;3:267-274. https://doi.org/10.1046/J.1461-0248.2000.00159.x [ Links ]
11. Fonseca Á, Inácio J. Phylloplane yeasts. In: Rosa C, Gábor Ρ, editors. Biodiversity and ecophysiology of yeasts. Berlin/Heidelberg: Springer; 2006. p. 263-301. https://doi.org/10.1007/3-540-30985-3_13 [ Links ]
12. Jumpponen A, Jones KL. Massively parallel 454 sequencing indicates hyperdiverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol. 2009;184:438-448. https://doi.org/10.1111/j.1469-8137.2009.02990.x [ Links ]
13. Kemler M, Garnas J, Wingfield MJ, Gryzenhout M, Pillay K-Α, Slippers B. Ion torrent PGM as tool for fungal community analysis: A case study of endophytes in Eucalyptus grandis reveals high taxonomie diversity. PLoS ONE.2013;8, e81718. https://doi.org/10.1371/journal.pone.0081718 [ Links ]
14. Zimmerman NB, Vitousek PM. Fungal endophyte communities reflect environmental structuring across a Hawaiian landscape. Proc Natl Acad Sei USA. 2012;109:13022-13027. https://doi.org/10.1073/pnas.1209872109 [ Links ]
15. Hoffman MT, Arnold AE. Geographic locality and host identity shape fungal endophyte communities in cupressaceous trees. Mycol Res. 2008;112:331- 344. https://doi.org/10.1016/j.mycres.2007.10.014 [ Links ]
16. U'Ren JM, Lutzoni F, Miadlikowska J, Laetsch AD, Arnold AE. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am J Bot. 2012;99:898-914. https://doi.org/10.3732/ajb.1100459 [ Links ]
17. Guimaraes JB, Pereira P, Chambel L, Tenreiro R. Assessment of filamentous fungal diversity using classic and molecular approaches: Case study -Mediterranean ecosystem. Fungal Ecol. 2011;4:309-321. https://doi.org/10.1016/j.funeco.2011.01.006 [ Links ]
18. Kemler M, Witfeld F Begerow D, Yurkov A. Phylloplane yeasts in temperate climates. In: Buzzini P, Lachance MA, Mjrkov A, editors. Yeasts in natural ecosystems: Diversity. Cham: Springer Nature; 2017. p. 171-197. https://doi.org/10.1007/978-3-319-62683-3_6 [ Links ]
19. Greischar MA, Koskella B. A synthesis of experimental work on parasite local adaptation. Ecol Lett. 2007;10:418-434. https://doi.Org/10.1111/Í-1461-0248.2007.01028.x [ Links ]
20. Kaitz O, Shykoff JA. Local adaptation in host-parasite systems. Heredity. 1998;81:361-370. https://doi.org/10.1046/jl365-2540.1998.00435.x [ Links ]
21. Enjalbert J, Duan X, Leconte M, Hovmoller MS, De Vallavieille-Pope C. Genetic evidence of local adaptation of wheat yellow rust (Puccinia striiformis f. sp. tritici) within France. Mol Ecol. 2005;14:2065-2073. https://doi.org/10.1111/J.1365-294X.2005.02566.X [ Links ]
22. Taylor JW, Turner E, Townsend JP, Dettman JR, Jacobson D. Eukaryotic microbes, species recognition and the geographic limits of species: Examples from the kingdom Fungi. Philos Trans R Soc Lond Β Biol Sei. 2006;361:19471963. https://doi.org/10.1098/rstb.2006.1923 [ Links ]
23. Smith H, Wingfield MJ, Petrini 0. Botryosphaeria dothidea endophytic in Eucalyptus grandis and Eucalyptus nitens in South Africa. For Ecol Manag. 1996;89:189-195. https://doi.org/10.1016/S0378-1127(96)03847-9 [ Links ]
24. Unterseher Μ, Schnittler Μ. Species richness analysis and ITS rDNA phylogeny revealed the majority of cultivable foliar endophytes from beech Fagus sylvatica. Fungal Ecol. 2010;3:366-378. https://doi.0rg/10.1016/j.funeco.2010.03.001 [ Links ]
25. Viles HA, Goudie AS. Weathering in the central Namib Desert, Namibia: Controls, processes and implications. J Arid Environ. 2011;93:20-29. https://doi.org/10.1016/j.jaridenv.2011.09.012 [ Links ]
26. Noy-Meir I. Desert ecosystems: Environment and producers. Annu Rev Ecol Syst. 1973;4:25-51. https://doi.org/10.1146/annurev.es.04.110173.000325 [ Links ]
27. Von Willert DJ. Welwitschia mirabilis Hook. fil. - das Überlebenswunder der Namibwüste [Welwitschia mirabilis Hook. fil. - the survival wonder of the Namib Desert]. Naturwissenschaften. 1994;81:430-442. German, https://doi.org/10.1007/BF01136642 [ Links ]
28. Clarke JT, Warnock RC, Donoghue PC. Establishing a time-scale for plant evolution. New Phytol. 2011;192:266-301. https://doi.org/10.1111/j1469-8137.2011.03794.x [ Links ]
29. Dilcher DL, Bernardes-De-Oliveira ME, Pons D, Lott TA. Welwitschiaceae from the Lower Cretaceous of northeastern Brazil. Am J Bot. 2005;92:1294-1310. https://doi.Org/10.3732/ajb.92.8.1294 [ Links ]
30. Henschel JR, Seely MK. Long-term growth patterns of Welwitschia mirabilis, a long-lived plant of the Namib Desert (including a bibliography). Plant Ecol. 2000;150:7-26. https://doi.Org/10.1023/A:1026512608982 [ Links ]
31. Pekarek E, Jacobson Κ, Donovan A. High levels of genetic variation exist in Aspergillus niger populations infecting Welwitschia mirabilis Hook. J Hered. 2006;97:270-278. https://doi.org/10.1093/jhered/esj031 [ Links ]
32. Whitaker C, Pammenter NW, Berjak Ρ Infection of the cones and seeds of Welwitschia mirabilis by Aspergillus niger var. phoenicis in the Namib-Naukluft Park. S Afr J Bot. 2008;74:41-50. https://doi.Org/10.1016/j.sajb.2007.08.008 [ Links ]
33. Jacobson KM, Jacobson PJ, Miller Jr OK. The mycorrhizal status of Welwitschia mirabilis. Mycorrhiza. 1993;3:13-17. https://doi.org/10.1007/BF00213462 [ Links ]
34. Mendes R, Pizzirani-Kleiner AA, Araujo WL, Raaijmakers JM. Diversity of cultivated endophytic bacteria from sugarcane: Genetic and biochemical characterization of Burkholderia cepacia complex isolates. Appl Environ Microbiol. 2007;73:7259-7267. https://doi.org/10.1128/AEM.01222-07 [ Links ]
35. Aljanabi SM, Martinez I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997;25:4692-4693. https://doi.Org/10.1093/nar/25.22.4692 [ Links ]
36. Gardes M, Bruns TD. ITS primers with enhanced specificityforbasidiomycetes - application to the identification of mycorrhizae and rusts. Mol Ecol. 1993;2:113-118. https://doi.Org/10.1111/j.1365-294X.1993.tb00005.x [ Links ]
37. White TJ, Bruns Τ, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis M, Gelfand D, Shinsky J, White Τ, editors. PCR protocols: A guide to methods and applications. San Diego, CA: Academic Press; 1990. p. 315-322. https://doi.org/10.1016/B978-0-12-372180-8.50042-1 [ Links ]
38. Glass NL, Donaldson GC. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl Environ Microbiol. 1995;61:1323-1330. [ Links ]
39. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403-410. https://doi.org/10.1016/S0022-2836(05)80360-2 [ Links ]
40. Katoh K, Misawa K, Kuma K, Miyata T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002;30:3059-3066. https://doi.org/10.1093/nar/gkf436 [ Links ]
41. Katoh K, Kuma K, Toh H, Miyata T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005;33:511-518. https://doi.Org/10.1093/nar/gki198 [ Links ]
42. Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. 2000;17:540-552. https://doi.org/10.1093/oxfordjournals.molbev.a026334 [ Links ]
43. Vaidya G, Lohman DJ, Meier R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics. 2011;27:171-180. https://doi.Org/10.1111/j.1096-0031.2010.00329.x [ Links ]
44. Stamatakis A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312-1313. https://doi.org/10.1093/bioinformatics/btu033 [ Links ]
45. Silvestro D, Michalak I. raxmlGUI: A graphical front-end for RAxML. Org Divers Evol. 2012;12:335-337. https://doi.org/10.1007/s13127-011 -0056-0 [ Links ]
46. Aylor DL, Price EW, Carbone I. SNAP: Combine and Map modules for multilocus population genetic analysis. Bioinformatics. 2006;22:1399-1401. https://doi.org/10.1093/bioinformatics/btl136 [ Links ]
47. Price EW, Carbone I. SNAP: Workbench management tool for evolutionary population genetic analysis. Bioinformatics. 2005;21:402-404. https://doi.org/10.1093/bioinformatics/bti003 [ Links ]
48. Paradis Ε. pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics. 2010;26:419-420. https://doi.org/10.1093/bioinformatics/btp696 [ Links ]
49. R Core Team. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2016. [ Links ]
50. Zalar P, Gostincar C, De Hoog GS, Ursic V Sudhadham Μ, Gunde-Cimerman Ν. Redefinition of Aureobasidium pullulans and its varieties. Stud Mycol. 2008;61:21 -38. https://doi.org/10.3114/sim.2008.61.02 [ Links ]
51. Woudenberg JHC, Groenewald JZ, Binder M, Crous PW. Altemaria redefined. Stud Mycol. 2013;75:171-212. https://doi.org/10.3114/sim0015 [ Links ]
52. Ellison CE, Hall C, Kowbel D, Welch J, Brem RB, Glass NL, et al. Population genomics and local adaptation in wild isolates of a model microbial eukaryote. Proc Natl Acad Sei USA. 2011;108:2831-2836. https://doi.org/10.1073/pnas.1014971108 [ Links ]
53. Geml J, Tulloss RE, Laursen GA, Sazanova NA, Taylor DL. Evidence for strong inter- and intracontinental phylogeographic structure in Amanita muscaria, a wind-dispersed ectomycorrhizal basidiomycete. Mol Phylogenet Evol. 2008;48:694-701. https://doi.org/10.1016/j.ympev.2008.04.029 [ Links ]
54. Elbert W Taylor PE, Andreae MO, Poschl U. Contribution of fungi to primary biogenic aerosols in the atmosphere: Wet and dry discharged spores, carbohydrates, and inorganic ions. Atmos Chem Phys Discuss. 2007;7:4569-5588. https://doi.org/10.5194/acp-7-4569-2007 [ Links ]
55. Sun X Wang Q, Lu X, Okane I, Kakishima M. Endophytic fungal community in stems and leaves of plants from desert areas in China. Mycol Prog. 2012;11:781 -790. https://doi.org/10.1007/s11557-011-0790-x [ Links ]
56. Suryanarayanan TS, Wittlinger SK, Faeth SH. Endophytic fungi associated with cacti in Arizona. Mycol Res. 2005;109:635-639. https://doi.org/10.1017/S0953756205002753 [ Links ]
57. Rosa LH, Vaz ABM, Caligiorne RB, Campolina S, Rosa CA. Endophytic fungi associated with the Antarctic grass Deschampsia antarctica Desv. (Poaceae). Polar Biol. 2009;32:161-167. https://doi.org/10.1007/s00300-008-0515-z [ Links ]
58. Kimura N, Tsuge T. Gene cluster involved in melanin biosynthesis of the filamentous fungus Alternaria alternata. J Bacteriol. 1993;175:4427-4435. https://doi.org/10.1128/jb.175.14.4427-4435.1993 [ Links ]
59. Pridham JB, Woodhead S. The biosynthesis of melanin in Alternaria. Phyto-chemistry. 1977;16:903-906. https://doi.org/10.1016/S0031-9422(00)86689-3 [ Links ]
60. Youngchim S, Morris-Jones R, Hay RJ, Hamilton AJ. Production of melanin by Aspergillus fumigatus. J Med Microbiol. 2004;53:175-181. https://doi.org/10.1099/jmm.0.05421-0 [ Links ]
61. Durreil LW. Studies of Aureobasidium pullulans (de Bary) Arnaud. Mycopathologia et Mycologia Applicata. 1968;35:113-120. https://doi.org/10.1007/BF02049574 [ Links ]
62. Gadd GM. Melanin production and differentiation in batch cultures of the polymorphic fungus Aureobasidium pullulans. FEMS Microbiol Lett. 1980;9:237-240. https://doi.Org/10.1111/j.1574-6968.1980.tb05644.x [ Links ]
63. Gessler NN, Egorova AS, Belozerskaia TA. Melanin pigments of fungi under extreme environmental conditions (Review). Appl Biochem Microbiol. 2014;50:105-113. https://doi.org/10.1134/S0003683814020094 [ Links ]
64. Siddique AB, Khokon AM, Unterseher Μ. What do we learn from cultures in the omics age? High-throughput sequencing and cultivation of leaf-inhabiting endophytes from beech (Fagus sylvatica L.) revealed complementary community composition but similar correlations with local habitat conditions. MycoKeys. 2017;20:1-16. https://doi.org/10.3897/mycokeys.20.11265 [ Links ]
 Correspondence:
Correspondence:
Martin Kemler
Email: martin.kemler@rub.de
Received: 25 Nov. 2019
Revised: 28 May 2020
Accepted: 17 Aug. 2020
Published: 29 Mar. 2021
Editors: Teresa Coutinho, Salmina Mokgehle
Funding: Department of Science and Innovation (DSI)-National Research Foundation (NRF) Centre of Excellence in Tree Health Biotechnology (CTHB). University of Pretoria


{kind=link}
{kind=link}
{kind=link}
{kind=link}