










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.1-2 Pretoria ene./feb. 2021
http://dx.doi.org/10.17159/sajs.2021/7137
RESEARCH ARTICLE
The cryptic case of Otomys sloggetti (Sloggett's vlei rat): Interpreting murid molar morphology in the fossil record
Thalassa MatthewsI; Turid H. NelII
ICentre of Excellence in Palaeosciences, Iziko Museums of South Africa, Cape Town, South Africa
IISFF Centre for Early Sapiens Behaviour (SapienCE), University of Bergen, Bergen, Norway
ABSTRACT
Vlei rats (Family: Muridae; Subfamily: Otomyinae) have a widespread distribution in southern Africa. They are favoured prey of barn and spotted eagle owls, and frequently become associated with archaeological deposits when the owls roost in cave sites. The phylogeny of several Otomyinae species is enigmatic, and Otomys sloggetti (Sloggett's vlei rat) is no exception. This species has been referred to as the 'ice rat' and present distribution ranges are seemingly limited to mountainous areas, at high altitude, in Lesotho, Drakensberg and the Karoo. It was thus surprising and unexpected when specimens closely resembling Otomys sloggetti (identification was based on molar morphology) were found in several archaeological sites on the south and west coasts of South Africa, and also in modern owl pellet assemblages - all extralimital to the current reported distribution. However, further examination of and comparison between these specimens, as well as extensive differences observed between comparative Otomys sloggetti specimens from museum collections, highlighted potential problems associated with the common practice of using tooth morphology to identify fossil murid species. We identified six molar morphotypes from the fossil and modern material, all of which bore a morphological resemblance to O. sloggetti. The material discussed in this paper suggests that cryptic, undescribed vlei rat species, or subspecies, have been in the past, and may yet be, co-occurring with modern populations of O. karoensis and O. irroratus. Phylogenetic studies need to be done in conjunction with morphological studies, as, currently, the relationship between the huge variation seen in interspecific morphology with genetics is little understood, different Otomys species are not always distinguishable morphologically, and considerable chromosomal polytypes have been found. Our findings highlight the need for extensive cladistic and genetic research on the Otomyinae.
SIGNIFICANCE:
• Mice and shrews from fossil sites are frequently used by archaeologists as indicators of past climatic and environmental conditions. Research into the species present in fossil assemblages is usually done on a single site basis and intersite comparisons are rare. The taxonomic conundrums presented by a vlei rat found in several South African archaeological sites indicates that such comparisons could result in the re-evaluation of identifications, and/or indicate the presence of cryptic species/subspecies. Phylogenetic studies are needed in conjunction with morphological studies, as the relationship between variations in interspecific tooth morphology (used to identify taxa) with genetics is little understood. This in turn will help to elucidate the relationship between morphology, biogeography and local adaptations.
Keywords: Otomyinae, Pinnacle Point, Klipdrift Shelter, tooth morphology
Introduction
The identity, and biogeographical affiliations, of fossil rats and mice (Family: Muridae) are important as these small animals, which have short lifespans, small ranges, and in some cases particular habitat requirements, are used to provide environmental information in order to reconstruct ancient environments, and trace palaeoenvironmental change over time. The correct identification of these animals in fossil sites is thus important as errors can lead to erroneous assumptions and conclusions being made about past environments, and the effects of climate change. Here we focus on a vlei rat species, namely Otomys sloggetti, and use the fossil record of this taxon on the south coast of South Africa to illustrate some of the issues and potential problems associated with the common practice of using tooth morphology to identify fossil murid species.
Research into the murid species present in fossil assemblages is usually done on a single site basis, and it is extremely rare for researchers to compare the same taxon across sites. The taxonomic conundrums presented in this paper indicate that such comparisons might result in the re-evaluation of identifications, as inconsistencies and errors may ensue if museum specimens have been incorrectly identified - a scenario which is particularly likely in the case of cryptic species (species which look morphologically the same, but belong to different species). Cross-site comparisons, such as those made here, may also indicate variability within a species suggestive of misidentification, and/or the presence of cryptic species. We compare the molar morphology of a variety of fossil Otomys cf. sloggetti from a number of south coast archaeological sites, and a west coast site, with each other, with modern Otomys sloggetti, and with two other Otomys species (O. irroratus and O. karoensis). These comparisons include comparative material from museum collections, and from two south coast barn owl pellet collections.
Taxonomy and phylogeny of Otomys sloggetti
Otomys sloggetti is a rat which belongs to the relatively speciose 'vlei rat' (also called 'laminate toothed rat') tribe. These herbivorous short-tailed rodents (Subfamily: Murinae) are frequently mentioned as being 'volelike' in appearance. The Otomyini are represented by some 32 recognised species (including two of which are undescribed) and are endemic and widespread in sub-Saharan Africa, with a number of species occurring in South Africa.1 The exact number of extant Otomys species is contentious and a number of cryptic species have been identified, while others await further elucidation.2-5 Some studies suggest that speciation has occurred along biomes, with phenotypic and genotypic divergent lineages corresponding to the Fynbos/Albany Thicket and Grassland biomes in the case of Otomys irroratus and Otomys karoensis.2,3Geometric morphometric analyses carried out on the crania of Otomys unisulcatus also failed to support the genetic groupings, but rather followed biome boundaries, indicating previous environmental adaptations.6 This, together with the adaptability observed in fossil Otomys communities when faced with environmental and climatic change on the south coast of South Africa7, as well as the fact that up to three or more Otomys species may live sympatrically in an area1,2,7, indicates the flexibility and plasticity of this murid subfamily.
The phylogeny of Otomys sloggetti, and the number of lineages and species present, as is the case with many of the other otomyine species, is currently unresolved.1,2 In a study that looked at mitochondrial and nuclear genes, as well as morphological characteristics, specimens previously identified as Otomys sloggetti were found to represent two clades, and three lineages.8 A cladistic analysis of 45 morphological (craniodental) and 46 binary allozyme characters indicated that Otomys sloggetti was basal to a 'mesic clade' of southern and eastern African species9, and the idea that Otomys sloggetti represents a basal lineage has been reiterated elsewhere10. The most recent research into the phylogeny of O. sloggetti by Taylor et al.2 differentiated three distinct mtDNA clades of O. sloggetti in the Drakensberg, Maluti and Sneeuberg ranges, respectively, extending the range of this species to beyond the Sneeuberg and the southern Great Escarpment. Further morphometric and karotypic analyses are required to elucidate the extent to which these species are cryptic.2 The five laminae observed in the upper third molar of Otomys sloggetti, and also in the extinct fossil species Otomys gracilis, are noted as being a feature of earlier forms of the genus, and it is suggested that younger (more recent) taxa, such as O. irroratus, O. karoensis and O. laminatus, have more laminae.11 Previously, O. sloggetti and O. unisulcatus were placed in a separate family (Afyotomys)12, but recent genetic research has resulted in them being reclassified as Otomys1,2. Fossil Otomys sloggetti have been found in a few Cradle of Humankind fossil sites in Gauteng Province, including Gladysvale Cave13, Border Cave14 (where cf. O. sloggetti was tentatively identified) and Sterkfontein Cave15, which date as far back as 3.3 Ma16.
Small mammals as palaeoenvironmental indicators
As mentioned previously, archaeologists and palaeontologists frequently use rats, mice and shrews from fossil sites as indicators of past climatic and environmental conditions.17-19 These animals generally become associated with fossil deposits when barn and eagle owls, which roost in the same caves occupied by people, regurgitate the bones and teeth of their prey in their pellets. Over time, the pellets disintegrate, leaving lenses of bones and teeth in the sediment. As these owls have small home ranges, and select a broad range of prey, they provide a good sample of the small mammal species living in the vicinity of the cave site.14,17 The teeth of rodents and soricids (shrews) are generally used to identify the small mammal species present in archaeological deposits. The laminate molars of vlei rats, and the terminology used in this paper, is illustrated in Figure 1.
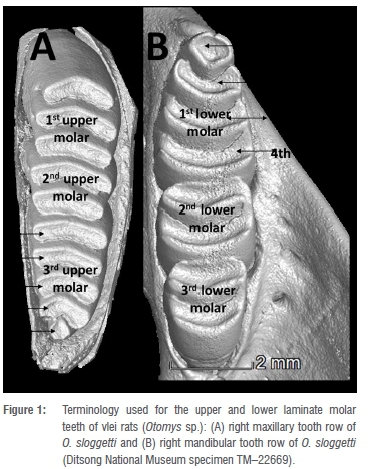
Vlei rats have laminate teeth and generally show a high degree of intraspecific variability in terms of molar shape, and sometimes even in the number of laminae. For example, O. karoensis is noted as sometimes having five, rather than six, laminae on the upper third molar.1 This variability, which includes biogeographical differences in morphology and size, and the fact that the appearance of the occlusal surface of the tooth changes over time with wear (in some species wear leads to the joining of previously separate laminae), makes this a difficult group of rodents to identify. In fossil deposits, the cranial bones are frequently broken to the degree that only isolated molars are recovered. This means that the approach of identifying features used with modern comparative material, such as presence/absence of grooves on the lower incisor, and the shape of the petrotympanic foramen, cannot be used. O. sloggetti is distinguishable from all other Otomys species in having ungrooved lower incisors, and a slit-shaped petrotympanic foramen.2,9 In the case of material from fossil deposits, however, the lower first molar (M1) of the mandible, and upper third molar (M3) of the maxilla (see Figure 1), are the most distinctive and useful teeth for the identification of Otomys species.
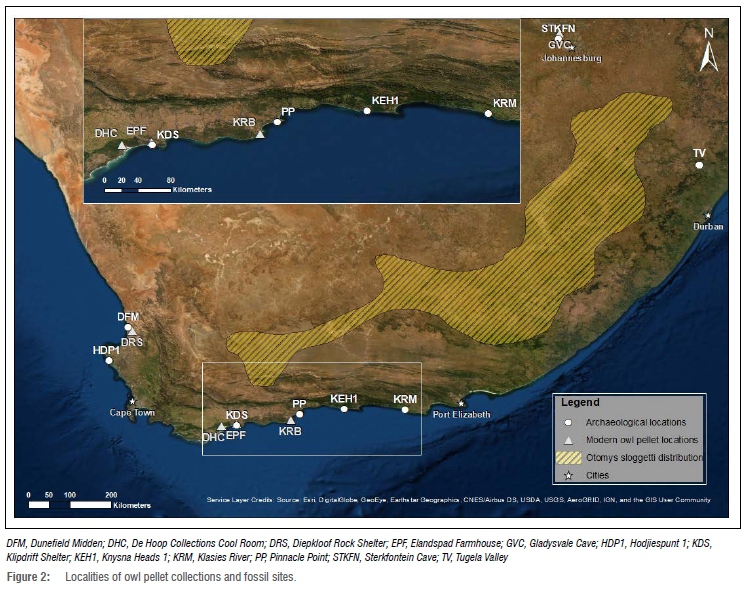
The fossil specimens referred to in this article are currently curated at the Iziko South African Museum and come from a number of archaeological sites including the Pinnacle Point complex (PP), Knysna Heads 1 (KEH1), Dunefield Midden (DFM), Sterkfontein Cave (STKFN), Klasies River main site (KR) and Klipdrift Shelter (KDS) (see Figure 2 for these localities). Comparative, modern material was obtained from the Ditsong National Museum of Natural History, as well as Iziko. Modern Otomys specimens were recovered from owl pellet assemblages collected at Elandspad Farmhouse (EPF) and De Hoop Collections Cool Room (DHC) in the De Hoop Nature Reserve, Keurbooms (KRB) in the Wilderness area, and Diepkloof Rock Shelter (DRS) on the west coast. Figure 2 shows the fossil and pellet collection sites mentioned in this paper, and the current extent of O. sloggetti's distribution based on the IUCN Red List.20 Computed tomography (CT) scans of individual fossils, and some of the modern material, were carried out at the Central Analytical Facilities of Stellenbosch University (South Africa), using their micro- and nano-CT scanning facilities. Length and breadth measurements (see Appendix 1) of all M1 and M3 molars were made on nano-CT scans of the specimens, using the 'measuring' and 'dimensions' functions of VGMax, version 3.3. Measurements of all the teeth presented here are given in Appendix 1. Note that these must be considered within the context of age and wear patterns, as older individuals, with more worn teeth, will automatically yield higher measurements than younger individuals, as the occlusal surface becomes longer, and broader, with wear. Photographs of modern specimens from EPF were taken with a Leica M205A camera attached to a Leica M275 microscope. Specimens from KDS were photographed with a Leica DFC 295 camera attached to a Leica M125 C microscope.
Otomys sloggetti and the south coast fossil record
Otomys sloggetti sensu lato frequently co-occurs with two other Otomys species, namely Otomys karoensis and Otomys irroratus, in fossil deposits on the south coast.7O. sloggetti sensu lato was identified in the fossil deposits based on the M3 and M1, and is differentiated by two specific molar morphologies. Firstly, by the presence of five laminae on the upper third molar (M3) as opposed to the six laminae generally found in Otomys karoensis and Otomys irroratus, and secondly, a rounded anterior first laminae (M1) (which differs to the rectangular first lamina of the other two species), and in the distribution of the laminae across the occlusal surface of the molar. Another vlei rat with the same general morphology of the M1 and M3, O. unisulcatus, has also made an extralimital appearance in some south coast fossil horizons, but the shape and general appearance of the molars are differentiate from that of O. sloggetti (see Figures 5 and 7) as the M3 has generally smaller dimensions and is roughly rectangular in shape, and the anterior and second laminae of the M1 merge to form a characteristic 'C' shape, with only slight wear.




The appearance of O. sloggetti in a number of fossil south coast South African sites (see Figure 2 for localities) was unexpected, as it has been called a high altitude and alpine endemic species, and its current distribution includes the Drakensberg, Sneeuberg and the southern Great Escarpment2, with a couple of isolated populations on mountains in the Karoo8,21-23. This species is often referred to as the 'ice rat' due to its appearance in areas which reach icy temperatures. The appearance of O. sloggetti in the south coast fossil record was surprising as it appeared to belie the categorisation of this taxon as being a 'high altitude endemic' and 'montane-adapted'9. Otomys sloggetti was identified in
three coastal cave sites situated on Pinnacle Point, just to the southwest of Mossel Bay (PP9C, PP30 and PP13B), as well as in another coastal archaeological cave site situated north of the Pinnacle Point sites on the eastern Knysna Head (KEH1).7 At KEH1, O. sloggetti was identified in horizons dating to around the Last Glacial Maximum (~23 ka), and in the Pinnacle Point sites sometime during the period 157-180 ka (PP13B), at ~150 ka (PP30), and at ~130 ka (PP9C).7 Due to an observed high degree of variation between fossil specimens, existing phylogenetic uncertainties, and variation between modern O. sloggetti teeth obtained from comparative museum collections, identification of the fossil taxa was later changed to O. cf. sloggetti sensu lato. The appearance of O. cf. sloggetti in surface deposits in PP9C and PP13C was suggested to have resulted from the mixing of surface and fossil deposits, and/ or be an indication of the persistence of this taxon until fairly recently in the region.7
Further southwards, additional specimens (O. sloggetti sensu lato) were recovered from Klipdrift Shelter, a coastal archaeological site situated in the De Hoop Nature Reserve. Two specimens were found in horizons dated to 51.7±3.3 ka, and another in deposits dated to 60.3±3.8 ka.24 At least three individuals were found in layers that are undated, but assumed, on the basis of stratigraphy, to be younger than 52 ka. Otomys sloggetti sensu lato (represented by M1 molars) was also recovered from the Klasies River main site in the Howiesons Poort layers (~65-46 ka) (T.H.N. unpublished data), and has also been identified in a couple of west coast fossil sites dating to the Holocene (Dunefield Midden) and Late Pleistocene (at the palaeontological site of Hoedjiespunt 125).
The unexpected find of what appeared to be O. cf. sloggetti in fossil deposits was then repeated during investigations of micromammals from modern owl pellet accumulations (Figures 3 and 4). Two five-laminate M3 molars were recovered from a comparative eagle owl pellet collection made from Keurbooms, near Wilderness on the south coast in 20 0 57, and additional O. cf. sloggetti teeth were found by T.H.N., in October 2018, in barn owl pellets from two separate locations in the De Hoop Nature Reserve. On the west coast, O. cf. sloggetti was also found (represented by M1 and M3 molars) in what appeared to be a relatively modern, disaggregated barn owl roost site from Diepkloof Rock Shelter near Elands Bay, which had been deposited on top of an archaeological horizon (Parkington JP 2018 June 1, personal communication).
As mentioned above, O. cf. sloggetti was identified on the basis of five laminae on the M3, and four laminae on the M1 (with the anterior laminae of the latter being roughly ovoid in shape, and clearly differing in morphology from associated O. karoensis and O. irroratus specimens which have rectangular-shaped anterior laminae). This diagnosis became more complicated, however, as morphological differences in the cranial bones and the M3 were then noted between two incomplete skulls obtained from modern owl pellet collections made in De Hoop Nature Reserve, namely the Elandspad Bulk Pellet sample (EPF), and De Hoop Collections Cool Room roost site (DHC). This find had potential implications for the identification of O. sloggetti in the fossil record on the basis of using a five-laminate M3, and it became clear that further investigation was needed into the identity of the modern five-laminate taxa. The two skulls were thus CT scanned and compared with those of O. irroratus, O. karoensis, and O. sloggetti (Figures 3 and 4). The former two species were included as they are ubiquitous throughout South African fossil sites.6
Figure 3 illustrates that O. sloggetti (TM-16519B) differs quite considerably from the two skulls from the pellets, as well as O. karoensis and O. irroratus, in general skull form and proportions: the skull cap is rounder in shape relative to the others, the orbital foramen is broader and extends less anteriorly to posteriorly, the nasal foramen is rounded in shape rather than rectangular, and the shape and size of the maxilla, zygomatic arch and the zygomatic plate are likewise clearly different. The upper incisors of all the taxa exhibit a single groove. The DhC-10-3 skull appears very similar, although not identical, to O. karoensis in terms of the shape of the zygomatic arch, general skull shape, the infraorbital foramen, and the premaxilla. Small differences may be attributed to the fact that it belongs to a smaller and (as indicated by the wear pattern on the molars) younger, individual. The premaxilla and maxilla from the Elandspad Bulk Sample (EPF-bulk-1) specimen differed in size and morphology, not only to O. sloggetti, but also to all the other taxa illustrated in Figure 3. In order to illustrate these morphological differences, and to compare this specimen with another of a similar M3 molar morphology, Figure 4 focuses on the maxilla and the zygomatic arch of EPF-bulk-1 and compares it with the modern comparative five-laminate M3 cf. O. karoensis (DHC-10-3).
The area where the incisive bone meets the zygomatic process of the maxilla is curved in O. karoensis (DHC-10-3) but straight in the indeterminate Otomys from EPF. Other differences include the size and shape of the infraorbital foramen (much larger in the case of the EPF Otomys), and the zygomatic arch is different in size and shape between the taxa. The differences observed in maxillary morphology are likewise reflected in the M3, as illustrated in Figure 5, where the distribution of the laminae across the occlusal surface of the tooth, and the size and shape of the anterior lamina, differ. The fact that the Elandspad specimen shows morphological differences in the maxilla and the M3 supports the identification of this as a taxon or morphotype differentiate from O. karoensis. A search was made through the Elandspad owl pellets for M1 teeth which looked different from those of O. karoensis and O. irroratus, and which could possibly be a match for the M3 Elandspad specimen. Two such specimens were recovered from the Elandspad bulk collection, one of which is illustrated in Figure 7. It is distinguished by a notably large, round, first lamina (EPF-bulk-12, see inset photo in Figure 7).
Comparison between extant and fossil Otomys taxa
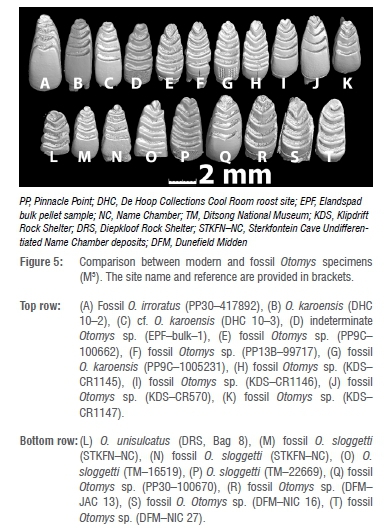
Figure 5 shows a comparison of the M3 of modern O. sloggetti from two different regions in South Africa with each other, and with fossil specimens from archaeological sites on the south coast, west coast, and the Cradle of Humankind (Sterkfontein Cave, Gauteng Province). An Otomys irroratus specimen is included in both Figures 5 and 7 to illustrate the similarity of this species in terms of molar morphology to O. karoensis. The site name and specimen reference numbers are provided in brackets in Figure 5.
The morphological differences could be attributable to biogeography, but are far greater than the degrees of interspecific variation generally observed in murids (T.M. and T.H.N. personal observation), and most likely reflect genetic divergence, like that observed between modern O. sloggetti populations living in mountainous areas in the Drakensberg, Sneeuberg and Lesotho.2 This suggestion is supported by research8 which notes that specimens identified as O. sloggetti in the field were found to represent two genetic clades. The one clade was consistent with the description of O. sloggetti, but the other was distinct, not only from O. sloggetti, but from all other Otomys species.2,8 This species, endemic to the southern Drakensberg Mountain Range, has subsequently been described (nov. sp. Otomys willani).2It is differentiated from O. sloggetti by six laminae on the M3 and grooved lower incisors. Intraspecific variation, albeit to a less marked degree, is also observed in the Sterkfontein Cave O. sloggetti specimens (Figure 5, M and N) in terms of the degree of curvature of the laminae, and general tooth shape. Both specimens come from the so-called 'Name Chamber' deposits which are mixed in that they contain deposits from both Member 4 and Member 5E, with the Oldowan (Member 5e) deposits dating to ~2-1 Ma, and/or Member 4 (~2.8-2 Ma).26 The taxonomic relevance of the morphological differences is unclear, given the stratigraphic/age uncertainties. The Sterkfontein fossil specimens are considerably smaller than the modern comparative O. sloggetti and almost all the fossil O. sloggetti sensu lato, providing an example of the flexibility in size of this genus in both modern and fossil populations (see Appendix 1).
As mentioned previously, O. karoensis has been noted to sometimes have five laminae on the M3. The DHC specimens in Figure 5 ('B' and 'D') illustrate such a case. These specimens differ slightly in that DHC 10-2 ('B') has a tiny auxiliary cusplet situated proximally on the anterior laminae, which indicates a variation occurring in the phenotype. Such additional cusplets (note this feature also appears in the fossils 'G' and J' in Figure 5) would disappear with wear and would not be discernible in older individuals. This kind of intraspecific variation is not uncommon among murids, and small additional cusplets occur infrequently on murid teeth (T.H.N. and T.M. personal observation).
The EPF M3 (EPF-bulk-1, see 'D' in Figure 5), whose maxilla and premaxilla show differences to O. karoensis as noted above, is different from the fossil and modern O. karoensis in that it has a larger and more rounded anterior cusp, which is embedded in the second lamina to a greater degree than seen in O. karoensis. It also differs in that the spacing between the anterior to the posterior lamina of the occlusal surface is wider, and the laminae are more curved and orientated centrally towards the middle of the tooth, whereas O. karoensis laminae slant towards the labial side of the tooth.
The south coast fossil specimen from PP30 (Figure 5 'Q') shows a close fit in terms of morphology and size with the modern O. sloggetti specimen TM-22669 (Figure 5 'P'), as does one of the three fossils from Dunefield Midden (Figure 5 'R'), but the two other Dunefield Midden fossil specimens (Figure 5 'S' and 'T'), also originally identified as O. sloggetti sensu lato, are harder to match as the occlusal enamel-dentine surface of the teeth is very worn.
Four fossil specimens from Pinnacle Point (PP9C and PP13B) (Figures 5 and 6, E, F and G), and four from Klipdrift shelter (Figures 5 and 6, H, I, J and K), were previously thought to represent O. cf. sloggetti; however, they appear morphologically very similar to the modern specimen from EPF (EPF-bulk-1), in terms of size, morphology, orientation of laminae, and general tooth shape. The fossil Otomys 'G' (PP13B), as well as 'J' and 'K' (KDS), all have five laminae (G and J also exhibit a tiny auxillary cusplet), and are morphologically very similar to the five-laminate O. karoensis in terms of shape and orientation of laminae, and we tentatively suggest that they belong to this species.
A comparison of the specimens initially identified as O. sloggetti during analysis of the various archaeological sites may be split into two morphological groups, arbitrarily called Morphotype 1 and Morphotype 2. Morphotype 1 (Figure 5: D, E, F, H and I) includes the modern EPF specimen, and fossils from KDS, PP9C and PP13B. Morphotype 1 is defined as exhibiting a large, roundish anterior laminae which is embedded into the second laminae, and the other laminae are curved. This differs to O. karoensis and O. irroratus where the laminae are more linearly arranged, and the anterior lamina is relatively smaller and more rounded-rectangular in shape. Morphotype 2 was identified in the fossil sites of PP30 and DFM and includes P Q and R, and probably S and T, as well as one of the modern comparative O. sloggetti (TM-22669), which the fossils most closely resemble in terms of general morphology, size, and orientation of laminae. Morphotype 2 teeth yielded greater breadth measurements (breadth of M3: 2.0-2.2 mm, length of M3: 3.2-4 mm) than Morphotype 1 teeth (breadth of M3: 1.8-1.9 mm, length of M3: 2.9-3.6 mm), but overlapped in terms of length.
When viewed laterally, the five, or six, laminae present on the various Otomys taxa, as well as the auxillary cusplets, are clearly discernible (Figure 6). Note that the specimens, and the order of specimen line-up, are identical in Figures 5 and 6.
Figure 7 makes a similar comparison to that of Figure 5, but this time with the M1 from modern O. sloggetti, fossil specimens from the south and west coasts and Sterkfontein Cave. Unfortunately, no M1 belonging to O. cf. sloggetti or O. karoensis were recovered from Dunefield Midden and this site is thus not represented in Figure 7.
In Figure 7, the top row of specimens from B to E are considered to represent O. karoensis, although E from PP9C was originally differentiated from other O. karoensis at the site on the basis of size and the relative roundness of the first lamina. However, it appears very similar to the modern specimen from DHC 10-2 (Figure 7, D), and given this likeness, is most probably an O. karoensis. Fossil O. unisulcatus specimens are represented in Figure 7 (see S, T, U and V), while W represents a modern specimen (from Diepkloof Rock Shelter).
The molar F (from Sterkfontein Cave) shows similarities with O. karoensis, although it was the closest match to O. sloggetti out of a large number of M1 molars examined from the Sterkfontein Name Chamber deposits. This lack of an obvious match to the M3O. cf. sloggetti at Sterkfontein could be due to a number of circumstances; however, it is worth considering the possibility that the M3 molars, which admittedly look convincingly like O. sloggetti, belong to a taxon which has an M1 which differs from that of O. sloggetti. The fossils G, J, K, L, M, N, O, P Q and R were all originally identified as O. cf. sloggetti, but when they are lined up and compared it becomes clear that there is a good deal of variation, and, as noted in the discussion above, we currently have no understanding of the link between the different morphotypes and if they represent the same taxon. The fossils J, K, L, M, N and O (also possibly 'P' although digestion and wear complicate assessment) all appear to represent the same morphotype and come from KRM, PP13B, PP30, and KEH1. This morphotype (called Morphotype 3) is represented by an anterior lamina which is rectangular in shape and embedded at an angle into the second laminae (size range: 2.8-3.3 mm length, 1.7-1.9 mm breadth). The modern Otomys sp. from Diepkloof Rock Shelter (Q) and the specimen from KDS (R) have more rounded first laminae, and the orientation of the laminae differs from the other fossils, and from each other. Both these specimens fall out of the size ranges shown by Morphotype 3, and are smaller in both length and breadth. The modern Diepkloof specimen (Q; 2.7 mm length, 1.5 mm breadth) shows a strong morphological resemblance to the fossil Otomys species from PP30 (G; 2.8 mm length, 1.7 mm breadth) although it is smaller. Differentiating morphotypes in the M1 is thus less straightforward than seen in the M3, and although specimens J, K, L, M, N, O and P appear to represent one morphotype (referred to as Morphotype 3 from this point), there are a number of other specimens, such as G and Q (Morphotype 4), and R (which is relatively narrow, and unique in having a small round anterior lamina and relatively linearly orientated laminae) which represents another morphotype (Morphotype 5). The linear arrangement of cusps in Morphotype 5 resemble those of the modern specimen O. sloggetti (TM-16519), whereas Morphotype 3, Morphotype 4 and Morphotype 5 specimens all share some similarities with the other modern comparative O. sloggetti (TM-22669), but are generally smaller. Once again, it is impossible to interpret the observed variation, which could be indicating a different sub-species or species, or may simply be reflecting morphological plasticity in the phenotype. Notably, morphological differences between the fossil specimens and the one modern O. sloggetti specimen (TM-22669) are less than that observed between the two modern museum specimens in Figure 7. As observed in the M3, the two museum specimens differed in the size and distribution of the cusps varying across the occlusal surface, and the extent of curvature of the laminae (see Figure 7, H and I). The two modern O. sloggetti (TM-22669, TM-16519) were broader than all the fossil specimens measured (O. unisulcatus excluded), although there was some overlap with the fossils in terms of length.
The modern specimen from EPF (EPF-bulk-12, Figure 7, 'X') (hereafter referred to as Morphotype 6) is differentiated from Morphotypes 3, 4 and 5 taxa by a larger (relative to the other laminae) and more rounded anterior cusp - the former have a smaller and more rectangular-rounded shape. This M1 (two molars with this morphology were recovered from the bulk samples) may match the unidentified Otomys maxilla and M3 from the EPF bulk pellet samples, or it may represent a variation of cf. O. karoensis. The similarity observed in the morphology of the M3 of the EPF specimen with some of the fossils (i.e. Morphotype 1), is not as obvious when it comes to the M1, and it is not possible to match the EPF M1 and M3 specimens with any certainty.
Discussion and conclusions
An intrasite examination of fossil specimens identified as O. cf. sloggetti during previous research indicates that, although there is homogeneity observed between some specimens, there is a degree of variation, which, together with the differences observed between modern comparative O. sloggetti material, is very hard to interpret. There are, however, two morphotypes distinguishable in the fossil O. cf. sloggetti M3, and four in the M1, material, as described above. We suggest that the differentiation of these morphotypes, at least in the case of Morphotypes 1 and 2 in the case of the M3, and Morphotype 3 in the case of the M1 is validated by the fact that several specimens of the same morphotypes have been found in several south coast fossil sites, a couple of west coast sites, and in both fossil and modern material. In addition, the morphotypes were recognised as differing morphologically to O. karoensis and O. irroratus, and most closely resembling O. sloggetti, by two independent researchers. This, together with the fact that the fossil record indicates the presence of the morphotypes from ~150 000 ka, suggests that they have some kind of phenotypic origin and are not merely local aberrations. It is not, however, possible to match the upper and lower morphotypes with any certainty, which is indicative of the fact that we have no clear understanding of the relevance of the different morphotypes, and their genetic affiliations. It is also not possible to ascertain the relationship between the different morphotypes with O. cf. sloggetti, or other Otomys taxa. There is a good morphological match between the unidentified Otomys from the EPF bulk sample, and the fossil Morphotype 1, but at this stage it is impossible to say if they represent the same, or some closely related, taxon. The precise relationship of the EPF bulk sample Otomys taxon to the fossil (and modern) taxa remains uncertain given all the unknown variables, and the evidence for appreciable genetic divergence in O. sloggetti, and other Otomyini. In conclusion, both the fossil and modern specimens suggest a large degree of variation in molar morphology and size, and both Morphotype 1 and Morphotype 2 in the case of the M3, and Morphotypes 3, 4 and 5 in regard to the M1, show affinities with O. sloggetti. The genetic relevance of the observed similarities and differences will remain obscure until they can be interpreted within a framework which clarifies the link between skull and molar morphology, and the genotype of modern taxa.
The presence of O. cf. sloggetti in surface deposits in some of the Pinnacle Point sites was puzzling, and it was suggested that this might have occurred due to mixing with fossil deposits.7 This review of the purported O. sloggetti material from the owl pellets and fossil sites suggests that it is probably not O. sloggetti, and, as noted above, the jury remains out as to the exact phylogeny of the different morphotypes. The observed differences between the two O. sloggetti museum specimens illustrate the fact that museum collections may not provide a reliable taxonomic reference where cryptic species are involved, and are likely to be related to the fact that the specimens were collected from different mountain ranges, and probably represent different species.2
It has been noted that specimens identified as O. sloggetti in the field were found (in an investigation utilising nuclear and mitochondrial gene regions, as well as morphological characteristics8) to represent two clades. One was consistent with the description of O. sloggetti, but the other was distinct not only from O. sloggetti, but from all other Otomys species, and the authors concluded that this could be a new, and novel, species. Subsequent to this, a new Otomys species was described from the Sneeuberg Centre of Floristic Endemism in the southern Drakensberg Mountains (Otomys willani sp. nov.), and the same publication notes the presence of two specimens from the Mountain Zebra National Park in the Eastern Cape which showed a well-supported and relatively deeply divergent lineage that has no supported relationship with any other Otomyini species.2 The fossil and modern material discussed in this paper suggests that such cryptic, undescribed species, or subspecies, have been, and may be, co-occurring with modern populations of O. karoensis and O. irroratus. Phylogenetic studies need to be done in conjunction with morphological studies of skulls and molars, as, currently, the relationship between the huge variation seen in interspecific morphology with genetics is little understood, different Otomys species are not always distinguishable morphologically, and considerable chromosomal polytypes have been found.2 The otomyines are an interesting family of murids and assessing the degree of morphological variation in modern O. sloggetti, O. irroratus/O. auratus, and O. karoensis populations will provide essential information on morphological variation, and how it relates to biogeography and adaptation, and will in turn provide interesting ecological and evolutionary information on a family of small mammals which exhibit great adaptability and phenotypic plasticity. Clearly, further genetic and cladistic research is needed to unravel the cryptic and complicated taxonomy of Otomys taxa. Given the issues raised by this research into the south and west coast fossil record, we recommend that future research into the Otomyini focus on modern collections (including owl pellet collections) from the south and west coasts and the Eastern Cape, using an integrated molecular, karyotypic and morphological approach.
Acknowledgements
The support of the DST-NRF Centre of Excellence in Palaeosciences (to T.M.) and SFF Centre for Early Sapiens Behaviour (to T.H.N.) towards this research is hereby acknowledged. Financial support was also received from the National Research Foundation of South Africa (T.M.) and the National Research Council of Norway (T.H.N.), and is duly acknowledged. Thanks are due to Curtis Marean (SACP4: South Africa Coast Paleoclimate, Paleoenvironment, Paleoecology, Paleoanthropology Project) and Naomi Cleghorn who led the Pinnacle Point and Knysna Head excavations, respectively; Christopher Henshilwood who led the Klipdrift Shelter excavations; and Sarah Wurz who led the Klasies River excavations. We also thank Prof. John Parkington and Louisa Hutton (University of Cape Town) who provided access to the Dunefield Midden and Diepkloof Rock Shelter micromammal material, and to Margaret Avery, Cape Nature staff and Ole Fredrik Unhammer for help in locating and collecting modern owl pellets from the De Hoop Nature Reserve. Kathleen Kuman and Bernhard Zipfel kindly provided access to the Sterkfontein fossil material. A big thank you to Muofhe Tshibalanganda, Stefan le Roux and Anton du Plessis at the Central Analytical Facilities CT scan facility (Stellenbosch University) for all their assistance and support during the scanning and rendering of fossil and modern material.
Competing interests
We declare that there are no competing interests.
Authors' contributions
T.M. was the project leader and carried out all the CT scanning on extant and fossil material, and took the images for the publication with the exception of Figure 2 which was done by T.H.N. T.M. and T.H.N. both contributed to the various drafts leading up to the submission of this article. Both T.M. and T.H.N. provided fossil material, and assessed the morphotypes reported on in this paper.
References
1. Monadjem A, Taylor PJ, Denys C, Cotterill FPD. Rodents of sub-Saharan Africa: A biogeographic and taxonomic synthesis. Berlin/Munich/Boston: Walter de Gruyter; 2015. [ Links ]
2. Taylor PJ, Kearney T, Dalton DL, Chakona G, Kelly CM, Barker NFP Biomes, geology and past climate drive speciation of laminate-toothed rats on South African mountains (Murinae: Otomys). Zool J Linn Soc. 2019;189(3):1046-1066. https://doi.org/10.1093/zoolinnean/zlz134 [ Links ]
3. Taylor PJ, Maree S, Van Sandwyk J, Baxter R, Rambau RV. When is a species not a species? Uncoupled phenotypic, karyotypic and genotypic divergence in two species of South African laminate-toothed rats (Murinae: Otomyini). J Zool. 2009;277(4):317-332. https://doi.org/10.1111/j.1469-7998.2008.00542.x [ Links ]
4. Taylor PJ, Lavrenchenko LA, Carleton MD, Verheyen E, Bennett NC, Oosthuizen CJ, et al. Specific limits and emerging diversity patterns in east African populations of laminate-toothed rats, genus Otomys (Muridae: Murinae: Otomyini): Revision of the Otomys typus complex. Zootaxa. 2011;3024:1-66. http://hdl.handle.net/2263/18125 [ Links ]
5. Engelbrecht A, Taylor PJ, Daniels SR, Rambau RV. Cryptic speciation in the southern African vlei rat Otomys irroratus complex: Evidence derived from mitochondrial cyt b and niche modelling. Biol J Linn Soc. 2011;104(1):192-206. https://doi.org/10.1111/j.1095-8312.2011.01696.x [ Links ]
6. Edwards S, Claude J, Van Vuuren BJ, Matthee CA. Evolutionary history of the Karoo bush rat, Myotomys unisulcatus (Rodentia: Muridae): Disconcordance between morphology and genetics. Biol J Linn Soc. 2011;102(3):510-526. https://doi.org/10.1111/j.1095-8312.2010.01583.x [ Links ]
7. Matthews T, Marean CW, Cleghorn N. Past and present distributions and community evolution of Muridae and Soricidae from MIS 9 to MIS 1 on the edge of the Palaeo-Agulhas Plain (south coast, South Africa). Quat Sci Rev. 2019;235, Art. #105774. https://doi.org/10.1016/j.quascirev.2019.05.026 [ Links ]
8. Phukuntsi MA, Kearney T, Brettschneider H, Dalton DL, Oosthuizen M. Hidden Identities: Cryptic species in the Otomys genus (Cuvier 1824) (Rodentia: Muridae: Otomyinae) revealed by mitochondrial and nuclear DNA in South Africa. J Phylogenet Evol Biol. 2016;4, Art. #168. http://doi.org/10.4172/2329-9002.1000168 [ Links ]
9. Taylor PJ, Denys C, Mukerjee M. Phylogeny of the African murid tribe Otomyini (Rodentia), based on morphological and allozyme evidence. Zool Script. 2004;33:389-402. https://doi.org/10.1111/j.0300-3256.2004.00157.x [ Links ]
10. Denys C. Phylogenetic affinities of the oldest East African Otomys (Rodentia, Mammalia) from Olduvai Bed I (Pleistocene, Tanzania). Neues Jahrb Geol Palaontol Abh. 1989;12:705-725. https://doi.org/10.1127/njgpm/1989/1990/705 [ Links ]
11. Chevret P, Denys C, Jaeger JJ, Michaux J, Catzeflis F, Molecular and palaeontological aspects of the tempo and mode of evolution in Otomys (Otomyinae: Muridae: Mammalia). Biochem Syst Ecol. 1993;21:123-131. https://doi.org/10.1016/0305-1978(93)90017-l [ Links ]
12. Wilson DE, Reeder DM, editors. Superfamily Muroidea. Mammal species of the world: A taxonomic and geographic reference. Baltimore, MD: JHU Press; 2005. p. 894-1531. [ Links ]
13. Avery DM. A preliminary assessment of the micromammalian remains from Gladysvale Cave, South Africa. Palaeontol Afr. 1995;32:1-10. [ Links ]
14. Avery DM. Late Quartenary incidence of some micromammalian species in Natal. Durban Museum Novitates. 1991;16(1):1-11. [ Links ]
15. Avery DM. Notes on the systematics of micromammals from Sterkfontein, Gauteng, South Africa. Palaeontol Afr. 2000;36:83-93. http://hdl.handle.net/10539/16381 [ Links ]
16. Denys C. Of mice and men. Evolution in East and South Africa during Plio-Pleistocene times. African biogeography, climate change and human evolution. New York: Oxford University Press; 1999. p. 226-252. [ Links ]
17. Matthews T, Marean C, Nilssen P Micromammals from the Middle Stone Age (92-167 ka) at Cave PP13B, Pinnacle Point, south coast, South Africa. Palaeontol Afr. 2009;44:112-120. [ Links ]
18. Nel TH, Henshilwood CS. The small mammal sequence from the c. 76-72 ka Still Bay levels at Blombos Cave, South Africa - Taphonomic and palaeoecological implications for human behaviour. PLoS ONE. 2016;11(8), e0159817. https://doi.org/10.1371/journal.pone.0159817 [ Links ]
19. Nel TH, Wurz S, Henshilwood CS. Small mammals from Marine Isotope Stage 5 at Klasies River, South Africa - Reconstructing the local palaeoenvironment. Quat Int. 2018;471:6-20. https://doi.org/10.1016/j.quaint.2017.08.074 [ Links ]
20. Pillay N, Taylor P Baxter R, Child MF. Otomys sloggetti. The IUCN Red List of Threatened Species [webpage on the Internet]. c2017 [cited 2019 Sep 09]. e.T15659A110019838. http://dx.doi.org/10.2305/IUCN.UK.2017-2.RLTS.T15659A110019838.en [ Links ]
21. Schwaibold U, Pillay N. Behavioral strategies of the African ice rat Otomys sloggetti robertsi in the cold. Physiol Behav. 2006;88(4-5):567-574. https://doi.org/10.1016/j.physbeh.2006.05.013 [ Links ]
22. Mokotjomela T, Schwaibold U, Pillay N. Population surveys of the ice rat Otomys sloggetti robertsi in the Lesotho Drakensberg. Afr Zool. 2010;45(2):225-232. https://doi.org/10.1016/j.physbeh.2006.05.013 [ Links ]
23. Taylor PJ, Denys C, Mukerjee M. Phylogeny of the African murid tribe Otomyini (Rodentia), based on morphological and allozyme evidence. Zool Script. 2004;33:389-402. https://doi.org/10.1111/j.0300-3256.2004.00157.x [ Links ]
24. Henshilwood CS, Van Niekerk KL, Wurz S, Delagnes A, Armitage SJ, Rifkin RF, et al. Klipdrift shelter, southern Cape, South Africa: Preliminary report on the Howiesons Poort layers. J Archaeol Sci. 2014;45:284-303. https://doi.org/10.1016/j.jas.2014.01.033 [ Links ]
25. Matthews T, Denys C, Parkington JE. The palaeoecology of the micromammals from the late middle Pleistocene site of Hoedjiespunt 1 (Cape Province, South Africa). J Hum Evol. 2005;49:432-451. https://doi.org/10.1016/j.jhevol.2005.05.006 [ Links ]
26. Avery DM, Stratford DJ, Sénégas F. Micromammals and the formation of the Name Chamber at Sterkfontein, South Africa. Geobios. 2010;43(4):379-387. https://doi.org/10.1016/j.geobios.2010.01.002 [ Links ]
 Correspondence:
Correspondence:
Thalassa Matthews
Email: tmatthews.matthews@gmail.com
Received: 10 Sep. 2019
Revised: 01 June 2020
Accepted: 17 July 2020
Published: 29 Jan. 2021
Editor: Margaret Avery
Funding: South African National Research Foundation, National Research Council of Norway



{kind=link}