










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.117 no.1-2 Pretoria ene./feb. 2021
http://dx.doi.org/10.17159/sajs.2021/6758
RESEARCH ARTICLE
Hominin lower limb bones from Sterkfontein Caves, South Africa (1998-2003 excavations)
Travis Rayne PickeringI; Jason L. HeatonIII; Ron J. ClarkeII; Dominic StratfordIV; A.J. HeileI
IDepartment of Anthropology University of Wisconsin-Madison' Madison, Wisconsin, USA
IIEvolutionary Studies Institute' University of the Witwatersrand, Johannesburg, South Africa
IIIDepartment of Biology, BirminghamSouthern College, Birmingham, Alabama, USA
IVSchool of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
We describe late Pliocene and early Pleistocene hominin fossils from Sterkfontein Caves (South Africa), including two femoral specimens, as well as a partial tibia and a partial fibula. The fossils are likely assignable to Australopithecus africanus and/or Australopithecus prometheus and the morphology of each corroborates previous interpretations of Sterkfontein hominins as at least facultative bipeds.
SIGNIFICANCE:
• A recent series of papers by our research team describes the morphology of a hominin skeleton from Sterkfontein Caves (South Africa), nicknamed 'Little Foot'. Based on its unique skull morphology, R.J. Clarke, the skeleton's discoverer, places it in the species Australopithecus prometheus, as distinct from the better-known and co-occurring Australopithecus africanus. Here we describe additional hominin thigh and leg fossils from Sterkfontein that, when considered in a comparative context, support the hypothesis that there was significant (probably interspecific) variation in South African hominin postcranial morphology during the late Pliocene and early Pleistocene.
Keywords: Australopithecus, femur, tibia, fibula
Introduction
A long history of palaeoanthropological research at Sterkfontein Caves (Gauteng, South Africa), starting in 1936, has yielded a massive store of hominin fossils.1-5 As one in a series of papers, we report here on four previously undescribed hominin fossils excavated by R.J.C. between 1998 and 2003. Those other papers present descriptions and interpretations of sizeable samples of teeth6 and of axial and upper limb bones7,8. The sample described here comprises fewer materials, representing two femur specimens, a partial tibia and a partial fibula. All four specimens preserve sufficient anatomy to indicate that the individuals from which they derived were bipeds, but more comprehensive functional interpretations of the fossils are limited by their fragmentary nature. Each also preserves taphonomic information that we report.
Two of the fossils derive from Member 4 of the Sterkfontein Formation, the site's most hominin-rich deposit, which is probably slightly greater than 2.6 to around 2.5 million years old (Ma) (Figure 1).9-13 The other two are part of the Jacovec Cavern fossil assemblage, which includes a modest sample of hominin specimens, previously described by Clarke14. The original interpretation of the Jacovec stratigraphy identified several generations of cave fill, including three sedimentary units identified as 'orange', 'brown', and 'stony'.14 More recent research demonstrates multigenerational infilling and reworking of the chamber's sediments, which created several secondary deposits and a talus composed of orange and brown units.15 Dating of the Jacovec deposits is still in progress. Based on the presence of a fossil attributed to Equus, Kibii16 proposed a relatively young age of 2.34 Ma for the recovered faunal assemblage. However, Partridge et al.14 provided an age of 4.02±0.27 Ma for orange sediments exposed on the chamber ceiling, which yielded a partial hominin cranium cataloged as StW 578. It is concluded that all hominin fossils thus far recovered from the chamber floor derive from this in-situ orange unit.14,15 Accordingly, we propose that until such time as multiple, stratigraphically associated, and dateable proxies contradict this radiometric date, it remains the best plausible estimate for the age of the Jacovec fossils. Based on craniodental analyses, Clarke5,17-19 has argued for the presence of two coeval hominin species in Member 4: Australopithecus africanus and Australopithecus prometheus. Additionally, analysis of hominin teeth from Jacovec indicate that both taxa also occur in that deposit.
Methods
We provide information on each fossil's spatial context, its relative completeness and condition, and its morphology and osteometrics. The legend to Figure 1 summarises the excavation coordinate system employed at Sterkfontein. As to condition, we recorded the degree to which a specimen suffered subaerial weathering, using Behrensmeyer's20 well-known weathering stage system, and noted its degree of staining by manganese dioxide. Bone surface modifications were also identified using 10x power magnification.21 All fractured bone surfaces were assessed with reference to the 'angle formed by the fracture surface and bone cortical surface'22(p.34). Typically, fracture angles on long limb bones that were created when the bone was 'green' (i.e. before significant loss of a bone's organic fraction and its desiccation) are usually either acute or obtuse, while those created on dry long limb bones are usually right angles.22-24 We used Mitutoyo™ digital calipers to collect standard osteometric linear measurements and collected standard osteometric angular measurements with SPI™ 0-180° protractors.25-27 We followed recently published methods28 in order to derive femoral neck anteversion on the single proximal femur specimen in the analysed sample.
Results
StW 598 (Jacovec Cavern)
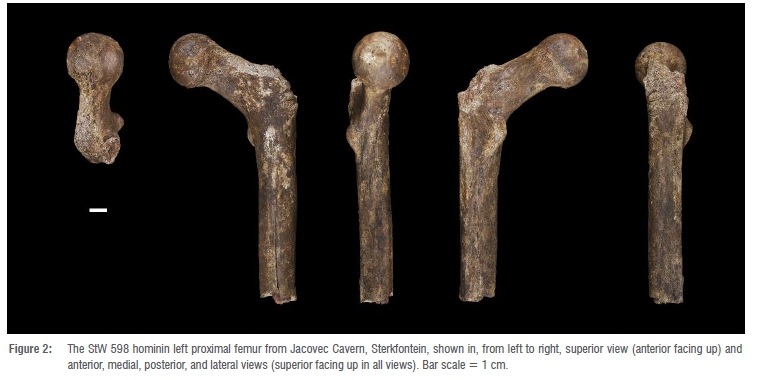
This proximal left femur was described preliminarily by Clarke14. It is an exquisite specimen, stained uniformly by manganese dioxide but essentially unweathered (stage 0) and preserving the head and nearly half of the diaphysis (maximum length = ~153.0 mm) (Figure 2). The fossil is truncated distally along a relatively straight, right-angled breakage plane, indicating that the bone was leached of much, if not all, of its organic content when it was fractured.
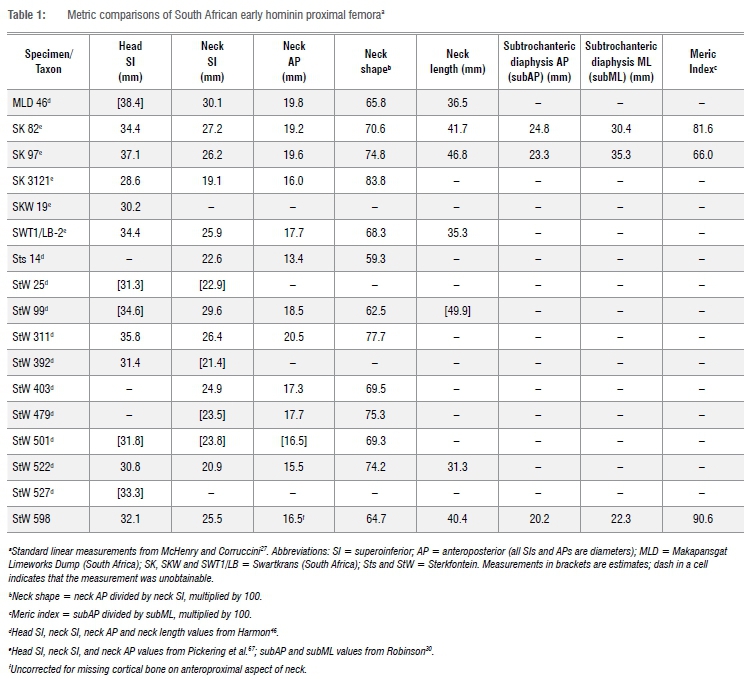
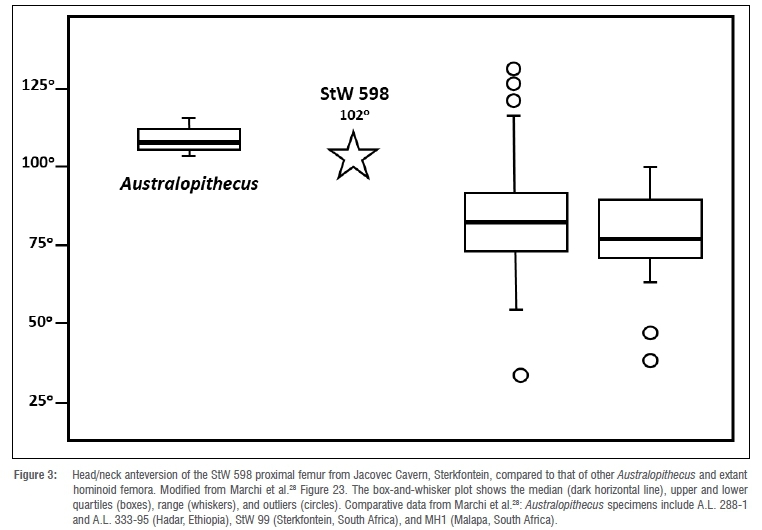
Further, the fracture surface is partially coated by breccia, attesting to the antiquity of the breakage event that created it. A crack emanates from the middle of the anterior edge of this fracture surface and travels proximally, along the middle of the anterior surface of the diaphysis, for a length of ~80.0 mm. The crack is wider mediolaterally for its distal ~two-thirds than it is proximally. A similarly constructed longitudinal crack courses the middle of the posterior diaphysis (along the medial lip of the linea aspera), starting distally in the middle of the dorsal edge of the specimen's distal fracture surface and terminating ~78.0 mm proximal to that point, near the junction of the base of the lesser trochanter and the proximal terminus of the pectineal line. Both cracks are infilled by breccia, which caused their mediolateral expansions, especially distally. Other ancient modification to StW 598 occurs at its proximolateral corner. Unlike the previously described incidences of damage, this alteration of the fossil was most likely inflicted when the bone was still fresh and its trabeculae were packed with nutritionally attractive red marrow and grease. The crenulated surface3 of the base of the largely missing greater trochanter, as well as several deeply invasive tooth gouges in the exposed trabeculae at the anterolateral portion of the distal margin of that damaged surface, indicate that the trochanter was chewed away by a prehistoric carnivore(s). This carnivore-inflicted damage continues superomedially across the anteroproximal femoral neck, where several layers of lamellae were peeled away in a dorsoventrally wide strip (maximum width = ~14.0 mm) that terminates -11.0 mm from the superior rim of the femoral head. The head of StW 598 is small and spherical, with a relatively large fovea capitis that is placed dorsally on the medial aspect of the joint surface (Table 1). The fovea capitis is deep, with a superior and dorsal margin that is contiguously sharply raised. The femoral head is hafted to the bone's diaphysis at a neck-shaft angle of 121°, via a mediolaterally elongated and anteroposteriorly compressed neck (Table 1). StW 598 has a fairly marked femoral anteversion angle of 102° (Figure 3). The obturator externus groove of the specimen is not palpable superiorly and just barely so dorsolaterally. Likewise, the specimen's intertrochanteric crest and intertrochanteric and spiral lines are underdeveloped. In contrast, the fossil shows a small but robust, superoinferiorly elongated (maximum superoinferior length = -16.3 mm; maximum mediolateral width = -10.4 mm) lesser trochanter, a well-developed pectineal line and an especially rugose gluteal tuberosity.
The lesser trochanter projects slightly medially beyond the medial margin of the proximal diaphysis, so that it (the tubercle) is just visible when viewing the anterior aspect of the bone. The gluteal marking is a well-developed, posterolaterally placed ridge that merges with the pectineal line -49.0 mm distal to the inferior margin of the lesser trochanter, at a small, distally opening nutrient foramen, to form the superior terminus of the mediolaterally broad linea aspera. Viewed medially or laterally, the subtrochanteric diaphysis appears relatively flat anteriorly (especially proximally) and only slightly more convex posteriorly, forming a gentle posterolaterally placed angle along the course of the linea aspera (Table 1). The shaft cross-section is roughly circular at the distal fracture edge, with an anteroposterior diameter of 20.5 mm and mediolateral diameter of 21.1 mm at that level (these dimensions are uncorrected for the slight mediolateral gaps in the antero- and posterodistal diaphysis described above). The thickest portion of the cortex at this natural fracture surface is posteriorly, at the position of the linea aspera (7.4 mm). Because the diaphyseal midpoint cannot be determined precisely on the broken specimen, we cannot report a pilasteric index (midshaft anteroposterior diameter divided by midshaft mediolateral diameter, multiplied by 1 0029). However, an index of 97 is calculated using these measurements taken from the distalmost edge of the fossil.
StW 619 (Jacovec Cavern)
This partial distal epiphysis of a left femur is heavily mottled by deposits of manganese dioxide but shows little subaerial weathering (stage 0) (Figure 4). All areas of damage on the fossil are ancient, as evidenced by the fact that the exposed trabeculae are smoothed and polished and are filled by red cave sediments. The specimen is broken proximally, along a relatively straight transverse plane, at the inferior margin of the distal metaphysis. The edge of this fracture surface is roughly right-angled for its entire circumference; two short but deep, probable carnivore, tooth scores emanate in an inferoanterior direction from the medial margin of this fracture edge. Most of the lateral condyle and lateral epicondyle of StW 619 are missing, although much of a roughened impression for the lateral head of the gastrocnemius is preserved. The lateral patellar lip of the specimen is broken away laterally and proximally but it is obvious from an inferior view that the lateral lip projected significantly anteriorly (Figure 4). The region of the lateral meniscal groove is missing.
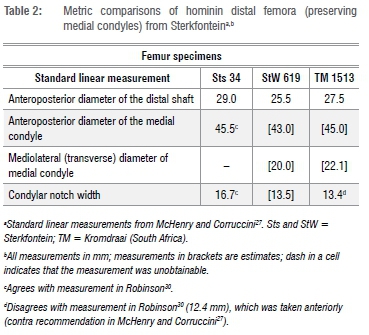
Several areas of the medial condyle are also eroded, including the entirety of its superior border from that feature's anterior point of connection with the medial patellar lip to its dorsal termination. This arc of damage extends laterally into the region in which the medial meniscal groove was positioned, obliterating the medial extent of that feature. We are, however, able to discern what we judge to be the lateralmost extremity of the groove as indicated by arrows in Figure 4. This probable groove remnant is quite shallow. StW 619 lacks a well-developed medial condylar boss. The distal medial condyle does not drop appreciably inferiorly dorsal to the presumed meniscal groove; instead, the distal medial condyle is fairly smooth across its extent, except for a low and mediolaterally narrow, anteroposteriorly elongated elevation along the length of its lateral edge. The adducter tubercle of StW 619 projects only modestly in superior direction, away from the solid mass it forms in connection medially with the blunt, strongly projecting medial epicondyle. In medial view, the medial condyle is elliptical in shape. Maximum (non-anatomical; taken mediolaterally) linear length of StW 619 is 55.5 mm. In general form and size, StW 619 is very similar to Sts 34 and TM 1513 - two hominin distal femora from Sterkfontein Member 4 that also preserve medial condyles (Table 2).30
StW 653 (Member 4, mixed S,T,U/47)
This partial proximal epiphysis and extreme proximal metaphysis of a right tibia is well stained by manganese dioxide and is unweathered (stage 0) (Figure 5). Its maximum length is mediolateral at 49.7 mm. The anterolateral aspect of the specimen is missing and the exposed trabeculae in that area are filled with lightly calcified breccia. Dorsolaterally, the specimen's proximal end preserves damage reminiscent of ragged edge chewing imparted by carnivores (cf.Brain3), but there is no independent indication of feeding damage on the rest of the specimen in the form of tooth scores or pits. The distal fracture surface of StW 653 is obscured by adhering breccia but appears to be right-angled.
Most of the medial condyle of StW 653 is still intact, as is the approximate dorsal third of the lateral condyle. The medial condyle is dorsoventrally elongated (estimated anteroposterior diameter = 34.5 mm; estimated mediolateral diameter = 22.5 mm) and slightly concave, bounded by a blunt but superiorly projecting rim dorsomedially (the rest of the rim is missing so we cannot comment on its original form). The concavity of the medial condyle is accentuated by the acute manner in which the medial intercondylar tubercle rises from it superiorly. The intercondylar eminence and the posterior and most of the anterior intercondylar areas are also preserved, as is the lateral intercondylar tubercle. The last falls less severely distolaterally toward the lateral condyle than does the medial intercondylar eminence fall distomedially toward the medial condyle. The superior apices of both intercondylar eminences are chipped, exposing their underlying trabeculae. The small portion of lateral condyle is convex dorsally, appears moderately concave ventrally, and is positioned more superiorly than is the medial condyle; its dorsomedial corner appears to be marked by a small 'meniscal notch' (cf. Tardieu31). In sum, the morphology of the tibial plateau is reminiscent of those of 'typical' modern human tibiae. Distal to the plateau, the proximolateral branch of the soleal line shows a degree of development that is also comparable to those of modern human tibiae; the groove for the semimembranosus is humanlike in morphology (i.e. deep and circular ventrally; superoinferiorly shorter and anteroposteriorly elongated dorsally) and position, immediately at the base of the dorsomedial corner of the medial condyle. The attachment area for the medial collateral ligament is very rugged, projecting strongly medially from the proximal metaphysis, inferior to it.
StW 680 (Member 4, M/46 18'5'-19'5')
This partial diaphysis of a hominin fibula, measuring 84.3 mm in maximum length, is well stained by manganese dioxide but is unweathered (stage 0) (Figure 6). It lacks surficial tooth scores or pits, but one of its ends terminates in an irregular break reminiscent of carnivore-induced ragged-edge chewing. This end is densely packed with trabeculae, indicating that it is metaphyseal. This region lacks a diagonal curve of an anterior border or crest toward the position of a lateral malleolus, as would be predicted for a distal fibula portion. Accordingly, we conclude that this part of the specimen is from the proximal end of the bone. Superiorly, the cross-section of the fossil is roughly triangular, with two sides of the triangle being slightly convex and divided by a low ridge, while the third side is flat-to-concave, and bounded on each side by sharply defined crests. When viewed in standard anatomical position, the lateral and posterior sides of a typical hominin fibula are convex, while its medial side, lying between well-defined crests, is concave. Following this understanding, it is clear that StW 680 is a left fibula. In addition, the specimen compares favourably to two other hominin left fibulae from Sterkfontein, StW 356 (Member 4) and StW 573 (Member 2).

The distal termination of the specimen is spiral but its fracture edge is right-angled, indicating that the break was probably induced by static loading (e.g. sediment compaction) and when the specimen was at least partially degreased. Its diaphysis is more robust than those of the two other Sterkfontein fibula mentioned above, and its fibular neck is round in cross-section. Martin and Saller26 recommend that diameters of the fibular neck be taken at that feature's smallest circumference. Following that guideline, the anteroposterior diameter of the neck of StW 680 is 11.0 mm and its mediolateral diameter is 10.5 mm. Distal to the neck, the medial and lateral surfaces diverge from each other at an acute angle formed by a low anterior border that is more elevated (i.e. anteriorly projecting) distally than it is proximally; the medial surface is flat proximally to very slightly concave distally for its preserved length, while the lateral surface bulges convexly for most of its length superiorly along a rounded ridge and is flat inferiorly. The posterior border, for its whole length, takes roughly the same rounded form as the lateral surface. In contrast, the interosseous border is sharp but still does not project all that markedly from the main body of the diaphysis.
Discussion
Neotaphonomic research, utilising modern baboon carcasses as proxies for early hominin cadavers, shows that the knee joint is especially susceptible to destruction by feeding carnivores.32 In this context, it is worth mentioning that taphonomic studies have also concluded that carnivores played at least some role in the creation of the hominin fossil assemblages from Sterkfontein Member 4 and Jacovec.4,7,14 We also note the following general characteristics of the South African hominin fossil record: proximal femora are more common than are distal specimens; proximal tibiae are rarer than are distal specimens; and, fibular specimens of any completeness are exceedingly exceptional.2-5 In sum, it thus seems especially fortunate that the chewed (and possibly chewed) thigh and leg specimens described here survived to become part of the Sterkfontein fossil record.
Member 4 fossils
Bipedalism is a defining hominin characteristic and the knee obviously plays a central role in mammalian locomotion, so it is no surprise that palaeoanthropologists have paid particular attention to that joint. More specific to Sterkfontein, Berger and Tobias33 claimed that the anteroposterior convexity of the lateral condyle of StW 514a, a hominin proximal tibia from Member 4, as well as the fossil's lack of a 'meniscal notch' might indicate that (at least one) Australopithecus at the site locomoted in a 'chimpanzee-like' manner. However, a study that compared digital, three-dimensional surface areas of the lateral tibial condyles of various extant African hominoids, South African Australopithecus, and East African Australopithecus afarensis to two-dimensional surface area and arc and chord length measurements of the same feature concluded 'that tibial condylar curvature is a weak discriminator of locomotor variation in extant, and presumably fossil, hominoids'34(p.124). Further, Tardieu35 showed earlier that the lateral condyles of other early hominin tibiae - none of which would otherwise be characterised as 'chimpanzee-like' - also lack notches; more recently, the absence of lateral condylar notches has even been documented in some tibiae of fully bipedal modern humans.36 Zipfel and Berger37 acknowledged these observations in a study comparing StW 514a to another hominin proximal tibia from Sterkfontein Member 4, StW 396, but still emphasised, as did Berger and Tobias33(p.343) before them, that the semimembranosus attachment site of StW 514a 'forms a marked circular depression situated on the posteromedial margin of the condyle immediately below the medial condylar surface', implying that this enthesis morphology is distinctly chimpanzee-like. Our own observations of modern human tibiae disagree with this insinuation; in our experience, many modern human tibiae show large, strongly indented semimembranosus attachment sites with distinct borders, such as is illustrated in Figure 7. In sum, it thus seems that the StW 541a is not, in fact, particularly 'chimpanzee-like', but instead simply expresses a combined morphology that falls within the bounds of normal variation for hominin tibiae.
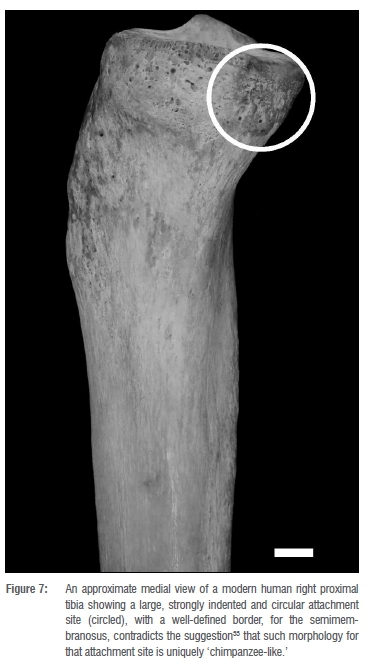
The same is true of StW 653, the new Member 4 hominin tibia described here, which compares quite favourably to not only other Australopithecus tibiae but also to those of modern humans. For example, the new fossil's strongly developed soleal line accords with those of fully bipedal modern humans. In no way does this suggest that StW 653 should be assigned to the genus Homo, but instead confirms the conclusions of others who question the efficacy of using certain morphological features to place early hominin tibia fossils into particular genera and/or species (see for example Dugan and Holliday36).
In contrast, the new StW 680 fibula from Member 4 differs markedly from those of modern humans and is instead more like those of extant African apes and also compares favourably to the Sterkfontein Australopithecus fibula fossils, StW 356 and StW 573. It possesses a generally triangular neck and has a high neck robusticity index (mediolateral diameter/ anteroposterior diameter *100)28 of 95.4, compared to a mean for two other Australopithecus proximal fibulae of 95.7 and a mean for 23 modern human fibulae of 81.8±16.6 (range = 52.6-111.5) (comparative data from Marchi et al.28). Moreover, the shape of the origin for the peroneus longus is convex, like those of other early hominin fibulae, including especially those attributed to Australopithecus, but unlike the origin on modern human fibulae.28
Jacovec fossils
As first noted by Clarke14, in possessing a relatively small head and long neck, StW 598 resembles SK 82 and SK 9730 - proximal femur fossils from Swartkrans (South Africa) that are usually assigned to Paranthropus robustus. StW 598 also resembles StW 99, a large femur specimen from Sterkfontein that is usually attributed to Australopithecus, but that - given its morphological continuity with the Swartkrans Paranthropus femur fossils and its possible origin from the Paranthropus-bearing Member 5 unit of the Sterkfontein Formation13 - is likely actually also Paranthropus (Table 1). With its long neck, StW 598 is, however, very similar to definitive Australopithecus femur specimens, StW 479 and StW 367, from Sterkfontein Member 4. StW 598 and StW 367 are, in fact, so similar that upon their cursory comparison, one could understandably conclude that the fossils are antimeres. Further detailed observations prove that conclusion erroneous, but the salient point made is that the morphology of StW 598 is not unique in the hominin fossil record. Adding to the comparative complexity, StW 522, a presumptive Australopithecus femur from Sterkfontein, shows a small head and short neck (Table 1).14
StW 598 has a remarkably high meric index of 90.6 (Table 1), compared to a mean meric index of 74.8±3.5 (range = 66.4-81.7) for 18 South and East African Australopithecus and Paranthropus femora.28 The cause(s) of femoral shaft shape remain hypothetical, but biomechanical explanations include reference to the influences of both the vasti and gluteal complexes. However, the meric index ranges of both knuckle-walking African apes (mean meric index of 42 Pan troglodytes femora = 85.0±5.5 [range = 71.1-95.5]; mean meric index of 47 Gorilla gorilla femora = 83.2±4.0 [range = 76.5-93.8]28) and terrestrial bipedal modern humans (mean meric index of 195 Homo sapiens femora = 80.8±6.8 [range = 56.1-96.6]28) not only overlap but also encompass the index value of StW 598. Thus, we are currently reluctant to extrapolate any functional interpretations of the fossil based on its round diaphyseal cross-section.
Likewise, although the position and narrow, ridge-like form of StW 598's gluteal enthesis suggest a relatively humanlike insertion of its gluteal musculature38, without a preserved greater trochanter, it would be unwise to place too much explanatory emphasis on that muscle scar. Indeed, in many other features, StW 598 is quite dissimilar to the proximal femora of modern humans. For instance, its anteroposteriorly compressed, superoinferiorly tall, and strongly anteverted neck is unlike the femoral necks of extant H. sapiens and, instead, mirrors the morphology of nearly every other known Australopithecus femur specimen.28,39
Moving to the distal femur, the femora of bipedal hominins are/were adapted, via significant anterior projection (i.e. 'elevation') of the lateral patellar lip, to resist dislocation of the patella under the load of stance phase valgus. A simple way to visually assess the functional elevation of the lateral patellar lip of a femur is to view the specimen distally, with an
axis running through the condylar meniscal grooves oriented horizontal to the flat base over which the femur is positioned.40 Unfortunately, because of the fragmentary nature of the fossil, the meniscal axis of StW 619 cannot be estimated with precision. However, placing the medial patellar surface of the specimen horizontally in distal view 'corresponds closely to the orientation recommended by Lovejoy40,41(p1232999-1) (Figure 8). This view clearly illustrates the appreciable depth of StW 619's patellar groove, as well as the significant functional elevation of its lateral patellar lip. Similarly, even though StW 619 lacks a discernible medial condylar boss, when the specimen is rotated into an estimated position of full extension and viewed distally, the anterior termination of the medial condyle assumes an ovoid shape. Lovejoy40 emphasises that this shape, in this view, is an ancillary indication that the tibial contact area of a femoral condyle was elongated relative to those of the femora of quadrupedal mammals. Bipedal primates with such elongated, ellipsoid femoral condyles possess(ed) knees that are 'tibial dominant'.40
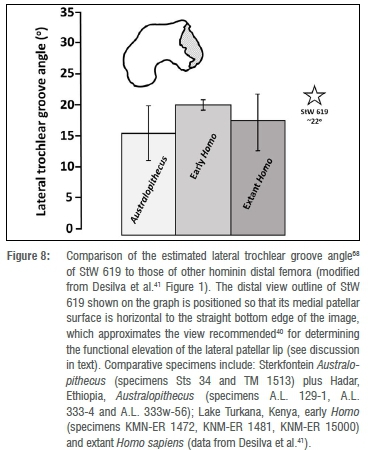
The conclusion that the knee of the StW 619 hominin - although not an exact morphological match for a modern human knee - was nonetheless capable of full, modern humanlike bipedal extension, is not unexpected given a broader view of the hominin fossil sample from Jacovec. As discussed above, the proximal femur StW 598 shows derived features, such as an elongated neck, a low neck-shaft angle and a relatively small head, observed in other proven bipedal hominins.38,41-48 In addition, Pickering et al.7 (see also Partridge et al.14) have also described two lumbar vertebrae from Jacovec that indicate the hominin(s) from which they derived possessed intrinsic lumbar lordosis. Lumbar lordosis is, of course, a critical component of hominid bipedality30, as it functions to maintain orthogrady by ameliorating strain on the dorsal spinal ligaments and by absorbing shock emanating from upright activities on terrestrial substrates49-51.
None of this is to suggest that all of the Jacovec hominins were necessarily as fully committed to terrestrial bipedalism as are modern humans. The StW 605 manual proximal phalanx from Jacovec is too damaged to quantify its included angle but it appears fairly curved longitudinally.8 Consensus posits a causal link between curved manual proximal phalanges and significant degrees of arboreal behaviour in primates.52,53 Similarly, the StW 606 hominin clavicle from Jacovec shows a mix of modern humanlike and apelike features, the latter of which would have endowed the hominin from which it derived with good climbing abilities.7,14
Conclusion
When added to results from previous studies of Australopithecus postcranial samples from Sterkfontein5,7,8,30,54-59, this study corroborates that those samples include functionally heterogeneous mixes of elements, some of which indicate postural and locomotor behaviours that are apelike and others of which indicate human-like adaptations. Starting from this basic understanding, there is, in our opinion, a fundamental problem that underlies many ensuing debates over the postcranial functional morphology of Australopithecus. Specifically, we are troubled by the fact that many disputants in these debates misunderstand or ignore the completely salient possibility (likelihood?) that the Sterkfontein Member 4 hominin postcranial collection samples at least two coeval species, A. africanus and A. prometheus, as is the case for the large craniodental sample of hominin fossils from that depositional unit.5,6,17-19 For instance, Harmon60 detected significant variation in the shape of proximal femora typically assigned to A. africanus, which she considered intraspecific variation, but that might actually be an interspecific difference. In contrast, Clarke5 demonstrated that two hominin first metatarsals from Sterkfontein Member 4 show distinct morphologies indicative of different modes of locomotion and thus concluded that the fossils likely represent separate species (see also Deloison61). Additionally, Kibii and Clarke57 suggested that the pelves of the partial Sterkfontein Member 4 skeletons, Sts 14 and StW 431, do not necessarily sample the same species. This issue has been clarified recently with the cleaning and reconstruction of the StW 573 skeleton of a female A. prometheus from Sterkfontein Member 262, which has a pelvis of similar size and morphology as that of the StW 431 male skeleton. Thus, it seems that StW 573 and StW 431 represent, respectively, female and male A. prometheus, while the much smaller female pelvis, Sts 14, seems to be that of A. africanus. In addition, the femur of A. prometheus, as exemplified by StW 573, possesses a short neck joined to a platymeric diaphysis63, leading to the logical conclusion that the Jacovec femur, StW 598, with a long neck and rounded diaphysis (as described above), probably represents A. africanus. Last, both these femoral forms differ from presumptive Paranthropus femora from Swartkrans and that of StW 99, from Sterkfontein Member 4.14,30 Collectively, these results seem to indicate significant interspecific hominin postcranial morphological variability in late Pliocene and early Pleistocene South Africa.
Acknowledgements
Our work was facilitated by a Kellet Mid-Career Award from the University of Wisconsin-Madison to T.R.P. and by a National Research Foundation (South Africa) grant to D.S., and by many years of support from the Palaeontological Scientific Trust (South Africa). In addition, we thank Kathleen Kuman for her consistent financial support of our research through National Research Foundation grants. Kuman also continues to provide logistical and personal support of our research, as do our colleagues, Andrew Phaswana and Abel Molepolle. Thanks to Matt Caruana for general suppor t of our work and especially for some images used in this paper. We thank Bernhard Zipfel for his interest in our research and for facilitating our comparative work at the University of the Witwatersrand. Likewise, we owe Marion Bamford and the Evolutionary Studies Institute at Wits much gratitude for supporting our research at Sterkfontein. Last, we are especially grateful to Jane Carruthers and Margaret Avery for their forbearance and level-headed advice as they shepherded this paper to publication.
Competing interests
We declare that there are no competing interests.
Authors' contributions
T.R.P. conceived and designed the study, collected and analysed the data, and wrote the manuscript. J.L.H. conceived and designed the study and collected and analysed the data. R.J.C. conceived and designed the study and collected and analysed the data. D.S. collected and analysed the data. A.J.H. collected and analysed the data.
References
1. Broom R, Schepers GWH. The South African fossil ape-men: The Australopithecinae. Pretoria: Transvaal Museum; 1946. [ Links ]
2. Tobias PV Copley K, Brain CK. South Africa. In: Oakley K, Campbell B, Molleson TI, editors. Catalogue of fossil hominids, part I: Africa (revised edition). London: British Museum of Natural History; 1977. p. 95-151. [ Links ]
3. Brain CK. The hunters or the hunted? An introduction to African cave taphonomy. Chicago, IL: University of Chicago Press; 1981. [ Links ]
4. Pickering TR, Clarke RJ, Moggi-Cecchi J. Role of carnivores in the accumulation of the Sterkfontein Member 4 hominid assemblage: A taphonomic reassessment of the complete hominid fossil sample (19361999). Am J Phys Anthropol. 2004;125(1):1-15. https://doi.org/10.1002/ajpa.10278 [ Links ]
5. Clarke RJ. Australopithecus from Sterkfontein Caves, South Africa. In: Reed K, Fleagle J, Leakey REF, editors. Paleobiology of Australopithecus. Dordrecht: Springer; 2013. p. 105-123. [ Links ]
6. Heaton JL, Pickering TR, Clarke RJ. Description and initial analysis of hominin craniodental remains from Sterkfontein Members 4-5 and Jacovec Cavern, South Africa (1998-2017). J Hum Evol. In press 2021. [ Links ]
7. Pickering TR, Heaton JL, Clarke RJ, Stratford D. Hominin vertebrae and upper limb bone fossils from Sterkfontein Caves, South Africa (1998-2003 excavations). Am J Phys Anthropol. 2018;3(168):459-l80. https://doi.org/10.1002/ajpa.23758 [ Links ]
8. Pickering TR, Heaton JL, Clarke RJ, Stratford D. Hominin hand bone fossils from Sterkfontein Caves, South Africa (1998-2003 excavations). J Hum Evol. 2018;1(118):89-102. https://doi.org/10.1016/j.jhevol.2018.02.014 [ Links ]
9. Vrba ES. The significance of bovid remains as indicators of environment and predation patterns. In: Behrensmeyer AK, Hill AFP editors. Fossils in the making. Chicago, IL: University of Chicago Press; 1980. p. 247-271. [ Links ]
10. Vrba ES. Biostratigraphy and chronology, based on Bovidae, of southern hominid-associated assemblages: Makapansgat, Sterkfontein, Taung, Kromdraai, Swartkrans; also Elandsfontein (Saldanha), Broken Hill (now Kabwe) and Cave of Hearths. Congres international de paleontologie humaine, 1er congres, tome 2. Nice: CRNS; 1982. [ Links ]
11. Delson E. Chronology of South African australopith site units. In: Grine F, editor. Evolutionary history of the 'robust' australopithecines. New York: Aldine de Gruyter; 1988. p. 317-324. [ Links ]
12. McKee JK, Thackeray JF, Berger LR. Faunal seriation of southern African Pliocene and Pleistocene fossil deposits. Am J Phys Anthropol. 1995;3(96):235-250. https://doi.org/10.1002/ajpa.1330960303 [ Links ]
13. Kuman K, Clarke RJ. Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5. J Hum Evol. 2000;6(38):827-847. https://doi.org/10.1006/jhev.1999.0392 [ Links ]
14. Partridge T, Granger DE, Caffee MW, Clarke RJ. Lower Pliocene hominid remains from Sterkfontein. Science. 2003;300:607-612. [ Links ]
15. Mavuso S. The sedimentology of the Jacovec Cavern, Sterkfontein [MSc thesis]. Johannesburg: University of the Witwatersrand; 2018. [ Links ]
16. Kibii J. Comparative taxonomic, taphonomic and palaeoenvironmental analysis of 4-2.3 million-year-old australopithecine cave infills at Sterkfontein [PhD thesis]. Johannesburg: University of the Witwatersrand; 2006. [ Links ]
17. Clarke R. Australopithecus and early Homo in southern Africa. In: Delson E, editor. Ancestors: The hard evidence. New York: Alan R. Liss; 1985. p. 171-177. [ Links ]
18. Clarke R. Advances in understanding the craniofacial anatomy of South African early hominids. In: Corruccini R, Ciochon R, editors. Integrative paths to the past: Essays in honor of F. Clark Howell. Upper Saddle River, NJ: Prentice Hall; 1994. p. 205-222. [ Links ]
19. Clarke R, Kuman K. The skull of StW 573, a 3.67 Ma Australopithecus prometheus skeleton from Sterkfontein Caves, South Africa. J Hum Evol. 2019;134:102634. https://doi.org/10.1016/j.jhevol.2019.06.005 [ Links ]
20. Behrensmeyer AK. Taphonomic and ecologic information from bone weathering. Paleobiology. 1978;4(2):150-162. https://doi.org/10.1017/S0094837300005820 [ Links ]
21. Blumenschine RJ, Marean CW, Capaldo S. Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces. J Archaeol Sci. 1996;23(4):493-507. https://doi.org/10.1006/jasc.1996.0047 [ Links ]
22. Villa P, Mahieu E. Breakage patterns of human long bones. J Hum Evol. 1991;21(1):27-48. https://doi.org/10.1016/0047-2484(91)90034-S [ Links ]
23. Pickering TR, Domínguez-Rodrigo M, Egeland CP Brain CK. The contribution of limb bone fracture patterns to reconstructing early hominid behaviour at Swartkrans Cave (South Africa): Archaeological application of a new analytical method. Int J Osteoarchaeol. 2005;15(4):247-260. https://doi.org/10.1002/oa.780 [ Links ]
24. Alcantara Garcia V Barba Egido R, Barral del Pino JM, Crespo Ruiz A, Eiriz Vidal A, Falquina Aparicio Á, et al. Determinación de procesos de fractura sobre huesos frescos: un sistema de análisis de los ángulos de los planos de fracturación como discriminador de agentes bióticos [Determination of fracture processes on fresh bones: A system for analyzing the angles of the fracturing planes as a discriminator of biotic agents]. Trabajos de Prehistoria. 2006;63:37-45. Spanish. [ Links ]
25. Martin R. Lehrbuch der Anthropologie in Systematischer Darstellung mit Besonderer Berücksichtigung der Anthropologischen Methoden für Studierende, Ärzte und Forschungsreisend [Textbook of anthropology in a systematic representation with special consideration of anthropological methods for students, doctors and explorers]. Vol. 2. Stuttgart: Gustav Fischer Verlag; 1928. German. [ Links ]
26. Martin R, Saller K. Lehrbuch der Anthropologie in Systematischer Darstellung mit Besonderer Berücksichtigung der Anthropologischen Methoden für Studierende, Ärzte und Forschungsreisend [Textbook of anthropology in a systematic representation with special consideration of anthropological methods for students, doctors and explorers]. Vol. 1. Stuttgart: Gustav Fischer Verlag; no date. German. [ Links ]
27. McHenry HM, Corruccini RS. The femur in early human evolution. Am J Phys Anthropol. 1978;49(4):473-488. https://doi.org/10.1002/ajpa.1330490407 [ Links ]
28. Marchi D, Walker CS, Wei P Holliday TW, Churchill S, Berger LR, et al. The thigh and leg of Homo naledi. J Hum Evol. 2017;104:174-204. https://doi.org/10.1016/j.jhevol.2016.09.005 [ Links ]
29. Holtby J. Some indices and measurements of the modern femur. J Anat. 1918;52:363-382. [ Links ]
30. Robinson JT. Early hominid posture and locomotion. Chicago, IL: University of Chicago; 1972. [ Links ]
31. Tardieu C. Morpho-functional analysis of the articular surfaces of the knee joint in primates. In: Chiarelli SB, Corruccini RS, editors. Primate evolutionary biology. Berlin: Springer-Verlag; 1981. p. 68-80. [ Links ]
32. Pickering TR, Heaton JL, Zwodeski S, Kuman K. Taphonomy of bones from baboons killed and eaten by wild leopards in Mapungubwe National Park, South Africa. J Taphonomy. 2011;9:117-159. [ Links ]
33. Berger LR, Tobias PV. A chimpanzee-like tibia from Sterkfontein, South Africa and its implications for the interpretation of bipedalism in Australopithecus africanus. J Hum Evol. 1996;30(4):343-348. https://doi.org/10.1006/jhev.1996.0027 [ Links ]
34. Organ J, Ward CV. Contours of the hominoid lateral tibial condyle with implications for Australopithecus. J Hum Evol. 2006;51(2):113-127. https://doi.org/10.1016/j.jhevol.2006.01.007 [ Links ]
35. Tardieu C. The knee joint in three hominoid primates: Application to Plio-Pleistocene hominids and evolutionary implications. In: Taub D, King F, editors. Current perspectives in primate biology. New York: Van Nostrand; 1986. p. 182-190. [ Links ]
36. Dugan J, Holliday TW. Utility of the lateral meniscal notch in distinguishing hominin taxa. J Hum Evol. 2009;57(6):773-776. https://doi.org/10.1016/j.jhevol.2009.07.006 [ Links ]
37. Zipfel B, Berger LR. Partial hominin tibia (StW 396) from Sterkfontein, South Africa. Palaeontol Afr. 2009;44:71-75. [ Links ]
38. Lovejoy CO, Meindl R, Ohman J, Heiple K, White TD. The Maka femur and its bearing on the antiquity of human walking: applying contemporary concepts of morphogenesis to the human fossil record. Am J Phys Anthropol. 2002;119(2):97-133. https://doi.org/10.1002/ajpa.10111 [ Links ]
39. Ruff C, Higgins R. Femoral neck structure and function in early hominins. Am J Phys Anthropol. 2013;150(4):512-525. https://doi.org/10.1002/ajpa.22214 [ Links ]
40. Lovejoy CO. The natural history of human gait and posture: Part 3, the knee. Gait & Posture. 2007;25(3):325-341. https://doi.org/10.1016/j.gaitpost.2006.05.001 [ Links ]
41. DeSilva J, Holt K, Churchill S, Carlson K, Walker C, Zipfel B, et al. The lower limb and mechanics of walking in Australopithecus sediba. Science. 2013;340:1232999-1-5. https://doi.org/10.1126/science.1232999 [ Links ]
42. Heiple K, Lovejoy CO. The distal femoral anatomy of Australopithecus. Am J Phys Anthropol. 1971;35:75-84. https://doi.org/10.1038/235175a0 [ Links ]
43. Lovejoy CO, Johanson DC, Coppens Y Hominid lower limb bones recovered from the Hadar formation: 1974-1977 collections. Am J Phys Anthropol. 1982;57(4):679-700. https://doi.org/10.1002/ajpa.1330570411 [ Links ]
44. Jungers WL. Lucy's limbs: Skeletal allometry and locomotion in Australopithecus afarensis. Nature. 1982;297:676-678. https://doi.org/10.1038/297676a0 [ Links ]
45. Stern JT, Susman RL. The locomotor anatomy of Australopithecus afarensis. Am J Phys Anthropol. 1983;60:279-317. [ Links ]
46. Harmon EH. The shape of the early hominin proximal femur. Am J Phys Anthropol. 2009;139(2):154-171. https://doi.org/10.1002/ajpa.20966 [ Links ]
47. Lovejoy CO. The natural history of human gait and posture: part 1, spine and pelvis. Gait & Posture. 2005;21(1):95-112. https://doi.org/10.1016/s0966-6362(04)00014-1 [ Links ]
48. White TD, WoldeGabriel G, Asfaw B, Ambrose S, Beyene Y Bernor RL, et al. Asa Issie, Aramis and the origin of Australopithecus. Nature. 2006;440:883-889. https://doi.org/10.1038/nature04629 [ Links ]
49. Lovejoy CO, McCollum M. Spinopelvic pathways to bipedality: why no hominids ever relied on a bent-hip-bent-knee gait. Philos Trans R Soc Lond. 2010;365:3289-3299. https://doi.org/10.1098/rstb.2010.0112 [ Links ]
50. Gracovetsky SA, Iacono S. Energy transfer in the spinal cord. J Biomed Engineer. 1987;9:99-114. [ Links ]
51. Vialle R, Levassor N, Rillardon L, Templier A, Skalli W, Guigui P. Radiographic analysis of the sagittal alignment and balance of the spine in asymptomatic subjects. J Bone Joint Surg A. 2005;87:260-267. [ Links ]
52. Susman RL. Comparative and functional morphology of hominoid fingers. Am J Phys Anthropol. 1979;50(2):215-236. https://doi.org/10.1002/ajpa.1330500211 [ Links ]
53. Rein TR. The correspondence between proximal phalanx morphology and locomotion: Implications for inferring the locomotor behavior of fossil catarrhines. Am J Phys Anthropol. 2011;46(3):435-445. https://doi.org/10.1002/ajpa.21599 [ Links ]
54. Vrba ES. A new study of the scapula of Australopithecus africanus from Sterkfontein. Am J Phys Anthropol. 1979;51:117-130. [ Links ]
55. Clarke RJ, Tobias PV. Sterkfontein Member 2 foot bones of the oldest South African hominid. Science. 1995;269:521-524. https://doi.org/10.1126/science.7624772 [ Links ]
56. Macchiarelli R, Bondioli L, Galichon V Tobias PV. Hip bone trabecular architecture shows uniquely distinctive locomotor behaviour in South African australopithecines. J Hum Evol. 1999;36(2):211-232. https://doi.org/10.1006/jhev.1998.0267 [ Links ]
57. Kibii JM, Clarke RJ. A reconstruction of the StW 431 Australopithecus pelvis based on newly discovered fragments. S Afr J Sci. 2003;99:225-226. [ Links ]
58. Haeusler M. New insights into the locomotion of Australopithecus africanus based on the pelvis. Evol Anthropol Suppl. 2002;1(S1):53-57. https://doi.org/10.1002/evan.10056 [ Links ]
59. McHenry HM, Berger LR. Body proportions in Australopithecus afarensis and A. africanus and the origins of the genus Homo. J Hum Evol. 1998;35:1-22. [ Links ]
60. Harmon E. Size and shape variation in the proximal femur of Australopithecus africanus. J Hum Evol. 2009;56(6):551-559. https://doi.org/10.1016/j.jhevol.2009.01.002 [ Links ]
61. Deloison Y Anatomie des os fossils de pieds des hominids d'Afrique du sud dates entre 2.4 et 3.5 millions d'annés: interpretation quant ä leur mode de locomotion [Anatomy of the fossil bones of the feet of South African hominids between 2.4 and 3.5 million years ago: Interpretation of their mode of locomotion]. Biometrie Hum Anthropol. 2003;21:189-230. French. [ Links ]
62. Clarke R. Excavation, reconstruction and taphonomy of the StW 573 Australopithecus prometheus skeleton from Sterkfontein Caves, South Africa. J Hum Evol. 2019;127:41-53. https://doi.org/10.1016/j.jhevol.2018.11.010 [ Links ]
63. Heaton J, Pickering TR, Carlson K, Crompton R, Jashashvili T, Beaudet A, et al. The long limb bones of the StW 573 Australopithecus skeleton from Sterkfontein Member 2: Descriptions and proportions. J Hum Evol. 2019;133:167-197. https://doi.org/10.1016/j.jhevol.2019.05.015 [ Links ]
64. Reynolds SC, Kibii JM. Sterkfontein at 75: Review of palaeoenvironments, fauna and archaeology from the hominin site of Sterkfontein (Gauteng Province, South Africa). Palaeontol Afr. 2011;46:59-88. [ Links ]
65. Clarke RJ. A deeper understanding of the stratigraphy of the Sterkfontein fossil hominid site. Trans R Soc S Afr. 2006;61(2):11-120. https://doi.org/10.1080/00359190609519960 [ Links ]
66. Robinson JT. Australopithecines and artefacts at Sterkfontein, part I: Sterkfontein stratigraphy and the significance of the Extension Site. S Afr Archaeol Bull. 1962;17(66):87-107. https://doi.org/10.2307/3886942 [ Links ]
67. Pickering TR, Heaton J, Clarke R, Sutton M, Brain CK, Kuman K. New hominid fossils from Member 1 of the Swartkrans Formation, South Africa. J Hum Evol. 2012;62(5):618-628. https://doi.org/10.1016/j.jhevol.2012.02.003 [ Links ]
68. Tardieu C, Glard Y Garron E, Boulay C, Jouve J-L, Dutour O, et al. Relationship between formation of the femoral bicondylar angle and trochlear shape: Independence of diaphyseal and epiphyseal growth. Am J Phys Anthropol. 2006;130(4):491-500. https://doi.org/10.1002/ajpa.20373 [ Links ]
 Correspondence:
Correspondence:
Travis Pickering
Email: travis.pickering@wisc.edu
Received: 26 July 2019
Revised: 01 July 2020
Accepted: 02 July 2020
Published: 29 Jan. 2021
Editor: Margaret Avery
Funding: University of Wisconsin-Madison, South African National Research Foundation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}