










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.116 no.11-12 Pretoria nov./dic. 2020
http://dx.doi.org/10.17159/sajs.2020/7916
REVIEW ARTICLE
The hunt for plant viruses in South Africa: The early days
Chrissie Rey
School of Molecular and Cell Biology University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Plant viruses cause widespread disease in agriculturally important crops, resulting in a reduction in both quality and quantity of produce. The introduction of intensive crop monoculture has resulted in an exponential increase in viral diseases which can cross over from wild indigenous plants. Viral pathogens also can occur in mixed infections, and rapid, sensitive and reliable diagnostic methods are required to identify and characterise the viruses responsible for the field diseases. In comparison to bacterial and fungal diseases, viral diseases are more difficult to diagnose. This review covers a period (1985-2011) in the history of virus discovery in South Africa during which several plant viruses from commercial and small-scale farms were identified and characterised. Interestingly, novel viruses were discovered in three crops, namely guar and cassava grown by small-scale farmers in Mpumalanga, and in commercial tobacco. The implication of these plant diseases is potential yield loss to farmers which can affect their livelihoods, and result in severe economic loss for the food and agriculture industries. Accurate identification of the causal viral agents of these viral diseases is a prerequisite for development of effective management strategies.
SIGNIFICANCE:
• This review provides a historical account of the discovery and characterisation of several viral pathogens of important agricultural crops grown by small-scale and commercial farmers in South Africa.
• Three novel plant viruses were isolated for the first time during the period (1985-2011) of this review.
Keywords: history of viruses, plant virus discovery, diagnostics, agricultural crops
Introduction
Origin, evolution and diversity of plant viruses
The history of virology interestingly began with the discovery of a novel infectious agent (tobacco mosaic virus), not in animals or humans, but in a tobacco plant.1 Viruses represent the most diverse, ubiquitous and numerous microorganisms defined to date. It has been speculated that viruses contributed to the origin of cellular life.2 Although the origin of viruses is not known due to lack of 'molecular fossil' information, extant evidence indicates a polyphyletic origin. As with human and animal viruses, plant viruses arose multiple times as hosts evolved and diverged in defined geographical regions. Plant viruses have a longstanding tight co-evolutionary history with their plant hosts, and while many of these do not cause disease, pathogenic viruses appear to dominate in economically important agricultural crops. Virus emergence is generally associated with ecological change or domestication of crops leading to intensive mono-agronomical practices. Complex ecological factors play a major role in plant virus emergence, host range expansion/diversification and plant-virus interactions.3 In nature, virus infections occur in multivirus-multihost communities, whereas in monoculture, although mixed infections do occur frequently, a single dominant virus species is usually associated with the disease. These viruses are transmitted by undiagnosed infected plant material or seeds introduced into geographical regions; or often these viruses transgress from wild plants in natural ecosystems into agri-systems, although from domestic crops into wild hosts can also occur.4 Virus populations are continuously evolving and adapting to new environments, vectors and hosts.5 Genetic diversity is achieved by several molecular mechanisms such as horizontal gene transfer, mutations (nucleotide substitutions), virus genome re-assortment or recombination. Virus populations behave as mutant spectra (quasispecies) composed of heterogeneous genetic variants around a master dominant sequence.6 Genetic diversity of a quasispecies at any given time is a result of natural selection and genetic drift, enabling viral emergence and altered pathogenesis. Factors determining the origin, emergence and diversification of virus populations in any specific geographical region/country are highly complex.
Diagnosis
Many viruses can remain undetected in the field, in particular those in wild plant hosts. Due to previous limitations of methods for virus detection and identification, many viruses remained 'hidden'. Early studies in the first six decades of plant virology (~1900-1960) were mainly focused on insect and mechanical transmission, centrifugation, serological assays and electron microscopy. The rise of molecular virology, including nucleic acid and protein technologies, has allowed for more rapid and accurate viral genome identification. Next-generation sequencing and metagenomics applied to plant virology in the last decade has provided rapid, efficient and high-throughput sequencing of DNA and RNA virus and viroid genomes.7 Metagenomic studies have also revealed a large diversity of viruses in wild plants.8 Next-generation sequencing combined with bioinformatics is also a powerful tool for de novo virus discovery and virus genome diversity studies. The potential contribution of minor genetic variants in a quasi-species to disease aetiology in the field is not yet known. Future next-generation sequencing studies on both temporal and spatial regulation of viral quasi-species in plant hosts merits further attention.9
A personal journey in plant virus hunting in southern Africa
To our knowledge, the first virus disease symptoms reported in South Africa was streak disease in maize.10 Maize streak disease was shown to be caused by a virus that is transmitted by a leafhopper vector.11 More recent studies have identified several ssRNA viruses occurring singly or in mixed infection in sweet potato in KwaZulu-Natal12, Limpopo13, and Eastern and Western Cape Provinces14. Several geminiviruses have also been detected in South Africa and southern African neighbouring counties causing serious yield reductions in cassava, maize, tomatoes, beans and sweet potatoes.15 Two monopartite begomovirus isolates, occurring either alone or in mixed infection in sweet potato (Ipomoea batatas) plants, were identified for the first time in South Africa in 2011 from samples near Louis Trichardt in the Limpopo Province.16 The complete genome sequence of one of the isolates corresponded to Sweet potato mosaic-associated virus (SPMaV; SPMaV-[ZA:WP:2011]), with which it shared 98.5% nucleotide identity. The second genome isolate sequence corresponded to a new variant of Sweet potato leaf curl Sao Paulo virus (SPLCSPV; SPLCSPV-[ZA:WP:2011]), with which it shared 91.4 % nucleotide identity.
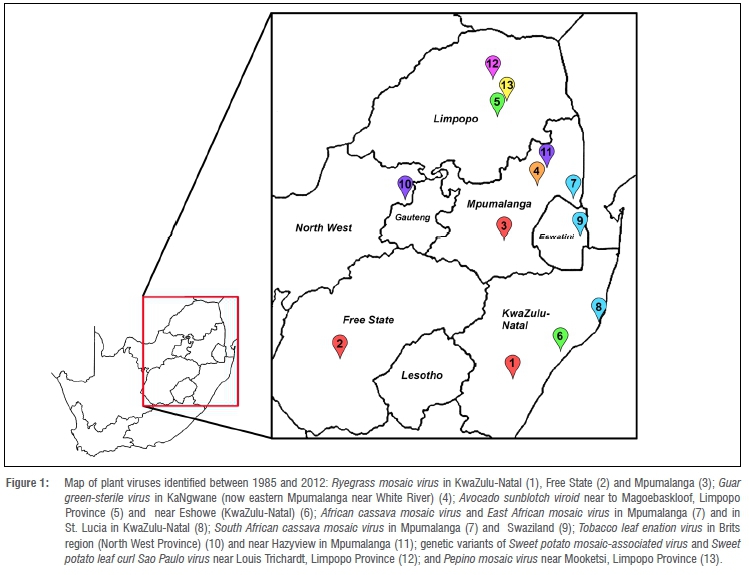
This review reports a number of new viruses or virus isolates that were identified in several provinces in South Africa between the years 1985 to 2011. These viruses are depicted in a geographical map (Figure 1).
A new green-sterile viral disease of guar in Mpumalanga
Guar (Cyamopsis tetragonoloba L.) is native to tropical Africa and Asia, and is grown in the USA, Pakistan, India and several countries in Africa as a livestock feedstuff.17 Guar meal has more recently also been used in poultry, and guar gum obtained from an annual pod is used as an emulsifier in baking mixes, cheeses, fats, oils, sauces and jams. The guar market in North America, Europe, Asia-Pacific, South America and Africa is projected to grow at a rate of 3.0% during the forecast period 2015-2024. Major importers of guar gum are Italy, South Africa, Russia, Australia, Netherlands, Japan, Brazil, Belgium and Canada. Guar was introduced into South Africa in the late 1940s and was grown by rural farmers in Mpumalanga and Northern Province.18 Guar was often used in intercropping with maize, cassava and groundnuts in Mpumalanga.
A number of potyviruses have been shown to infect guar, including peanut mottle potyvirus (PeMoV), Bean common mosaic virus (BCMV), and a symptomless seed-transmitted potyvirus from Indian, African and North American guar.19
While researching cassava mosaic disease in the Mpumalanga region, disease symptoms of reduced leaf size and number, and fewer, often sterile, inflorescences along the stem were observed on guar plants in the fields in KaNgwane (now incorporated into Mpumalanga). Often the stems remained green long after plant senescence, and 50% of the seeds were discoloured and distorted.18 The disease was named guar green sterile disease. It was suspected that this disease may be due to a potyvirus (ssRNA flexuous particles) and was named Guar green sterile virus (GGSV). Host range study in bean cultivars, and serological tests with antisera to Bean common mosaic virus (BCMV-SA) and Bean common mosaic necrosis virus (BCMNV), indicated that GGSV was serologically related. Further characterisation of guar green sterile disease was undertaken in order to elucidate the transmission, biological properties, and immunological relatedness to several other potyviruses. Mechanical inoculations on several indicator hosts18(Table1) resulted in symptoms of red vein necrosis, chlorosis and mosaic, while non-persistent aphid transmission on guar was not observed. Symptoms of leaf malformation and mosaic were observed on soybean (Glycine max) and several bean (Phaeseolus vulgaris L.) cultivars. Inability to obtain green sterile symptoms in guar was explained by the slow spread of the virus, low virus concentrations, and lack of symptoms in young guar plants. In the few cases in which local lesions were obtained, plants had a positive reaction to homologous antiserum raised against GGSV isolated from guar with green sterile symptoms. Seeds showing disease symptoms were positive when tested with BCMV antiserum, and antiserum raised to purified potyviruses extracted from guar leaves. Serological tests also demonstrated virus presence in seed coat and embryo tissue. Virus purification was only successful from fresh guar material collected from the field. Long flexuous particles averaging 750 X 15 nm were observed by transmission electron microscopy in leaf-dip preparations. Guar cv.TX-79-2741 was also strongly positive for Bean yellow mosaic virus (BYMV) antiserum. This study described a new disease symptom in guar, and indicated the putative involvement of a seed-transmitted legume potyvirus. A ssRNA shown to be 9.4 kb was isolated from purified GGSV virions, and Western blots showed the coat protein to be ~34 kD.20 The nucleotide sequence of the 3'-terminal region (1359 nt) of GGSV and translated amino acid was determined and compared with the latent seedborne guar symptomless potyvirus in guar imported into the USA from India.21 The presence of the amino acid triplet DAG in both GGSV and guar-US was consistent with aphid transmissibility. The coat protein amino acid sequence was 96% similar between GGSV and BCMV-NL4. GGSV and guar-US formed a tight cluster that was most closely related to the BCMV sub-group of potyviruses based on the coat protein and 3' UTR, strongly suggesting that GSV-US and GGSV are strains of BCMV-NL4.
Potyviruses represent one-quarter of known plant RNA viruses, and exhibit high nucleotide variation.22 This is an indication that potyviruses have an exceptional capacity to adapt to new hosts, vectors and environments. Further surveys of guar may demonstrate other potyvirus infections, and this could potentially have a negative impact on the use of this valuable crop, not only in the food and livestock industries but also in pharmacotherapy, where guar gum has more recently been used in the treatment of gut disorders.
The challenge of routine avocado sunblotch viroid detection in avocado trees
The avocado is one of the most important fruits in South African and global subtropical industries, and the local industry has increased rapidly in South Africa over the years.23 Avocado production in South Africa has traditionally been concentrated in the warm subtropical areas of the Limpopo and Mpumalanga Provinces in the northeast of the country between latitudes 22°S and 25°S. However, due to growing global demand and to produce year round, production is expanding in KwaZulu-Natal and the Eastern and Western Cape Provinces (up to 33°S).24 Avocado sunblotch is a serious disease of avocado (Persea americana Miller) worldwide. The disease affects both the yield and quality of the fruit. Fruit yield due to avocado sunblotch disease was reported to be reduced by 27.3% in cultivar Fuerte in the early 1980s in South Africa.25 The causal agent of sunblotch disease was first identified as a viroid (ASBVd)26, and is a low molecular mass circular ssRNA with a compact secondary structure comprising 247 nucleotides.
Sunblotch disease can be detected in avocado trees by identifying the typical symptoms in fruits; however, this approach is not applicable to infected asymptomatic trees.27 Diagnosis based on symptoms is not reliable and other sensitive diagnostic techniques are necessary to determine the health status of an avocado tree. Despite rapid diagnostic methods developed for viroid detection in the early 1980s, such as polyacrylamide gel electrophoresis (PAGE) and use of 32P-labelled complementary DNA probes28, these techniques were not reliable due to variability in viroid levels within branches and leaves. Problems with PAGE indexing for ASBVd were also reported in South Africa.29 Furthermore, contaminating polyphenols and polysaccharides in plant RNA extracts posed problems in RNA purification and gel separation. In order to improve screening for ASBVd, we developed a modified method to separate the viroid from other contaminating RNA species and contaminating plant compounds.30 Furthermore, we compared sensitivity of a cDNA versus a synthetic oligonucleotide probe for detection of ASBVd in plant extracts. From our study we showed that DNase treatment, and removal of polysaccharides, polyphenols, 4S and 5S ribosomal RNA by a modified method of CF-11 cellulose chromatography, improved hybridisation efficiency by 100-fold. The use of cDNA probes appeared to be a more reliable method for ASBVd-RNA detection compared to PAGE; however, the costs at the time of this study were high, and diagnosis laboratories were not equipped to handle cDNA techniques. The procedure was also very time intensive.
Other molecular techniques have been developed more recently; for example, a highly sensitive novel SYBR green-based method based on real-time reverse transcription (RT)-PCR was reported.31 The RT-qPCR is 100 times more sensitive to ASBVd than conventional RT-PCR. Infected asymptomatic trees play an important role in the epidemiology of this disease, and avocado nurseries need to be certified to ensure they provide pathogen-free avocado material. Although there is no cure for infected trees, early detection and sanitation practices may have a significant impact on avoiding the spread of this pathogen.
First report, characterisation and phylogenetic justification of Ryegrass mosaic virus-South African isolate
Annual ryegrass (Lolium multiflorum) is an important forage crop in many parts of the world including South Africa, where it is grown as a cool season pasture species under irrigation by commercial farmers for intensive dairy, lamb and beef production.32 It is mainly cultivated in Gauteng, North West, Free State and Kwa-Zulu-Natal Provinces. Ryegrass mosaic virus (RGMV) is reported to infect only members of the Poaceae family, including ryegrass (Lolium perenne L. and L. multiflorum Lam.), bromegrass (Bromus) species and oats (Avena sativa L.).33Ryegrass mosaic virus belongs to the Potyviridae family, and is a flexuous filamentous particle, approximately 703 nm in length and 15 nm in diameter, and is transmitted by the mite vector Abaracus hystrix.33Symptoms of infection usually range from yellow to light green mosaic or streaking.34
While symptoms in ryegrass had been reported in South Africa, there was no information on the distribution, biology or strain of the virus in South Africa prior to 1989, and both the vector and causal agent of ryegrass mosaic disease had not been confirmed. From 1989 to 1990, a broad survey of viruses in pasture grasses was carried out in several provinces, and mechanical and vector transmission was performed.34 Transmission results showed that RGMV-SA was present at only three of the sites (Cedara, Nooitgedacht and Glen). Dot-blot and leaf press immunobinding assays, using anti-RMV-SA antiserum raised to purified RMV particles in rabbits, and electron microscopy confirmed the presence of RGMV.35|Fig.1) Vector identification (by Dr E Ueckermann at the Plant Protection and Research Institute, Pretoria) also confirmed the mite to be A. hystrix belonging to the Eriophyidae family. A further host range study was performed on several indicator hosts using both mechanical and vector transmission. Of the 15 plant species screened, RMV-SA was shown to be transmissible to all the tested cultivars of L. multiflorum as well as L. perenne, A. sativa, A. fatua, Dactylus glomerata, Bromis mollis and Festiuca arundinaceae. RGMV-SA was found to be a severe isolate as it also induced severe tissue necrosis in several hosts, including L. multiflorum, a similar result as shown in L. multiflorum cv.s22 by RGMV isolates from Wales.33 Transmission of RGMV-SA by A. hystrix was found to be in a semi-persistent manner. In order to study the relationship with other virus members of the Rymovirus and Potyvirus genera, antisera for various potyviruses were used in dot-blot immunobinding assays and indirect ELISA. Immunoblots probed with antisera raised to RGMV-SA, RGMV-W and RGMV-Ca from South Africa, Wales and Canada, respectively, were positive; however, antisera to potato virusY (potyvirus) and other rymoviruses [Hordeum mosaic virus (HorMV); Agropyron mosaic virus (AgMV); wheat streak mosaic virus] were negative. The coat protein from purified virus particle preparations was determined to be 32.1±0.52 kDa, and the size of RGMV-SA RNA extracted from purified virions was estimated to be 2.8 x 106 on 1% agarose gels.35 Further, molecular cloning and nucleotide sequencing of the 3'terminal end (2094 nt) of the first strain of a RGMV isolate was accomplished.36 Two putative polyprotein cleavage sites, Q/L and E/A, were found, both of which are novel amongst potyviruses.
Alignment of the amino acid sequence of RGMA-SA with other Rymovirus genus members showed limited identity with Potyvirus. These results indicated that RGMV-SA was a distinct virus genus Rymovirus within the Potyviridae family.37 Additionally, phylogenetic analyses of the rymovirus sequences revealed a distinct group of two clusters: RGMV, HorMV and AgMV in one group and Wheat streak mosaic virus (WSMV) and Brome streak mosaic virus (BrSMV) in the second group. Rymoviruses also clustered separately from Ipomovirus and Bymovirus genus members.37(Rg.1)
While no extensive studies on RGMV-SA have ensued in the years following its discovery and characterisation, breeding of this important forage crop is ongoing. Yield data from the L. multiflorum breeding programme at ARC-Cedara in KwaZulu-Natal Province provides substantial evidence of the benefits of breeding to improve yield.38 Because ryegrass mosaic disease can affect yields, it is important that ryegrass growers are aware of a potential problem and remain vigilant.
Unravelling the disease aetiology of tobacco leaf curl disease in southern Africa: Tobacco leaf enation virus, a novel field phytoreovirus in tobacco reported for the first time
Tobacco leaf curl disease occurs mainly in tropical and sub-tropical regions, but is also reported in temperate regions such as Japan and parts of Europe and USA. Tobacco leaf curl disease was first reported in the Netherlands East Indies in 1912; however, it is thought that a disease fitting the description of leaf curl was present in South Africa as early as 1902.39 Leaf curl in tobacco was once reported as the most destructive disease in East Africa, Zimbabwe and the North West Province of South Africa.39 Although the disease syndrome was attributed to a virus, named Tobacco leaf curl virus (TLCv)40, which was shown to be transmitted by a species of whitefly, only one attempt was successful in isolating geminivirus particles.41 Symptom variability in tobacco leaf curl disease, however, has been reported by several researchers worldwide, including in South Africa42, and while suspected to be caused by different geminivirus strains, this suspicion was never confirmed experimentally. Furthermore, repeated attempts to isolate a geminivirus from tobacco in South Africa had failed.43 Tobacco plants exhibiting leaf curl symptoms were collected between 1992 and 1995 from Zimbabwe and from the North West and Mpumalanga Provinces in South Africa. Three different leaf symptoms, based on type and severity, were distinguished and placed into three classes (I, II and III).44 Class I tobacco plants from South Africa predominantly exhibited stunting and stems appeared bent and irregular and the thickened leaves curled and twisted with thickened veins on the ventral surface and frilly enations along the midrib. Class II leaf curl symptoms were considered non-viral, and attributed to a physiological disorder. Transmission electron microscopy, dot-immunobinding assays and polyacrylamide gel electrophoresis (PAGE) failed to detect geminivirus particles or coat protein from symptomatic class I plants.45 DNA hybridisation with eight geminivirus-specific probes and PCR with three sets of geminivirus-specific degenerate primers failed to detect DNA. Interestingly, dsRNA extraction revealed 12 bands ranging from 4350 bp to 810 bp, which resembled plant reoviruses that are transmitted by leafhopper or planthopper vectors. Plant reovirus-like symptoms include enations, vein swellings and dwarfing.46Wound tumour phytoreovirus (WTV) was at the time the only plant reovirus known to infect dicotyledonous plants.47 As the dsRNA pattern and sizes and symptoms of class I leaf curl tobacco strongly suggested a phytoreovirus, further studies were performed to confirm this diagnosis. Reovirus extractions successfully isolated icosahedral particles with an outer core 60-65 nm in diameter and an inner core 40-15 nm in diameter. Twelve distinct non-polyadenylated dsRNAs were isolated from purified virions, and the total molecular masses of the dsRNAs ranged from 17.86 to 18.40 X 106 Da in polyacrylamide and agarose gels, respectively.48 Comparisons of the tobacco phytoreovirus from South Africa with other known phytoreoviruses (Maize rough dwarf fijivirus (MRDV), Garlic dwarf reovirus (GDV), Rice ragged stunt reovirus (RRSV), Rice black-streaked dwarf fijivirus (RBSDV), WTV and Rice dwarf phytoreovirus (RDV)) revealed a unique dsRNA banding pattern that was distinct but most similar to WTV, the type species of the genus Phytoreovirus. Hybridisations of WTV-cloned DNA probes (segments S4 and S6 to S9) and dsRNAs from infected tobacco indicated no significant sequence similarity, whereas an indirect ELISA with a polyclonal antiserum to WTV showed strong positive cross-reactivity to tobacco virions. Western blot analysis of structural viral proteins (apparent molecular weights of 93 kDa, 58 kDa, 48 kDa, 39 kDa and 36 kDa) associated with the dsRNAs isolated from infected tobacco in South Africa, suggested that these proteins may correspond to structural WTV-like proteins. The virus was named 'tobacco leaf enation virus' (TLEV).
To further establish the nature of TLEV-associated disease phenotype in tobacco, molecular characterisation of six dsRNA components was undertaken.49 Results of this study revealed the conserved terminal sequence: 5'GG(U/C)...UGAU 3' of segments S6-S12, while adjacent to these conserved terminal sequences are imperfect inverted repeats (7-15 bp in length), both features being common to reoviruses. The complete nucleotide sequences were determined for segments S5 (2610 bp), S7 (1740 bp), S8 (1439 bp), S10 (1252 bp), S11 (1187 bp) and S12 (836 bp). Comparison of full-length nucleotide sequences with corresponding segments of other phytoreoviruses, Rice gall dwarf virus (RGDV), Rice dwarf virus (RDV) and WTV has shown nucleotide and predicted amino acid identities within the range of 30-60%. TLEV consistently showed a higher identity to WTV than to other phytoreovirus species for which sequence data were available. Each segment had a single predicted open reading frame encoding proteins with calculated molecular weights of 90.6 kDa (S5), 58.1 kDa (S7), 47.7 kDa (S8), 39.8 kDa (S10); 35 kDa (S11) and 19.5 kDa (S12). The relatively low nucleotide and amino acid identity to other members of the genus confirmed that TLEV is a novel phytoreovirus, distinct from the only other reported dicotyledonous-infecting WTV and was the first report of a phytoreovirus in tobacco, and the first discovered reovirus on the African continent.
Discovery of Tobacco leaf curl Zimbabwe virus - a new distinct monopartite begomovirus associated with subgenomic defective DNA molecules
Class III leaf curl plants from Zimbabwe tested positive in PCR using two sets of geminivirus-specific degenerate primers that amplify the core region of the coat protein of DNA A or the bottom half of DNA A of most whitefly transmitted geminiviruses. Dot-blot hybridisation and triple antibody sandwich ELISAs for geminiviruses also were positive.50 The Zimbabwe virus isolates were named Tobacco leaf curl Zimbabwe virus (TbLCZWV). The complete DNA A sequence of TbLCZWV comprises 2767 nucleotides with six major open reading frames encoding proteins with molecular masses greater than 9 kDa.50 Agro-inoculation with a full-length TbLCZWV DNA A infectious clone resulted in systemic infection of tobacco and tomato. Symptoms of field-infected tobacco leaves (Nicotiana tabacum cv. HG) collected from Zimbabwe exhibited distinct downward leaf margin curling, vein thickening, wavy midribs and significantly distinct cup-shaped enations on the ventral surface. Symptoms observed in Nicotiana benthamiana, following TbLCZWV agro-inoculation, were stunting, severe upward leaf curling, leaf distortion, ventral vein thickening and bending of petioles (Figure 2a). In N. tabacum cv. Samsun, extensive puckering of dorsal leaf surfaces (Figure 2b) and upward leaf margin curling in newly developing leaves occurred (Figure 2c). N. tabacum cv. Samsun ventral leaf surfaces of infected plants exhibited vein thickenings on smaller veins of the infected plants that were absent in the healthy plants (Figure 2d).



Efforts to identify a DNA B component were unsuccessful. These findings suggested that TbLCZWV was a novel member of the monopartite group of begomoviruses, with its closest relative being Chayote mosaic virus (Figure 3). Abutting primer PCR amplified two ~1300-bp subgenomic defective DNA molecules originating from TbLCZWV DNA A. Many monopartite begomoviruses are associated with co-dependent betasatellites for successful infection.51 While a betasatellite was not found in N. tabacum cv. HG, it would prove interesting to revisit tobacco leaf curl disease in Zimbabwe. Since this discovery, a TbLCZWV isolate was reported from the Comoros archipelago.52 Further, a complex tobacco leaf curl begomovirus exhibiting differential disease phenotypes in the Comoros archipelago has recently been observed.53 Extensive genetic material exchange through recombination, molecular diversity and evolution of these begomovirus complexes warrants further research.

The elusive identification of South African cassava mosaic virus: A novel geminivirus provides further evidence for recombination
Cassava (Manihot esculenta Crantz) is one of the most important staple root crops whose starchy roots are a major source of dietary energy for more than 500 million people in tropical and sub-tropical regions and thus occupies a uniquely important position as a food security crop for smallholder farmers. In low-income areas of these countries, the crop is a staple food consumed by over 700 million people. Over the period 1980 to 2013, global cassava production has increased from 124 million tons to an estimated 263 million tons, and global fresh root production reached an estimate of 275.7 million tons in 2018.54 Cassava mosaic disease is considered to be the first virus disease reported in Africa55, where symptoms of leaf curl and mosaic were reported (Figure 2e). Cassava (Manihot esculenta Crantz) is thought to have been introduced, in two independent events, from the Americas onto the African continent in the 16th and 18th centuries56 and cultivation spread throughout sub-Saharan Africa. The introduction of this exotic crop into a new ecosystem provided opportunities for infection of cassava by geminiviruses from native plant species. The geminiviruses (family Geminiviridae) constitute the largest family of plant viruses, with over 450 distinct species that occur across all world regions with favourable climates for their insect vectors.57 Among the geminiviruses, begomoviruses (genus Begomovirus) are responsible for a large number of emergent crop diseases over the past 50 years throughout the tropical and subtropical regions worldwide.58 Begomoviruses are composed of circular ssDNA genomes, and can be monopartite (DNA A) or bipartite (DNA A and B). African cassava mosaic virus (ACMV) was the first reported begomovirus associated with cassava mosaic disease.59 Cassava mosaic viruses frequently occur in mixed synergistic infections, and readily exchange their genomes through recombination or re-assortment which leads to high genetic variability within species.60
It is speculated that cassava was introduced into South Africa from Zimbabwe, Mozambique and Mauritius, and was spread across the northern and eastern regions of Natal and Mpumalanga by Tsonga tribesman in the 1830s.61 Cassava is mainly cultivated by small, resource-poor farmers. A study in the early 1980s reported cassava mosaic disease symptoms in South Africa and Swaziland.61 While it is not known how cassava mosaic disease emerged in South Africa, it is speculated to have been introduced through infected cassava vegetative material via other neighbouring countries. An initial transmission electron microscope study to identify the causal agent of cassava mosaic disease in cassava in South Africa, and in mechanically inoculated indicator host N. benthamiana, was unable to detect typical geminivirus particles or inclusion bodies in the nucleus.62 However, using antiserum raised against ACMV-Kenya, immunogold labelling located geminiviral antigen in infected cassava leaf tissue. On the basis of a partial DNA A sequence, and comparative serological results with three monoclonal antibodies against EACMV, ACMV and Indian cassava mosaic virus (ICMV), a novel geminivirus was identified, South African cassava mosaic virus.63,64The individual common region and coat protein nucleotide sequences of SACMV clustered with the monopartite Tomato yellow leaf curl virus-Sardinia (TYLCV-SR), followed by East African cassava mosaic virus [Malawi-8N-2007] and East African cassava mosaic virus-Tanzania [Tanzania].64 Southern blots with three DIG-labelled ACMV-Kenya, ICMV and Tomato golden mosaic virus (TGMV) DNA B probes, and nucleotide sequencing of a 687-bp DNA B component, proved that SACMV was a bipartite begomovirus that clustered with the Old World sub-group.
Earlier serological surveys in west, central and east African countries suggested that cassava mosaic begomoviruses, ACMV, EACMV and the ACMV-EACMV recombinant (EACMV-Uganda) had separate non-overlapping distributions65; however, East African cassava mosaic virus-Cameroon [Cameroon-1998] was reported more west in Cameroon.66 A later PCR study using core coat protein and EACMV-Uganda primers in southern Africa (Angola, Zambia, Swaziland, Mozambique and Zimbabwe) demonstrated that the three viruses occurred in most of these countries, but EACMV-Uganda only occurred in mixed infections.67 All three viruses were present in South Africa, and further heteroduplex mobility assay profiling was able to distinguish four different virus species and 11 different strains/isolates.68 In addition to EACMV, ACMV, SACMV, EACMV-Uganda and East African cassava mosaic virus-Malawi were also detected. Full-length DNA A sequence comparisons with other begomoviruses available first demonstrated that SACMV was most closely related to EACMV-Tanzania and Malawi isolates (Figure 3). Later, full-length DNA A and B genomes were shown also to be closely related to EaCMV-Malawi and EACMV-Uganda mild and severe isolates, respectively.69 One significant recombination event spanning the entire AC4 open reading frame on DNA A was identified, and interestingly, this open reading frame clustered with a group consisting of ICMV, ACMV, Tomato leaf curl virus-Australia (TLCV-AU) and Tomato yellow leaf curl virus (TYLCV) isolates from Sardinia and Sicily.69 Following the discovery of SACMV in South Africa, Swaziland, Mozambique and Zimbabwe, it was later reported from Madagascar.70 Collectively, studies confirm that SACMV is most closely related to a clade that contains East African cassava mosaic virus, East African cassava mosaic virus-Tanzania, East African cassava mosaic virus-Cameroon, East African cassava mosaic virus-Uganda, East African cassava mosaic virus-Zanzibar, East African cassava virus-Malawi, and East African cassava mosaic virus-Kenya, which occur in east Africa. Evidence supports recombination events amongst African, Mediterranean and southwest Indian Ocean islands begomoviruses.71 More recently, three additional cassava begomovirus species, Cassava mosaic virus-Madagascar, East African cassava mosaic virus-Kenya and East African cassava mosaic virus-Zanzibar - have been discovered in cassava in sub-Saharan Africa. Two distinct species, Indian cassava mosaic virus and Sri Lankan cassava mosaic virus, are described from the Asian sub-continent.58,72
With growing human populations and drought associated with climate change predicted for the future, cassava can provide one solution for food security in South Africa, and in other countries in the SADC region. Cassava starch has other potential diversified industrial uses such as food additives, biofuels, biodegradable packaging and animal feedstock.73 Introduction of high-yielding varieties, improved pest and disease control and better processing methods could increase cassava production in Africa. Because begomoviruses are a major threat, genetic engineering for virus resistance through RNA interference and gene editing can provide critical solutions for virus disease management.
First report of Pepino mosaic virus infecting tomato in South Africa
Pepino mosaic virus (genus Potexvirus) is a highly infectious virus that is responsible for significant losses in yield of tomato fruit (Solanum lycopersicum) across Europe, Asia and the Americas. Pepino mosaic virus (PepMV) rapidly evolved from an emerging to an endemic pathogen in tomato crops worldwide.74 During the winter growing season of 2008, symptoms of uneven discolouration of tomato fruit and mosaic symptoms on leaves were detected in tomato farms in Limpopo Province. Double antibody-sandwich (DAS)-ELISA using polyclonal antibodies against PepMV (Agdia, Elkhart, IN, USA) confirmed PepMV in leaf and fruit samples. Mechanical inoculation of susceptible S. lycopersicum cv. Rooikhaki seedlings with infected sap was achieved, and all inoculated plants developed characteristic PepMV symptoms including leaf bubbling, distortion and curled leaves. A 986-bp region, that included the coat protein of the PepMV genome, was RT-PCR amplified and sequenced. Phylogenetic analysis clustered the South African sequence with EU (European), LP (Peruvian), US1 (United States)/CH1 (Chilean) and US2/CH2 isolates. This confirmed the first report of PepMV in South Africa.75
Concluding remarks
A number of diverse RNA and DNA viruses from many agriculturally important crops have been uncovered in South Africa over the past 11 decades. More recently, virus metagenomic surveys have been extended from agricultural systems to natural ecosystems. For example, a survey in 2010 in the fynbos in South Africa revealed a highly divergent geminivirus from wild spurge (Euphorbia caputmedusae), which is symptomless in this natural host but causes severe symptoms in N. benthamiana and tomato.76 Metagenomic studies will continue to reveal novel plant viruses in both wild plants and agricultural crops, and characterising the impact of viruses in natural ecosystems may lead to informed agricultural practices or alternative solutions to controlling these pathogens. One of the future challenges will be to understand the equilibrium between plant viruses and their hosts, and how the long term co-evolutionary balance between natural hosts and plant viruses can be broken by large-scale monoculture.9 While many useful molecular interaction studies have been performed in plant virus-model host systems, future studies of interactions in specific virus-host combinations in more complex ecosystems are warranted. Furthermore, next-generation sequencing and new sample preparation techniques will also allow researchers to sequence ancient viral genomes from archaic plant material, and shed more light on virus evolution.
Acknowledgements
I would like to thank the following funders that contributed to this research: Chemserve Steinhall (Pty) Ltd, the South African Avocado Growers' Association, ARC-Vegetable and Ornamental Plant Institute, ARC-Industrial Crops, African Products Pty Ltd (Anglo-American), the British Council, the Richard Ward Endowment Fund, the University of the Witwatersrand, and the Foundation for Research and Development (now the National Research Foundation).
I would also like to express my everlasting gratitude to Helen Garnett, my PhD supervisor and friend, who inspired me and allowed me the freedom to pursue my passion, and who encouraged me to be an independent thinker.
This review is an acknowledgement to all my Master's and PhD students who were involved in this virus discovery period (1985-2011) and many who joined me in an old Datsun 'bakkie' around South Africa and into the fields hunting for virus symptoms in plants. They are also co-authors on the research manuscripts cited in this historical review: Ashley Moses, Hedva Ben-Moshe, Sarah Salm, Anthony Smith, Shirley Johnson, S Khitsane Maria Paximadis, Gillian Dusterwald, Laura Duyver, Elvera D'Andrea, Jennifer Calvert-Evers, Anabela Picton, Shaun Berry, Leigh Berrie, Vincent Fondong, Debbie Carmichael, and Lindy Esterhuizen. I thank the research collaborators who also participated in this virus discovery adventure over these years: Brian Beck, Mbudzeni Sibara, Ed Rybicki, Hanu Pappu, Roy French, Judy Brown, Maureen Wolfson, Kenneth Palmer, Ken Mabasa, Guido Boccardo, Christian Potgieter, Schalk van Heerden, Henrietta van Heerden and Michele Cloete. I also mention other postgraduate students chasing viruses around South Africa, Mozambique, Tanzania and Nigeria, who gave me such joy: Angela Eni, Nurbibi Cossa, Jamisse Amisse, Valter Nuaila, Gration Rwegasira, Cyprian Rajabu, Julia Mulabisana and Thuli Nhlapo.
I would also like to thank the Agricultural Research Council (ARC) Research Institutes in Pretoria (ARC-Vegetable and Ornamental Plant Institute), Rustenburg (ARC-Industrial Crops) and Nelspruit (ARC-Tropical and Subtropical Crops) for their assistance and support over the years and the plant growers who welcomed us to the farms to examine and collect the plants.
Competing interests
I declare that there are no competing interests.
References
1. Creager AN, Scholthof KB, Citovsky V Scholthof HB. Tobacco mosaic virus. Pioneering research for a century. Plant Cell. 1999;11:301-308. https://doi.org/10.1105/tpc.11.3.301 [ Links ]
2. Moreira D, López-García P. Evolution of viruses and cells: Do we need a fourth domain of life to explain the origin of eukaryotes? Philos Trans R Soc B Biol Sci. 2015;370, Art. #20140327. https://doi.org/10.1098/rstb.2014.0327 [ Links ]
3. Roossinck MJ. Plants, viruses and the environment: Ecology and mutualism. Virology. 2015;479:271-277. https://doi.org/10.1016/j.virol.2015.03.041 [ Links ]
4. Roossinck MJ, García-Arenal F. Ecosystem simplification, biodiversity loss and plant virus emergence. Curr Opin Virol. 2015;10:56-62. https://doi.org/10.1016/j.coviro.2015.01.005 [ Links ]
5. Elena SF, Fraile A, García-Arenal F. Evolution and emergence of plant viruses. Adv Virus Res. 2014;88:161-191. https://doi.org/10.1016/B978-0-12-800098-4.00003-9 [ Links ]
6. Domingo E, Sheldon J, Perales C. Viral quasispecies evolution. Microbiol Mol Biol. 2012;76:159-216. https://doi.org/10.1128/MMBR.05023-11 [ Links ]
7. Hadidi A, Flores R, Candresse T, Barba M. Next-generation sequencing and genome editing in plant virology. Front Microbiol. 2016;7:1325. https://doi.org/10.3389/fmicb.2016.01325 [ Links ]
8. Roossinck MJ. Deep sequencing for discovery and evolutionary analysis of plant viruses. Virus Res. 2017;239:82-86. https://doi.org/10.1016/j.virusres.2016.11.019 [ Links ]
9. Sanfacon H. Grand challenge in plant virology: Understanding the impact of plant viruses in model plants, in agricultural crops and in complex ecosystems. Front Microbiol. 2017;8:360. https://doi.org/10.3389/fmicb.2017.00860 [ Links ]
10. Fuller C. Mealie variegation. In: 1st Report of the Government Entomologist, Natal,1899-1900. Pietermaritzburg: P Davis & Sons, Government Printers; 1901. [ Links ]
11. Storey HH. The transmission of streak disease of maize by the leafhopper Balclutha mbila Naude. Ann Appl Biol. 1925;12:422-139. https://doi.org/10.1111/j.1744-7348.1925.tb04238.x [ Links ]
12. Sivparsad BJ, Gubba A. Identification and distribution of viruses infecting sweet potato (Ipomoea batatas L.) in KwaZulu-Natal Province, South Africa. S Afr J Plant Soil. 2013;30(3):179-190. https://doi.org/10.1080/02571862.2013.854415 [ Links ]
13. Mulabisana J, Cloete M, Mabasa K, Laurie SM, Oelofse D, Rey MEC. Genetic characterisation of viruses infecting sweetpotato in South Africa. Acta Hortic. 2015;1105:155-162. https://doi.org/10.17660/ActaHortic.2015.1105.22 [ Links ]
14. Nhlapo TF, Rees DJG, Odeny DA, Mulabisana JM, Rey MEC. Viral metagenomics reveals sweet potato virus diversity in the Eastern and Western Cape provinces of South Africa. S Afr J Bot. 2019;117:256-267. https://doi.org/10.1016/j.sajb.2018.05.024 [ Links ]
15. Rey MEC, Ndunguru J, Berrie LC, Paximadis M, Berry S, Cossa N, et al. Diversity of dicotyledenous-infecting geminiviruses and their associated DNA molecules in southern Africa, including the south-west Indian Ocean islands. Viruses. 2012;4(9):1753-1791. https://doi.org/10.3390/v4091753 [ Links ]
16. Esterhuizen LL, Van Heerden SW, Rey MEC, Van Heerden H. Genetic identification of two sweet potato infecting begomoviruses in South Africa. Arch Virol. 2012;157:2241-2245. https://doi.org/10.1007/s00705-012-1398-4 [ Links ]
17. Hymowitz T. The trans-domestication concept as applied to guar. Econ Bot. 1972;26:49-60. https://doi.org/10.1007/BF02862261 [ Links ]
18. Ben Moshe H, Rey MEC, Sibara M, Garnett HM, Beck B. A new green sterile disease of guar in South Africa. Plant Dis. 1991;75:638-643. https://doi.org/10.1094/PD-75-0638 [ Links ]
19. Hansen AJ, Lesemann DE. Occurrence and characteristics of a seed-transmitted potyvirus from Indian, African and North-American guar. Phytopathology. 1978;68:841-846. https://doi.org/10.1094/Phyto-68-841 [ Links ]
20. Rey MEC, Johnson S, Smith A, Khitsane S. Preliminary characterization of a potyvirus associated with green sterile disease of guar. Plant Dis. 1995;79(2):172-176. https://doi.org/10.1094/PD-79-0172 [ Links ]
21. Gillespie AG, Pappu HR, Jain R, Rey MEC, Hopkins MS, Pinnow DL, et al. Characterization of a seedborne potyvirus of the Bean common mosaic virus subgroup occurring in guar, Cyamopsis tetragonoloba (L.) Taubert. Plant Dis. 1998;82:765-770. https://doi.org/10.1094/PDIS.1998.82.7.765 [ Links ]
22. Nigram D, LaTourrette K, Souza PFN, Garcia-Ruiz H. Genome-wide variation in potyviruses. Front Plant Sci. 2019;10:1439. https://doi.org/10.3389/fpls.2019.01439 [ Links ]
23. Zwane ZR, Gubba A, Jooste AEC. Review of avocado sunblotch disease. Sub-Tropical Quarterly Journal. 2017;19:32-36. [ Links ]
24. Donkin D. Overview of SA avocado industry [homepage on the Internet]. No date [cited 2020 Jan 31]. Available from: https://www.avocado.co.za [ Links ]
25. Da Graca JV Mason TE, Antel HJ. Effect of avocado sunblotch disease on fruit yield. South African Avocado Growers' Association Yearbook. 1983;6:86-87. [ Links ]
26. Palukaitis P, Hatta T, Alexander DM, Symons RH. Characterization of a viroid associated with avocado sunblotch disease. Virology. 1979;99:145. https://doi.org/10.1016/0042-6822(79)90045-X [ Links ]
27. Palukaitus P, Da Graca JV. Sunblotch-associated reduction in fruit yield in both symptomatic and symptomless carrier trees. South African Growers' Association Yearbook. 1985;8:59-60. [ Links ]
28. Bar-Joseph M, Yesodi V Franck A, Rosner A, Segev D. Recent experience with the use of synthetic DNA probes for the detection of avocado sunblotch viroid. South African Avocado Growers' Association Yearbook. 1986;9:75-77. [ Links ]
29. Da Graca JV Trench, TN. Problems with PAGE indexing for avocado sunblotch. South African Avocado Growers' Association Yearbook. 1985;8:61. [ Links ]
30. Rey MEC, Moses AV. Detection of avocado sunblotch viroid using cDNA and synthetic oligonucleotide probes. Phytophylactica. 1993;25:257-262. [ Links ]
31. Morey-León G, Ortega-Ramírez E, Julca-Chunga C, Santos-Chanta C, Graterol-Caldera L, Mialhe E. The detection of avocado sunblotch viroid in avocado using a real-time reverse transcriptase polymerase chain reaction. BioTechnologia. 2018;99(2):99-107. https://doi.org/10.5114/bta.2018.75653 [ Links ]
32. De Villiers JF, Vfen Ryssen JBJ. Performance responses of lambs of various ages to Italian ryegrass (Lolium multiflorum) fertilized with various levels of nitrogen. S Afr J Anim Sci. 2001;3:142-148. https://doi.org/10.4314/sajas.v31i3.3797 [ Links ]
33. Slykhuis JT, Paliwal YC. Ryegrass mosaic virus. Descriptions of plant viruses [webpage on the Internet]. [cited 2020 Jan 31]. Available from: www.dpvweb.net. [ Links ]
34. Salm SN, Rey MEC. Epidemiology of ryegrass mosaic virus in South Africa. J Grassl Soc South Afr. 1992;9(4):170-175. https://doi.org/10.1080/02566702.1992.9648319 [ Links ]
35. Salm SN, Rey MEC, Wolfson MM. A South African isolate of Ryegrass mosaic virus. Plant Pathol.1994;43:708-712. https://doi.org/10.1111/j.1365-3059.1994.tb01610.x [ Links ]
36. Salm SN, Rey MEC, French R. Molecular cloning and nucleotide sequencing of the 3' terminus of a South African strain of RgMV and in vitro expression of the coat protein gene. Arch Virol. 1996;141:185-195. https://doi.org/10.1007/BF01718600 [ Links ]
37. Salm SN, Rey MEC, Rybicki EP Phylogenetic justification for splitting the Rymovirus genus of the taxonomic family Potyviridae. Arch Virol. 1996;141:2237-2242. https://doi.org/10.1007/BF01718229 [ Links ]
38. Ammann SB, Smith A, Goodenough DCW. Pasture plant breeding in South Africa: Lessons from the past and future needs. Grassland Society of Southern Africa Congress 50; 2015 July 20-23; Pietermaritzburg, South Africa. [ Links ]
39. Lucas GB. 1975 Diseases of tobacco. Raleigh, NC: Biological Consulting Associates USA; 1975. [ Links ]
40. Storey HH. A new disease of the tobacco plant. Nature. 1931;128:187-188. https://doi.org/10.1038/128187b0 [ Links ]
41. Osaki T, Inouye T. Tobacco leaf curl virus. CMI/AAB Descriptions of Plant Wuses. 1981;no. 232. [ Links ]
42. McClean ADP Some leaf-curl diseases in South Africa. Part I. Leaf-curl disease of tobacco. Science Bulletin, Department of Agriculture and Forestry, South Africa. 1940;225:7-60. [ Links ]
43. Thatcher J. Whitefly transmission and ecology of tobacco leaf curl disease [master's dissertaion]. Pietermaritzburg: University of Natal; 1976. [ Links ]
44. Paximadis M, Rey MEC. Aetiology of tobacco leaf curl in southern Africa. Ann Appl Biol. 1997;131:449-457. https://doi.org/10.1111/j.1744-7348.1997.tb05172.x [ Links ]
45. Paximadis M, Dusterwald G, Duyver L, Rey MEC. Preliminary investigation of the causal agent(s) of a disease causing leaf curl of tobacco in South Africa. Plant Pathol. 1997;46:282-290. https://doi.org/10.1046/j.1365-3059.1997.d01-226.x [ Links ]
46. Francki RIB, Boccardo G. The plant Reoviridae. In: Joklik WK, editor. The Reoviridae. New York: Plenum Press; 1983. p. 505-563. https://doi.org/10.1007/978-1-4899-0580-2_10 [ Links ]
47. Omura T. Genomes and primary protein structures of phytoreoviruses. Semin Virol. 1995;6:97-102. https://doi.org/10.1006/smvy.1995.0012 [ Links ]
48. Rey MEC, D'Andrea E, Calvert-Evers J, Paximadis M, Boccardo G. Evidence for a phytoreovirus associated with tobacco exhibiting leaf curl symptoms in South Africa. Phytopathology. 1999;89:303-307. https://doi.org/10.1094/PHYTO.1999.89.4.303 [ Links ]
49. Picton A, Potgieter C, Rey MEC. Molecular analysis of six segments of Tobacco leaf enation virus, a novel phytoreovirus from tobacco. Virus Genes. 2007;35:387-393. https://doi.org/10.1007/s11262-007-0088-x [ Links ]
50. Paximadis M, Rey MEC. Genome organization of Tobacco leaf curl virus from Zimbabwe, a new distinct monopartite begomovirus associated with subgenomic defective DNA molecules. J Gen Virol. 2001;82:3091-3097. https://doi.org/10.1099/0022-1317-82-12-3091 [ Links ]
51. Briddon R, Ghabrial W, Lin SNS, Palukaitis P, Scholthof KBG, Vetten HJ. Satellites and other virus-dependent nucleic acids. In: King AMQ, Lefkowitz E, Adams MJ, Carstens EB, editors. Virus taxonomy - Ninth Report of the International Committee on Taxonomy of Viruses. New York: New York City Associated Press; 2012. p. 1209-1219. [ Links ]
52. Lett JM, Lefeuvre F, Naze F, Delatte H, Mohamed-Ali Y, Reynaud B. First report of Tobacco leaf curl Zimbabwe virus affecting tobacco in the Comoros Archipelago. Plant Pathol. 2006;55(4):567. https://doi.org/10.1111/j.1365-3059.2006.01421.x [ Links ]
53. Thierry M, Lefeuvre P, Hoareau M, Perefarres F, Delatte H, Reynaud B, et al. Differential disease phenotype of begomoviruses associated with tobacco leaf curl disease in Comoros. Arch Virol. 2012;157:545-550. https://doi.org/10.1007/s00705-011-1199-1 [ Links ]
54. Food and Agriculture Organization (FAO). Food outlook - Biannual report on global food markets. Cassava market developments and outlook. Rome: FAO; 2018. p.12-23. [ Links ]
55. Warburg O. Die kulturpflanzen usambaras [The crops of Usambara]. Mittcilungenaus den Deutschen Schutzgebieten. 1894;7:131. German. [ Links ]
56. Fauquet C, Fargette D. African cassava mosaic virus: Etiology, epidemiology and control. Plant Dis. 1990;74:404-411. https://doi.org/10.1094/PD-74-0404 [ Links ]
57. Zerbini FM, Briddon RW, Idris A, Martin DP Moriones E, Navas-Castillo J, et al. ICTV virus taxonomy profile: Geminiviridae. J Gen Virol. 2017;98:131-133. https://doi.org/10.1099/jgv.0.000738 [ Links ]
58. Brown JK, Zerbini FM, Navas-Castillo J, Moriones E, Ramos-Sobrinho R, Silva JC, et al. Revision of begomovirus taxonomy based on pairwise sequence comparisons. Arch Virol. 2015;160:1593-1619. https://doi.org/10.1007/s00705-015-2398-y [ Links ]
59. Stanley J, Gay MR. Nucleotide sequence of cassava latent virus DNA. Nature. 1983;301:260-262. https://doi.org/10.1038/301260a0 [ Links ]
60. Fondong VN, Pita JS, Rey MEC, de Kochko A, Beachy RN, Fauquet CM. Evidence of synergism between African cassava mosaic virus and a new double-recombinant geminivirus infecting cassava in Cameroon. J Gen Virol. 2000;81(1):287-297. https://doi.org/10.1099/0022-1317-81-1-287 [ Links ]
61. Trench TN, Martin MM, Hemmes EA. An assessment of cassava African mosaic disease in South Africa and Swaziland. S Afr J Plant Soil. 1985;2(3):169-170. https://doi.org/10.1080/02571862.1985.10634163 [ Links ]
62. Rey MEC. Immunocytochemical techniques in the investigation of the causal agent of cassava mosaic disease in southern Africa. Micron Microscopica Acta. 1987;18:307-316. https://doi.org/10.1016/0739-6260(87)90031-1 [ Links ]
63. Berrie LC, Palmer K, Rybicki EP, Hiyadat SH, Maxwell DP, Rey MEC. A new isolate of African cassava mosaic virus in South Africa. Afr J Root Tuber Crops. 1997;2:49-52. [ Links ]
64. Berrie LC, Palmer K, Rybicki EP, Rey MEC. Molecular characterization of a distinct South African cassava infecting geminivirus. Arch Virol. 1998;143:2253-2260. https://doi.org/10.1007/s007050050457 [ Links ]
65. Swanson MM, Harrison BD. Properties, relationships and distribution of cassava mosaic viruses. Trop Sci.1994;34:15-25. [ Links ]
66. Fondong VN, Pita JS, Rey MEC, Beachy RN, Fauquet CM. First report of the presence of East African cassava mosaic virus in Cameroon. Plant Dis. 1998;82(10):1172. https://doi.org/10.1094/PDIS.1998.82.10.1172B [ Links ]
67. Berry S, Rey MEC. Molecular evidence for the existence of biodiverse begomovirus populations in cassava in southern Africa. Arch Virol. 2001;146:1795-1802. https://doi.org/10.1007/s007050170065 [ Links ]
68. Berry S, Rey MEC. Differentiation of cassava-infecting begomoviruses using heteroduplex mobility assays. J Virol Methods. 2001;92:151-163. https://doi.org/10.1016/S0166-0934(00)00286-X [ Links ]
69. Berrie LC, Rybicki EP, Rey MEC. Complete nucleotide sequence of South African cassava mosaic virus: Further evidence for recombination among begomoviruses. J Gen Virol. 2001;82:53-58. https://doi.org/10.1099/0022-1317-82-1-53 [ Links ]
70. De Bruyn A, Harimalala M, Zinga I, Mabvakure BM, Hoareau H, Ravigne V, et al. Divergent evolutionary and epidemiological dynamics of cassava mosaic geminiviruses in Madagascar. BMC Evol Biol. 2016;16:182. https://doi.org/10.1186/s12862-016-0749-2 [ Links ]
71. Lefeuvre P Martin DP Hoareau M, Naze F, Delatte HM, Thierry A, et al. Begomovirus 'melting pot' in the south-west Indian Ocean islands: Molecular diversity and evolution through recombination. J Gen Virol. 2017;88(12):3458-3468. https://doi.org/10.1099/vir.0.83252-0 [ Links ]
72. Rey C, Vanderschuren H. Cassava mosaic and brown streak diseases: Current perspectives and beyond. Annu Rev Virol. 2017;4:429-452. https://doi.org/10.1146/annurev-virology-101416-041913 [ Links ]
73. Baguma Y Nuwamanya E, Rey C. The African perspective: Developing an African bio-resource based industry: The case for cassava. In: Virgin I, Morris JE, editors. Creating sustainable bioeconomies: The bioscience revolution in Europe and Africa. London: Routledge Publishers; 2016. https://doi.org/10.4324/9781315745183 [ Links ]
74. Hanssen IM, Thomma BPHJ. Pepino mosaic virus: A successful pathogen that rapidly evolved from emerging to endemic in tomato crops. Mol Plant Pathol. 2010;11:179-189. https://doi.org/10.1111/j.1364-3703.2009.00600.x [ Links ]
75. Carmichael DJ, Rey MEC, Naidoo S, Cook G, Van Heerden SW. First report of Pepino mosaic virus infecting tomato in South Africa. Plant Dis. 2011;95:767. https://doi.org/10.1094/PDIS-01-11-0036 [ Links ]
76. Bernardo P Golden M, Akram M, Naimuddin, Nadarajan N, Fernandez E, et al. Identification and characterisation of a highly divergent geminivirus: Evolutionary and taxonomic implications. Virus Res. 2013;177:35-45. https://doi.org/10.1016/j.virusres.2013.07.006 [ Links ]
 Correspondence:
Correspondence:
Chrissie Rey
Email: chrissie.rey@wits.ac.za
Received: 31 Jan. 2020
Accepted: 02 Apr. 2020
Published: 26 Nov. 2020
Editors: Teresa Coutinho, Salmina Mokgehle ©
Funding: Chemserve Steinhall (Pty) Ltd; South African Avocado Growers' Association; ARC-Vegetable and Ornamental Plant Institute; ARC-Industrial Crops; African Products Pty Ltd (Anglo-American); British Council; Richard Ward Endowment Fund, University of the Witwatersrand; National Research Foundation (South Africa)


{kind=link}