










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.116 no.5-6 Pretoria Mai./Jun. 2020
http://dx.doi.org/10.17159/sajs.2020/6754
RESEARCH ARTICLE
Large mammal exploitation during the c. 14-11 ka Oakhurst techno-complex at Klipdrift Cave, South Africa
Emmanuel DiscampsI; Christopher S. HenshilwoodII, III; Karen L. van NiekerkII
ICNRS UMR5608 TRACES, University of Toulouse Jean Jaurès, Toulouse, France
IISFF Centre for Early Sapiens Behaviour, University of Bergen, Bergen, Norway
IIIEvolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Understanding how hunter-gatherers adapted to the marked environmental changes of the last glacial-interglacial transition (~18 to 11.7 ka cal. BP) remains a key question for archaeologists. South Africa, with its rich and well-preserved archaeological sequences, has a major role to play in this study. Reconstructing the subsistence strategies of people during the Later Stone Age (LSA) is crucial for investigating human-environment interactions at this period in South Africa, yet data are scarce. Recent excavations at a new LSA site, Klipdrift Cave, in the southern Cape, revealed c. 14-11 ka levels with excellent faunal preservation associated with an Oakhurst lithic industry. Taphonomic and zooarchaeological analyses of these levels show an almost exclusive accumulation of large mammal remains by LSA groups, with evidence of meat removal, marrow extraction, fire use and the preferential import of nutritious elements back to the site. Large mammals from the site indicate a relatively stable environment dominated by open grasslands that is in accordance with isotopic analyses, with only subtle diachronic variability. Comparison of faunal dynamics with changes in lithic industries, shellfish density and composition reflects complex, asynchronous changes in the macromammal, micromammal, shellfish and lithic records throughout the Oakhurst levels. Rather than evidence of a strong impact of global climate change, Klipdrift Cave shows subtle shifts in subsistence patterns and technology that are better explained by internal societal dynamics and the history of the Oakhurst techno-complex, or local changes in site occupation and direct environment.
SIGNIFICANCE:
• LSA archaeological sequences can document the impact of the marked environmental changes of the Pleistocene-Holocene transition on hunter-gatherer societies. Studies of past subsistence strategies are central to our understanding of human-environment interactions in these contexts.
• Zooarchaeological, taphonomical and palaeoecological analyses of the large mammal remains from the excavated LSA sequence at Klipdrift Cave provide new data on these interactions. The data highlight asynchronous changes in subsistence patterns, lithic technology and local environment, supporting a complex interplay between climate change, local environment, societal changes and human prehistory.
• Klipdrift Cave data set also shows that excavation and analytical choices can strongly bias faunal analysis and environmental reconstructions based thereon.
Keywords: zooarchaeology, Later Stone Age, southern Cape, South Africa, palaeoenvironment
Introduction
Archaeologists have long tried to understand how hunter-gatherers adapted to the marked environmental changes that characterised the last glacial-interglacial transition from 18 to 11.7 ka cal. BP internationally and in South Africa.1 Such studies are hindered by the scarcity of reliable data on the subsistence strategies of human populations during this period named the Later Stone Age (LSA) in southern Africa. In 2010 and 2011, we (C.S.H. and K.L.v.N.) excavated a new site located in the southern Cape: Klipdrift Cave. Faunal preservation at the site is excellent, and the remains of a number of taxa were excavated, of which shellfish and tortoise are the most abundant,2-4 together with remains of dune mole rat, hyrax, micromammals, fish, birds (including ostriches and raptors) and large mammals. The latter are unpublished and form the focus of this study. Zooarchaeological analysis of large mammal remains allows us to provide new data on subsistence strategies and faunal exploitation patterns by LSA hunter-gatherers during the c. 14-11 ka Oakhurst lithic techno-complex in the southern Cape, showing that environmental change around the Late Pleistocene-Holocene transition cannot be simplistically linked to changes in subsistence and technology.
Material and methods
Klipdrift Cave site and stratigraphy
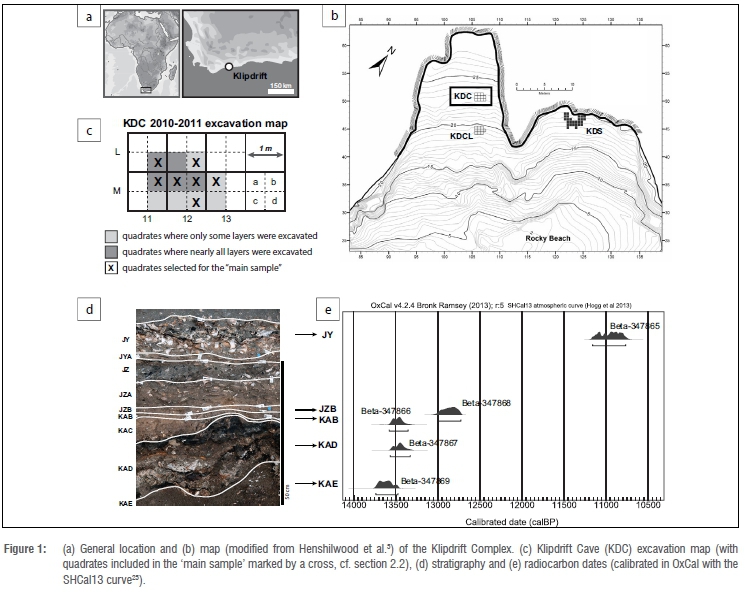
The Klipdrift Complex (34°27.0963'S, 20°43.4582'E), located on the coast of the southern Cape in the De Hoop Nature Reserve (Figure 1a), comprises three sectors (Figure 1b): Klipdrift Shelter (KDS, with Middle Stone Age c. 66-59 ka Howiesons Poort deposits), Klipdrift Cave (KDC) and Klipdrift Cave Lower (KDCL c. 70 ka). KDC, the focus of this paper, consists of several superimposed LSA layers containing hearths, stone and bone artefacts, ostrich eggshell beads, ochre and abundant faunal remains (macro- and micromammals, shellfish, fish, tortoise and bird). KDC, set in a steep quartzite cliff is c. 17 m above sea level and next to a rocky shoreline with few sandy beaches. Detailed information about the site context are available in Ryano et al.3 and Henshilwood et al.5
KDC was excavated in 2010 and 2011 over an area of 2.75 m2 (Figure 1c). Excavation was done by brush and trowel in 50x50-cm quadrates, and a Trimble VX Total Station was used to plot key artefacts and features. KDC layers are named, from top to bottom, JY JYA, JZ, JZA, JZB, KAB, KAC, KAD and KAE (Figure 1d). Five accelerator mass spectrometry radiocarbon dates on charcoal samples place the site's occupations between 13.8 and 10.7 ka cal. BP (Figure 1e). Typo-technological analysis identified a homogeneous lithic industry pertaining to the early phases of the Oakhurst lithic techno-complex.2,3
Faunal sample
All plotted remains of large mammals (i.e. larger than hyrax) were included in this study, as well as fragments of large mammals from coarse fractions (sieve residues) from a selection of quadrates, hereafter referred to as 'main sample' (Figure 1c). This main sample includes material from quadrates at the centre of the excavation (all layers from quadrates L11d, M11b, M12a, M12b) and others for key layers (L12d for layer JZ, M12d and M13a for layer KAD). Remains of large mammals (including long bone shaft fragments) from the coarse fraction of the main sample that could be identified anatomically were isolated and allocated a unique ID number. For the key layers JZ and KAD, this represents the sorting of coarse fractions from about 75% of the excavated volume, and, for other layers, about 50% of the excavated volume. The extended sample adds to the main sample by including plotted bones from all the other quadrates. Thus, main sample = plotted remains and coarse fractions from quadrates L11d, M11b, M12a, M12b (all layers), L12d (layer JZ only), M12d and M13a (layer KAD only); extended sample = main sample + plotted remains from other quadrates (Figure 1c).
Analysis of large mammal remains
Faunal remains were identified to skeletal element, taxon and/or size classes (adapted from Brain6; Table 1), using primarily the Ditsong Museum comparative collections, as well as skeletons acquired and processed by the rangers of the De Hoop Nature Reserve and E.D. Each fragment was entered in a FileMaker database after observation of its surface under low-angled light with a 30x hand lens and a stereomicroscope when necessary. Bone surface modifications pertinent to assessing the agent that accumulated the remains were coded (cut marks, percussion marks, tooth marks, digested bones and degree of burning7-9) for the 950 remains, including 270 long bone fragments. Fracture patterns on long bones were recorded as recent, green or dry break following criteria developed by Villa and Mahieu10. Skeletal-part representation was assessed using NNISR or normed number of identified specimens.11
To best compare proportions of taxonomic groups between layers of very different sample size, we computed adjusted Wald proportions and confidence intervals.12 The graphical representation of proportions with 95% confidence intervals allows for quick and efficient comparison while considering biases due to small sample sizes. Chi-square tests and Spearman correlations between variables were performed using the RAST software suite.13
Results
Taxonomie identifications
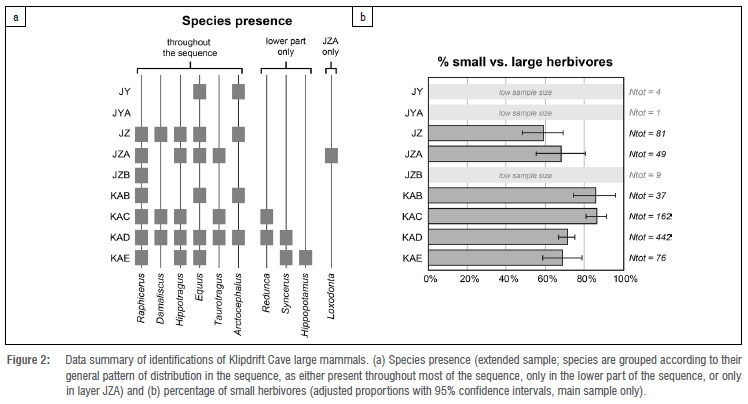
Of the 950 remains entered in the database (886 from the main sample), 101 could be identified beyond a size class attribution (Table 1). Carnivores are rare, with only five remains of Cape fur seal and four of unidentified small carnivores (potentially caracal and a smaller felid). Herbivores, notably bovids, dominate. Most are grazers that are often found in open grassland/savanna ecosystems. No strong diachronic patterning is evident in terms of species present throughout the sequence (Figure 2a). Even if some variations exist (e.g. hippopotamus, southern reedbuck and buffalo are only present in the lower part of the sequence - layers KAC to KAE - while elephant is present only in JZA), most might best be explained by sample size.
Comparisons between the main sample (that includes both plotted and coarse fraction remains) and the extended sample (only plotted remains from other quadrates) shows strong differences: large herbivores are considerably over-represented when only plotted remains are included in the analysis (70.3%, instead of 25.7% for the main sample; Table 2), a highly significant difference (chi-square = 63.743; p<0.001). This is to be expected considering that the larger remains of large herbivores have more chance of being plotted during excavation, distorting faunal compositions.14 Considering this bias, subsequent analyses in this study only consider the main sample.
Percentages by size classes show a slight but statistically supported diachronic pattern: the proportion of small herbivores (compared to large herbivores) is about 70% in layers KAD and KAE, increases slightly in layers KAB and KAC to about 85%, before reducing to about 60% in layer JZ (Figure 2b). Both these trends are statistically significant (between KAD and KAC: chi-square = 15.937, p<0.001; between KAB and JZA: chi-square = 4.80, p<0.05).
Taphonomic observations
Bone surfaces are generally well preserved at KDC. Anthropic action on bones is evident (Table 3) in the form of cut marks (6.7%, n=57), percussion marks (4.3%, n=38, including percussion striae, notches, bone flakes and peeling fractures of ribs) and, most notably, burnt bones (24.8% of the remains entered in the database for the main sample, n=203, without counting the thousands of small unidentifiable fragments in the coarse fractions). The assemblage is highly fragmented, with only 6.1% (n=51) being complete or sub-complete bones, mostly small ones (e.g. carpals, tarsals, sesamoids, third phalanges and caudal vertebras). Long bone shafts have fracture morphologies characteristic of green-bone breakage in 95% of cases (n=113 out of 119 diagnostic fractures; Table 3). Conversely, only two bones have evidence of digestion and one bone has tooth marks. None of the carnivores documented (Table 1) could be responsible for the KDC accumulation of large mammals (seal and small carnivores are rarely responsible for accumulating terrestrial large mammal remains). All these data point to a nearly exclusive accumulation of large mammal remains by LSA humans in KDC, with evidence of meat removal, marrow extraction and fire use.
Skeletal-part profiles
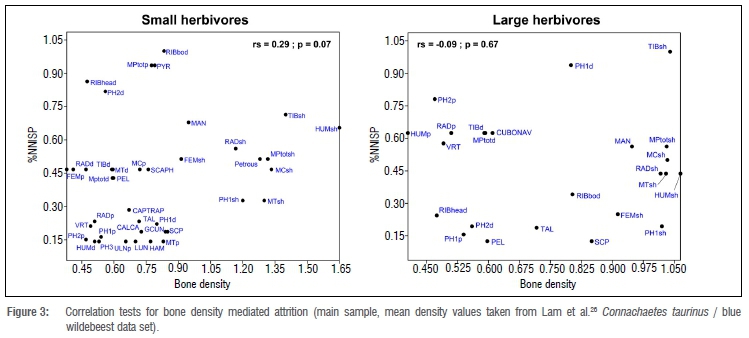
The small sample sizes per taxa and layer preclude detailed analysis of skeletal-part profiles. Transport strategies are thus generalised by merging all layers together in the main sample, and only distinguishing between small and large herbivores (Table 4, Figures 3 and 4). Nearly all elements were found in KDC, but in varying abundance. Elements rich in marrow or meat are over-represented (long bones), while the less nutritious short bones (carpals, tarsals, sesamoids and phalanges) are rare. Correlations between skeletal element abundance (expressed as %NNISP) and bone density are not statistically significant (p>0.05 for both small and large herbivores, Figure 3). This absence of correlation, together with the fact that bone surfaces are excellently preserved and that several foetal bones were found in the bone assemblage, point to a limited impact of post-depositional processes on body-part profiles. Skeletal-part profiles could reflect human transport decisions (i.e. preferential import of nutritious elements to KDC) and bone-processing techniques. With respect to the latter, the scarcity of carpals and tarsals - bones that are relatively robust and easily identifiable - is problematic. Considering the abundance of metapodials, tibia and radial bones (those that are anatomically connected to carpals and tarsals), the most parsimonious explanation is that short bones were brought back to KDC, but later destroyed by human activity, for example, by burning, whether intentional or not. This hypothesis is supported by the high percentage of carpals and tarsals that are burnt (44.4%).

When body part data are analysed by size classes (Figure 4), skeletal-part profiles appear more balanced for small herbivores (i.e. relatively comparable proportions of skull, axial and long bones) than for large ones. Complete carcasses of small bovids might have been brought back to KDC more often, while bigger prey was introduced in the form of carcass segments.
This possibility is notably evident in the relative abundance of small herbivore ribs but not those of large herbivores. Conversely, large herbivore tibiae, one of the bones with the richest marrow yield in African bovids,15 are over-represented, suggesting transport choices favouring nutrition-rich parts.
Discussion and conclusion
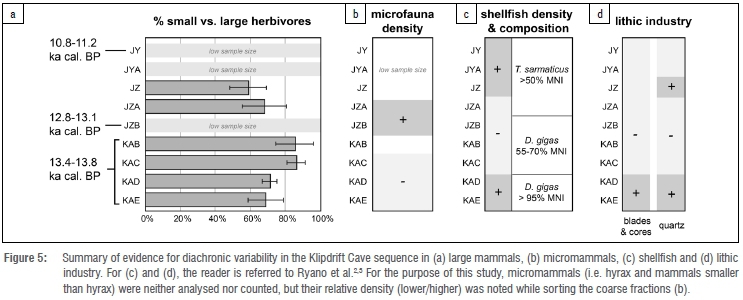
Analysis of Klipdrift Cave large mammal remains from the c. 14-11 ka Oakhurst levels reflects a relatively stable environment, most probably dominated by open grasslands. This finding accords with previous isotopic analyses that highlight little to no variation in the oxygen and carbon composition of KDC ostrich eggshells through time.16 Subtle variations are, however, observed in the relative proportions of small and large herbivores (Figure 5a). While sorting the coarse fractions for this study, variations in the abundance of micromammals was also noted (Figure 5b). Changes in the lithic industry and in shellfish density and composition were previously described.2,3 When summarised, these data sets reflect complex, asynchronous changes in the macromammal, micromammal, shellfish and lithic records throughout the Oakhurst levels (Figure 5).
Of specific interest are the two bottom layers, KAD and KAE. Their distinguishing features, compared with the overlying KAB and KAC layers, are a higher proportion of large herbivores, greater shellfish density and larger proportion of Dinoplaxgigas (giant chiton), as well as more blades, cores and quartz lithic elements. The radiocarbon dates obtained for these four layers overlap (Figure 1e), hence emphasising that change was rapid. These changes occurred around 13.5 ka cal. BP both after and before periods of major environmental changes documented at the last glacial-interglacial transition in the southern Cape (i.e. the two humid episodes described by Chase et al.17 are placed around 15.4-14.2 ka and 11.8-10.7 ka). This shift in subsistence patterns and technology is likely linked to the internal human dynamics and history of the Oakhurst techno-complex, and/or to local changes in site occupation and direct environment, and not to the influence of major climate change. The sample sizes for the upper layers of KDC are too low to allow for an investigation of potential shifts in large mammal exploitation at the onset of the Holocene, when major environmental changes are documented in the southern Cape.1
In other LSA sequences such as Boomplaas18,19, Byneskranskop 120 and Nelson Bay Cave21, an important faunal shift is identified at around 12 ka22 with a marked decrease of grazers (notably equids and alcephalines), replaced by smaller bovids (such as mountain reedbuck, grey rhebok, klipspringer and grysbok/steenbok). This change from a grass dominated environment to one in which bush, forest and fynbos increase has been correlated with the transition from the Robberg to the Oakhurst, suggests that the hunting of smaller antelopes was one driver for technological change.23,24 KDC shows an example of early Oakhurst layers associated with a predominance of large grazing herbivores, that weakens the presumed links between environmental and technological changes. Furthermore, KDC data indicate that the integration or exclusion of remains from coarse fractions in a faunal analysis can drastically alter the proportion of large identified herbivores. Excavation and/or analytical choices (e.g. if only plotted specimens are included) may strongly bias faunal lists and environmental reconstructions.14
KDC large mammal remains are well preserved and show strong evidence of anthropic accumulation with little to no contribution by carnivores.
However, the small sample size per species limits zooarchaeological analyses. Preliminary data suggest that KDC was used by LSA people as a site to which they preferentially brought nutritious-rich carcass segments, especially those of large herbivores, considering that smaller herbivore prey may have been brought to the site as complete carcasses. Future excavations at Klipdrift Cave will provide more information on the subsistence strategies of LSA people during the Oakhurst, both before and during the marked environmental changes of the last glacial-interglacial transition.
Acknowledgements
This research was funded in part by the European Research Council (ERC) under the European Union's Seventh Framework Programme (fP7/2007-2013)/ERC grant agreement no. 249587 awarded to C.S.H. Additional funding for this research was provided to E.D. by the University of Bergen; to C.S.H. and K.L.v.N. by the Research Council of Norway through its Centres of Excellence funding scheme, Centre for Early Sapiens Behaviour (SapienCE), project number 262618; to C.S.H. by a South African National Research Foundation Chair (SARChI) held at the Evolutionary Studies Institute, University of the Witwatersrand.
Authors' contributions
E.D.: Conceptualisation, methodology, data collection, sample analysis, data analysis, writing - the initial draft. C.S.H., K.L.v.N.: Klipdrift Cave site excavation, project leadership, project management, funding acquisition, writing - revisions.
References
1. Chase BM, Faith JT, Mackay A, Chevalier M, Carr AS, Boom A, et al. Climatic controls on Later Stone Age human adaptation in Africa's southern Cape. J Hum Evol. 2018;114:35-14. https://doi.org/10.1016/j.jhevol.2017.09.006 [ Links ]
2. Ryano KP. The Later Stone Age in the Southern Cape, South Africa, during the Terminal Pleistocene/Early Holocene with a focus on Klipdrift Cave [PhD thesis]. Johannesburg: University of the Witwatersrand; 2014. [ Links ]
3. Ryano KP Wurz S, Van Niekerk KL, Henshilwood CS. The technology of the early Oakhurst lithic techno-complex from Klipdrift Cave, Southern Cape, South Africa. Afr Archaeol Rev. 2017;34(1):93-119. https://doi.org/10.1007/s10437-017-9246-2 [ Links ]
4. Ryano KP Van Niekerk KL, Wurz S, Henshilwood CS. Shellfish exploitation during the Oakhurst at Klipdrift Cave, southern Cape, South Africa. S Afr J Sci. 2019;115(9/10), Art. #5578. https://doi.org/10.17159/sajs.2019/5578 [ Links ]
5. Henshilwood CS, Van Niekerk KL, Wurz S, Delagnes A, Armitage SJ, Rifkin RF, et al. Klipdrift shelter, southern Cape, South Africa: Preliminary report on the Howiesons Poort layers. J Archaeol Sci. 2014;45:284-303. https://doi.org/10.1016/j.jas.2014.01.033 [ Links ]
6. Brain CK. Some suggested procedures in the analysis of bone accumulations from southern African Quaternary sites. Ann Transv Mus. 1974;29(1):1-8. [ Links ]
7. Stiner MC, Kuhn SL, Weiner S, Bar-Yosef O. Differential burning, recrystallization, and fragmentation of archaeological bone. J Archaeol Sci. 1995;22:223-237. https://doi.org/10.1006/jasc.1995.0024 [ Links ]
8. Costamagno S, Théry I, Castel JC, Brugal JP Combustible ou non? Analyse multifactorielle et modèles explicatifs sur des ossements brülés paléolithiques [Fuel or not? Multifactorial analysis and explanatory models on Paleolithic burnt bones]. In: Théry I, Costamagno S, Henry A, editors. Gestion des combustibles au Paléolithique et au Mésolithique: nouveaux outils, nouvelles interpretations [Palaeolithic and mesolithic fuel management: New tools, new interpretations]. British International Series S1914. Oxford: Archaeopress; 2009. p. 65-84. Available from: http://halshs.archives-ouvertes.fr/halshs-00409188/ [ Links ]
9. Fernández-Jalvo Y Andrews P. Atlas of taphonomic identifications. Dordrecht: Springer; 2016. https://doi.org/10.1007/978-94-017-7432-1_1 [ Links ]
10. Villa P, Mahieu E. Breakage patterns of human long bones. J Hum Evol. 1991;21(1):27-48. https://doi.org/10.1016/0047-2484(91)90034-S [ Links ]
11. Grayson DK, Frey CJ. Measuring skeletal part representation in archaeological faunas. J Taphon. 2004;2(1):27-42. [ Links ]
12. Agresti A, Coull BA. Approximate is better than 'exact' for interval estimation of binomial proportions. Am Stat. 1998;52(2):119-126. https://doi.org/10.1080/00031305.1998.10480550 [ Links ]
13. Hammer 0, Harper DA, Ryan PD. PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron. 2001;4(1):9. [ Links ]
14. Discamps E, Faivre J-P. Substantial biases affecting Combe-Grenal faunal record cast doubts on previous models of Neanderthal subsistence and environmental context. J Archaeol Sci. 2017;81:128-132. https://doi.org/10.1016/j.jas.2017.03.009 [ Links ]
15. Blumenschine RJ, Madrigal TC. Variability in long bone marrow yields of East African ungulates and its zooarchaeological implications. J Archaeol Sci. 1993;20(5):555-587. https://doi.org/10.1006/jasc.1993.1034 [ Links ]
16. Roberts P Stable carbon and oxygen isotope analysis of ostrich eggshell (OES) samples from LSA and MSA levels at Blombos Cave, Klipdrift Shelter, and Klipdrift Cave (South Africa) [MSc thesis]. Oxford: University of Oxford; 2013. [ Links ]
17. Chase BM, Chevalier M, Boom A, Carr AS. The dynamic relationship between temperate and tropical circulation systems across South Africa since the last glacial maximum. Quat Sci Rev. 2017;174:54-62. https://doi.org/10.1016/j.quascirev.2017.08.011 [ Links ]
18. Klein RG. A preliminary report on the larger mammals from the Boomplaas Stone Age cave site, Cango Valley, Oudtshoorn District, South Africa. South Afr Archaeol Bull. 1978:66-75. https://doi.org/10.2307/3888252 [ Links ]
19. Faith JT. Taphonomic and paleoecological change in the large mammal sequence from Boomplaas Cave, western Cape, South Africa. J Hum Evol. 2013;65(6):715-730. https://doi.org/10.1016/j.jhevol.2013.09.001 [ Links ]
20. Schweitzer FR, Wilson ML. Byneskranskop 1: A late Quaternary living site in the southern Cape Province, South Africa. Ann S Afr Mus. 1982;88:1-203. [ Links ]
21. Klein RG. The late Quaternary mammalian fauna of Nelson Bay Cave (Cape Province, South Africa): Its implications for megafaunal extinctions and environmental and cultural change. Quat Res. 1972;2(2):135-142. https://doi.org/10.1016/0033-5894(72)90034-8 [ Links ]
22. Klein RG. Palaeoenvironmental implications of Quaternary large mammals in the fynbos region. Fynbos Palaeoecol Prelim Synth. 1983;75:116-138. [ Links ]
23. Deacon J. The Later Stone Age in the southern Cape, South Africa [PhD dissertation]. Cape Town: University of Cape Town; 1982. [ Links ]
24. Ambrose SH, Lorenz KG. Social and ecological models for the Middle Stone Age in southern Africa. In: Mellars P editor. The emergence of modern humans: An archaeological perspective. Ithaca, NY: Cornell University Press; 1990. p. 3-33. [ Links ]
25. Hogg AG, Hua Q, Blackwell PG, Niu M, Buck CE, Guilderson TP et al. SHCal13 Southern Hemisphere calibration, 0-50,000 years cal BP. Radiocarbon. 2013;55(4):1889-1903. https://doi.org/10.2458/azu_js_rc.55.16783 [ Links ]
26. Lam YM, Chen X, Pearson OM. Intertaxonomic variability in patterns of bone density and the differential representation of bovid, cervid, and equid elements in the archaeological record. Am Antiq. 1999;64(2):343-362. https://doi.org/10.2307/2694283 [ Links ]
27. Klein RG. On the taxonomic status, distribution and ecology of the blue antelope, Hippotragus leucophaeus (Pallas, 1766). Ann S Afr Mus. 1974;65(4):99-143. [ Links ]
28. Faith JT. Conservation implications of fossil roan antelope (Hippotragus equinus) in southern Africa's Cape Floristic Region. In: Louys J, editor. Paleontology in ecology and conservation. Berlin: Springer; 2012. p. 239-251. https://doi.org/10.1007/978-3-642-25038-5_12 [ Links ]
 Correspondence:
Correspondence:
Emmanuel Discamps
ediscamps@gmail.com
Received: 14 Aug. 2019
Revised: 31 Jan. 2020
Accepted: 03 Feb. 2020
Published: 27 May 2020
Editor: Maryna Steyn
Funding: European Research Council (grant no. 249587); University of Bergen; Research Council of Norway (SapienCE project number 262618); National Research Foundation (South Africa)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}