










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.115 no.9-10 Pretoria sep./oct. 2019
http://dx.doi.org/10.17159/sajs.2019/6404
RESEARCH LETTER
Endostructural assessment of a hominin maxillary molar (StW 669) from Milner Hall, Sterkfontein, South Africa
Bontle MatabogeI; Amélie BeaudetII, III; Jason L. HeatonI, IV; Travis R. PickeringV; Dominic StratfordII
IEvolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa
IISchool of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, Johannesburg, South Africa
IIIDepartment of Anatomy, University of Pretoria, Pretoria, South Africa
IVDepartment of Biology, Birmingham-Southern College, Birmingham, Alabama, USA
VDepartment of Anthropology, University of Wisconsin, Madison, Wisconsin, USA
ABSTRACT
The site of the Sterkfontein Caves, South Africa, is one of the richest early hominin fossil-bearing sites in Africa. Recent excavations in the Milner Hall locality have contributed to the discovery of new hominin specimens, including StW 669, a right permanent maxillary first molar (M1). StW 669 was excavated from the T1 deposits, which consist of a mixture of sediments from Members 2 and 5 of the Sterkfontein Formation. Accordingly, the deposits have the potential to contain remains of Australopithecus, Paranthropus and Homo. In this study, we employed micro-focus X-ray tomography in order to assess dental tissue proportions, enamel thickness distribution and enamel-dentine junction morphology as approaches to investigate the taxonomy of StW 669. We compare our results to those generated on the teeth of Australopithecus africanus, Paranthropus robustus, Homo erectus, Homo antecessor, Homo neanderthalensis and Homo sapiens. Our results suggest that StW 669 shares quantitative and qualitative affinities with M1s of Homo in terms of tissue proportions (i.e. two- and three-dimensional average and relative enamel thickness of 1.2-1.3 mm and 18.4, respectively) and enamel thickness distribution (i.e. thickest enamel on the lingual aspect of the protocone). However, data on the enamel-dentine junction morphology of StW 669 are inconclusive as to the tooth's taxonomic affinities. Pending additional morphometric analyses, our studies of inner morphology of the crown of StW 669 support its attribution to Homo.
SIGNIFICANCE:
•The Sterkfontein Caves have contributed significantly to our understanding of early human evolution and continue to do so. This study highlights the specific value of the Milner Hall locality as a valuable store of hominin fossils. Moreover, we tentatively clarify the enigmatic taxonomic status of StW 669, a right permanent maxillary first molar, excavated from the T1 deposit of Milner Hall. Pending additional morphometric evidence, our preliminary data on tissue proportions, enamel thickness distribution and enamel-dentine junction morphology, suggest an attribution of StW 669 to early Homo. This result is significant given the historical contention concerning the presence of early Homo at Sterkfontein
Keywords: Homo, late Pliocene-early Pleistocene, maxillary first molar, enamel-dentine junction, EDJ, enamel thickness
Introduction
The Milner Hall is a deep underground chamber within the Sterkfontein Caves (South Africa) that extends about 100 m in an east-west direction.1 The complex stratigraphic context of the Milner Hall fossiliferous depositional sequence - in which an early distal accumulation of the 3.67-Ma-old Member 2 (T3) and 2.18-Ma Oldowan artefact-bearing sediments from Member 5 (T2) contribute to the formation of the uppermost fossil- and artefact-bearing deposit, T11-3 - affords the potential for fossils of Australopithecus, Paranthropus and early Homo to be represented in the unit. Four hominin fossils - a manual proximal phalanx (StW 668), a right permanent maxillary first molar (M1) (StW 669) and two cranial vault fragments (StW 671 and StW 672) from the T1 depositional unit of the Central Underground Deposits excavation site of Milner Hall (STK-MH1) - have been described previously.2,4
The description and metrical analyses of the hominin remains excavated from T1 revealed an enigmatic mix of unique, primitive and derived morphological traits, some of which indicated affinities with the genus Homo.2,4 More specifically, the dimensions and shape of the crown of StW 669 were found to be more consistent with those of Homo than of Australopithecus and Paranthropus, and most similar to the Olduvai Homo habilis specimen OH 6 (Tanzania) and to the Homo naledi specimen UW 101-1688 from Rising Star (South Africa).2 The attribution of StW 669 to Homo sp. is of particular interest for assessing the presence of early Homo at Sterkfontein and improving our understanding of the early hominin taxonomic diversity in South Africa.5,6
In association with significant improvements in analytical tools, the development of advanced imaging techniques, more specifically micro-focus X-ray tomography, offers the opportunity to assess taxonomically relevant endostructural information non-invasively.7 Variation in tissue proportions, enamel thickness distribution and morphology of the enamel-dentine junction (EDJ) recorded in fossil hominins has been particularly useful for discriminating hominin taxa8-10 and for clarifying long-standing uncertainties on the assignment of isolated fossil teeth11.
In this context, we provide here additional evidence from tooth crown endostructural analysis in order to clarify the taxonomic status of StW 669 and to tentatively contribute to ongoing debates about hominin morphological and taxonomic diversity at Sterkfontein during the late Pliocene and early Pleistocene.
Material and methods
StW 669 is a rootless right M1 described by Stratford et al.2 The occlusal surface of StW 669, which has a square outline (i.e. mesiodistal diameter:buccolingual diameter ratio of 1.02), is relatively unworn but the enamel formation has been disrupted at the tip of the paracone, at the buccal occlusal margin of the paracone, and on the mesial face of the tooth.2 The occlusal area is small and relatively symmetric, and the areas of the paracone, hypocone and metacone are smaller than the areas of the protocone, hypocone and metacone.2 The paracone is particularly small and the metacone area:paracone area ratio is high (i.e. 1.26).2
To discuss the taxonomic attribution of StW 669, qualitative and quantitative results derived from the endostructural three-dimensional (3D) analysis of StW 669 are compared to published data on tissue proportions, enamel distribution and enamel-dentine junction morphology of the permanent M1 crowns of Australopithecus africanus9,12, Paranthropus robustus9, Homo erectus13, Homo antecessor14, Homo neanderthalensis11,14,15, European Middle Pleistocene Homo16 and modern Homo sapiens11,14,17-19. Unfortunately, to the best of our knowledge, there is no information available on the endostructure of the first molar of Homo naledi. As only a few specimens could be directly compared to StW 669 in terms of 3D tissue proportions (see Table 1), we also performed additional two-dimensional (2D) measurements on 2D sections that are analogous to previous studies (Table 2).
StW 669 was scanned with a Nikon XTH 225/320 LC dual-source industrial CT scanner at the microfocus X-ray tomography facility of the Evolutionary Studies Institute at the University of the Witwatersrand (Johannesburg, South Africa) using the following parameters: 70 kV, 100 µA, 3142 projections, an acquisition time of two frames per second and an isotropic voxel size of 26.7 µm (Figure 1). The dental tissues were obtained through a semi-automatic threshold-based segmentation and 3D mesh reconstructions via the software Avizo v.9.0 (Visualization Sciences Group Inc., Hillsboro, OR, USA) and verified by two users (B.M. and A.B.; Figure 1). The dental crown was digitally closed following the protocol detailed in Beaudet et al.20 This coronal volume embeds both the dentine material and pulp cavity. In addition to the 3D volume, we extracted a virtual section of StW 669 passing through the mesial dentine horns to assess the tissue proportions in 2D following previously published protocols.9,14,21
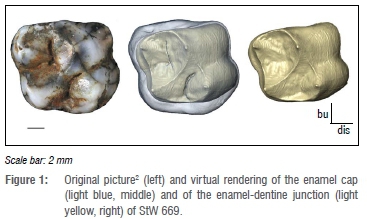
Three variables were measured in 3D and 2D8,9,21: volume (Ve, mm3) and area (c, mm2) of the enamel cap; volume (Vcdp, mm3) and area (b, mm2) of the coronal dentine that includes the coronal aspect of the pulp chamber; and surface (SEDJ, mm2) and length (e, mm) of the enamel-dentine junction. We then calculated two indices of enamel thickness: 3D (3D AET, mm) and 2D (2D AET, mm) average enamel thickness, calculated as the ratio between Ve/c and SEDJ/e; 3D (3D RET) and 2D (2D RET) relative enamel thickness, obtained through the ratios 3D AET/(Vcdp)1/3*100 and 2D AET/b1/2*100 and allowing direct, scale-free comparisons.8-9,21 Intra- and inter-observer tests for measurement accuracy run by two observers in previous published studies using a similar analytical protocol revealed differences less than 5%.22
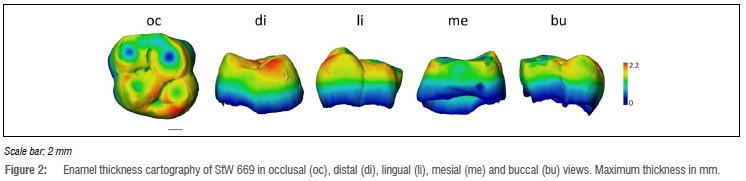
We assessed the 3D distribution of enamel across the crown by computing the distances between the occlusal and the enamel-dentine junction surfaces through the 'Surface Distance' module on Avizo v.9.0.20 The distances recorded were visualised at the outer enamel surface using a colour scale ranging from dark blue ('thinner') to red ('thicker').
Results
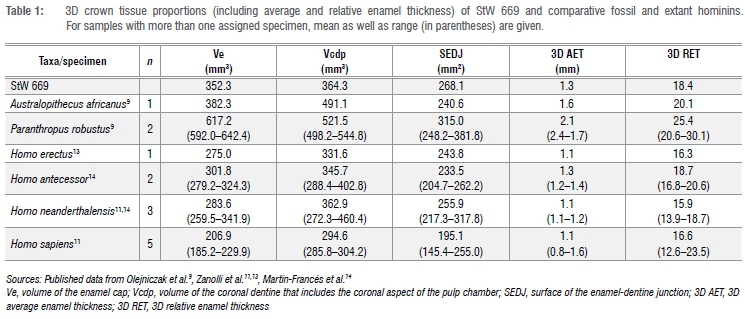
Tissue proportions
3D and 2D tissue proportions of StW 669 and comparative specimens are shown in Table 1 and Table 2. The volume of enamel (Ve) in StW 669 is higher than in H. erectus, H. antecessor, H. neanderthalensis and modern humans, but lower than in P. robustus, and fits more closely the value for A. africanus. Conversely, coronal dentine and pulp volume (Vcdp) in StW 669 is higher than in modern humans but lower than in A. africanus and P. robustus and approximates more closely the figures of H. erectus, H. antecessor and H. neanderthalensis. The surface of the enamel-dentine junction is intermediate between A. africanus, the Homo species and P. robustus. In general, the 2D enamel area (c), coronal dentine and pulp area (a) as well as the EDJ length (e) in StW 669 are lower than in A. africanus, P. robustus and H. erectus, but fits the range of, or at least are close to, the values reported for H. antecessor, H. neanderthalensis, European Middle Pleistocene Homo and H. sapiens.
In terms of average (3D AET) and relative (3D RET) enamel thickness, values recorded for StW 669 are lower than in the single representative of A. africanus and the two specimens of P. robustus and higher than in H. erectus and H. neanderthalensis, but are close to the values of H. antecessor (Table 1). Both 3D AET and 3D RET fall within the range of modern humans. The 2D AET fits the range of fossil and modern humans but is lower than in A. africanus and P. robustus. The 2D RET is consistent with the H. antecessor and H. sapiens ranges of variation, but is higher than in H. neanderthalensis and lower than in A. africanus, P. robustus and H. erectus.
Enamel distribution
The cartography that maps topographic enamel thickness variations at the outer crown surface of StW 669 is shown in Figure 2. Isolated spots correspond to enamel removal following wear (even if moderate here, i.e. there is no exposed dentine)2 or disruptions in the enamel formation (i.e. deep, irregularly shaped pits)2. The thickest enamel in StW 669 is found along the oblique ridge connecting the metacone and the protocone, on the lingual aspect of the protocone and on the distal aspect of the hypocone. In these respects, the enamel distribution of StW 669 fits the fossil and extant human condition by sharing thick enamel on the lingual aspect of the crown13,14,17 but differs from Australopithecus and Paranthropus that have thicker enamel at the cusp tips9.
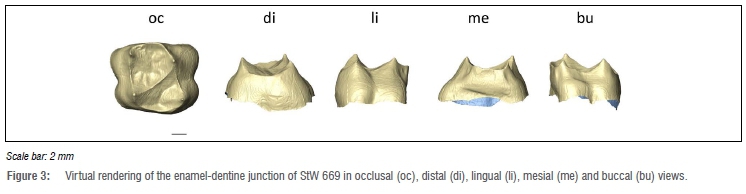
Enamel-dentine junction shape
The StW 669 EDJ shape is shown in Figure 3. The EDJ reflects the uncomplicated topography described for the occlusal surface.2 The EDJ shows an oval-rectangular outline. An uninterrupted oblique ridge links the metacone to the protocone and separates central and distal fossae, the former being larger than the latter. The mesial marginal ridge is sharp and particularly high compared to the lateral and distal ones. The dentine horns are equally elevated. The protocone is more distally set compared to the paracone while the hypocone is more lingually set compared to the protocone. Carabelli's trait expression corresponds to grade 1 (i.e. a groove is present).23 The StW 669 EDJ morphology is thus comparable to previous descriptions of Australopithecus12 and Homo13-15 M1s.
Discussion
With hominin remains dated to either 3.67 Ma or 2.18 Ma, the Milner Hall fossil assemblage has the potential to provide further evidence useful for assessing early hominin palaeobiodiversity and the taxonomic context of the Sterkfontein hominin-bearing deposits. Consistent with previous descriptions and measurements of the outer morphology,2 our micro-tomographic-assisted revision of StW 669 and characterisation of the endostructure reveal additional Homo affinities of the tooth. The identification of fossil Homo remains is critical in the context of Sterkfontein (and of South Africa as a whole) given the taxonomic ambiguity of specimens previously assigned to early Homo (e.g. StW 53)24 and the fragmentary nature of fossil specimens identified as Homo (e.g. SK 847)25. Moreover, the diagnosis of the hominin remains found in the stone tools bearing deposits of T1 of Milner Hall may have implications for discussing the identity of the toolmakers at Sterkfontein.
Unfortunately, our data are not sufficient to determine with certainty to which species of Homo StW 669 should be assigned. The lack of consensus on the diversity of early humans, particularly in South Africa6, represents another limiting factor. However, given the success of previous studies in identifying human fossil remains in South Africa by examining the molar crown inner structural organisation26, future quantitative analysis of the enamel-dentine junction shape using morphometric methods10-13 will be useful for refining the taxonomic status of StW 669.
Acknowledgements
We are indebted to B. Zipfel (Johannesburg) for granting access to the fossil specimen. We thank K. Jakata (Johannesburg) for microtomographic acquisitions. For technical and/or scientific discussion/collaboration we are grateful to R. Clarke (Johannesburg). We also thank the two anonymous reviewers for their comments that significantly improved this contribution and Associate Editor Maryna Steyn for managing the publication of this article. We thank the DST-NRF for sponsoring the Micro-XCT facility at Necsa, and the DST-NRF and Wits University for funding the microfocus X-ray CT facility in the Evolutionary Studies Institute (www.wits.ac.za/microct). The support of the DST-NRF Centre of Excellence in Palaeosciences (CoE-Pal) and the Palaeontological Scientific Trust (PAST) towards this research is hereby acknowledged. Opinions expressed and conclusions arrived at, are those of the authors and are not necessarily to be attributed to the Centre of Excellence in Palaeosciences.
Authors' contributions
B.M.: Data collection; sample and data analysis; validation; writing. A.B.: Conceptualisation; methodology; sample and data analysis; validation; writing; student supervision; project leadership. J.L.H.: Writing. T.R.P.: Writing. D.S.: Conceptualisation; writing; student supervision; project management; funding acquisition.
References
1.Stratford DJ, Grab S, Pickering TR. The stratigraphy and formation history of fossil-and artefact-bearing sediments in the Milner Hall, Sterkfontein Cave, South Africa: New interpretations and implications for palaeoanthropology and archaeology. J Afr Earth Sci. 2014;96:155-167. https://doi.org/10.1016/j.jafrearsci.2014.04.002 [ Links ]
2.Stratford D, Heaton JL, Pickering TR, Caruana MV, Shadrach K. First hominin fossils from Milner Hall, Sterkfontein, South Africa. J Hum Evol. 2016;91:167-173. https://doi.org/10.1016/j.jhevol.2015.12.005 [ Links ]
3.Granger DE, Gibbon RJ, Kuman K, Clarke RJ, Bruxelles L, Caffee MW. New cosmogenic burial ages for Sterkfontein Member 2 Australopithecus and Member 5 Oldowan. Nature. 2015;522:85-88. https://doi.org/10.1038/nature14268 [ Links ]
4.Beaudet A, Heaton JL, L'Abbé EN, Pickering TR, Stratford D. Hominin cranial fragments from Milner Hall, Sterkfontein, South Africa. S Afr J Sci. 2018;114(11/12), Art. #5262, 6 pages. https://doi.org/10.17159/sajs.2018/5262 [ Links ]
5.Clarke RJ. Homo habilis: The inside story. In: Sahnouni M, Semaw S, Garaizar JR, editors. Proceedings of the II Meeting of African Prehistory. Burgos: Consorcio CENIEH; 2017. p. 25-51. [ Links ]
6.Grine FE, Smith HF, Heesey CP, Smith EJ. Phenetic affinities of Plio-Pleistocene Homo fossils from South Africa: Molar cusp proportions. In: Grine FE, Fleagle JG, Leakey RE, editors. The first humans: Origin and early evolution of the genus Homo. Dordrecht: Springer; 2009. p. 49-62. https://doi.org/10.1007/978-1-4020-9980-9 [ Links ]
7.Macchiarelli R, Bayle P, Bondioli L, Mazurier A, Zanolli C. From outer to inner structural morphology in dental anthropology. The integration of the third dimension in the visualization and quantitative analysis of fossil remains. In: Scott GR, Irish JD, editors. Anthropological perspectives on tooth morphology, genetics, evolution, variation. Cambridge: Cambridge University Press; 2013. p. 250-277. https://doi.org/10.1017/CBO9780511984464.011 [ Links ]
8.Olejniczak AJ, Smith TM, Feeney RNM, Macchiarelli R, Mazurier A, Bondioli L, et al. Dental tissue proportions and enamel thickness in Neandertal and modern human molars. J Hum Evol. 2008;55:12-23. https://doi.org/10.1016/j.jhevol.2007.11.004 [ Links ]
9.Olejniczak AJ, Smith TM, Skinner MM, Grine FE, Feeney RNM, Thackeray JF, et al. Three-dimensional molar enamel distribution and thickness in Australopithecus and Paranthropus. Biol Lett. 2008;4:406-410. https://doi.org/10.1098/rsbl.2008.0223 [ Links ]
10.Skinner MM, Gunz P, Wood BA, Hublin J-J. Enamel-dentine junction (EDJ) morphology distinguishes the lower molars of Australopithecus africanus and Paranthropus robustus. J Hum Evol. 2008;55:979-988. [ Links ]
11.Zanolli C, Kullmer O, Kelley J, Bacon AM, Demeter F, Dumoncel J, et al. Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia. Nat Ecol Evol. 2019(3):755-764. https://doi.org/10.1038/s41559-019-0860-z [ Links ]
12.Braga J, Thackeray JF, Subsol G, Kahn JL, Maret D, Treil J, et al. The enamel-dentine junction in the postcanine dentition of Australopithecus africanus: Intra-individual metameric and antimeric variation. J Anat. 2010;216:62-79. https://doi.org/10.1111/j.1469-7580.2009.01154 [ Links ]
13.Zanolli C. Molar crown inner structural organization in Javanese Homo erectus. Am J Phys Anthropol. 2015;156:148-157. https://doi.org/10.1002/ajpa.22611 [ Links ]
14.Martín-Francés L, Martinón-Torres M, Martínez de Pinillos M, García-Campos C, Modesto-Mata M, Zanolli C, et al. Tooth crown tissue proportions and enamel thickness in Early Pleistocene Homo antecessor molars (Atapuerca, Spain). 2018;13(10), e0203334, 29 pages. https://doi.org/10.1371/journal.pone.0203334 [ Links ]
15.Martin RMG, Hublin JJ, Gunz P, Skinner MM. The morphology of the enamel-dentine junction in Neanderthal molars: Gross morphology, non-metric traits, and temporal trends. J Hum Evol. 2017;103:20-44. https://doi.org/10.1016/j.jhevol.2016.12.004 [ Links ]
16.Smith TM, Olejniczak AJ, Zermeno JP, Tafforeau P, Skinner MM, Hoffmann A, et al. Variation in enamel thickness within the genus Homo. J Hum Evol. 2012;62:395-411. https://doi.org/10.1016/j.jhevol.2011.12.004 [ Links ]
17.Kono RT, Suwa G, Tanijiri T. A three-dimensional analysis of enamel distribution patterns in human permanent first molars. Arch Oral Biol. 2002;47:867-875. [ Links ]
18.Kono RT. Molar enamel thickness and distribution patterns in extant great apes and humans: new insights based on a 3-dimensional whole crown perspective. Anthropol Sci. 2004;112:121-146. [ Links ]
19.Smith TM, Olejniczak AJ, Reid DJ, Ferrell RJ, Hublin J-J. Modern human molar enamel thickness and enamel-dentine junction shape. Arch Oral Biol 2006;51:974-95. https://doi.org/10.1016/j.archoralbio.2006.04.012 [ Links ]
20.Beaudet A, Dumoncel J, Thackeray JF, Bruxelles L, Duployer B, Tenailleau C, et al. Upper third molar internal structural organization and semicircular canal morphology in Plio-Pleistocene South African cercopithecoids. J Hum Evol. 2016;95:104-120. https://doi.org/10.1016/j.jhevol.2016.04.004 [ Links ]
21.Martin L. Significance of enamel thickness in hominoid evolution. Nature. 1985;314(6008):260-263. https://doi.org/10.1038/314260a0 [ Links ]
22.Bondioli L, Bayle P, Dean C, Mazurier A, Puymerail L, Ruff C, et al. Morphometric maps of long bone shafts and dental roots for imaging topographic thickness variation. Am J Phys Anthropol. 2010;142:328-334. [ Links ]
23.Ortiz A, Skinner MM, Bailey SE, Hublin JJ. Carabelli's trait revisited: An examination of mesiolingual features at the enamel-dentine junction and enamel surface of Pan and Homo sapiens upper molars. J Hum Evol. 2012;63:586-596. https://doi.org/10.1016/j.jhevol.2012.06.003 [ Links ]
24.Grine FE. The alpha taxonomy of Australopithecus africanus. In: Reed KE, Fleagle JG, Leakey RE, editors. The paleobiology of Australopithecus. Dordrecht: Springer; 2013. p. 74-104. https://doi.org/10.1007/978-94-007-5919-0 [ Links ]
25.Clarke RJ. The cranium of the Swartkrans hominid, SK 847 and its relevance to human origins [PhD dissertation]. Johannesburg: University of the Witwatersrand; 1977. [ Links ]
26.Zanolli C, Pan L, Skinner MM, Dumoncel J, Beaudet A, De Beer F, et al. What is South African early Homo? New insights from the molar endostructural signature. Paper presented at: Meeting of the European Society for the Study of Human Evolution; 2018 September 13-15; Faro, Portugal. [ Links ]
 Correspondence:
Correspondence:
Amélie Beaudet
beaudet.amelie@gmail.com
Received: 27 May 2019
Revised: 26 July 2019
Accepted: 12 Aug. 2019
Published: 26 Sep. 2019
EDITOR: Maryna Steyn
FUNDING: Palaeontological Scientific Trust; DST-NRF Centre of Excellence in Palaeosciences
*Data are available upon application from Dr Bernhard Zipfel, Curator of Fossil and Rock Collections at the University of the Witwatersrand (Bernhard.Zipfel@wits.ac.za)


{kind=link}
{kind=link}
{kind=link}
{kind=link}