










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.115 no.7-8 Pretoria Jul./Ago. 2019
http://dx.doi.org/10.17159/sajs.2019/5496
RESEARCH ARTICLES
An analysis of the effect of tooth wear on bovid identification
Juliet K. BrophyI; Gregory J. MatthewsII; George K. ThiruvathukalIII
IDepartment of Geography and Anthropology, Louisiana State University, Baton Rouge, Louisiana, USA
IIDepartment of Mathematics and Statistics, Loyola University Chicago, Chicago, Illinois, USA
IIIDepartment of Computer Science, Loyola University Chicago, Chicago, Illinois, USA
ABSTRACT
Previous research provides a method for reducing the subjectivity in taxonomic identification of species in the family Bovidae by quantifying the occlusal surface of molar teeth using elliptical Fourier analysis. In this current study, we specifically test what effect medium to late tooth wear has on the identification of bovids when using the form (size and shape) of the occlusal surface to classify specimens. To achieve this, the classification results of teeth with ≥85% of their occlusal surface (training data set) were compared with the results of teeth with <85% of their occlusal surface (test data set) due to wear. In the training data set, all tribes classified correctly >87% of the time with both Alcelaphini and Reduncini classifying correctly >90% of the time. The worn teeth in the test data set classified correctly at lower rates, but all tribes still collectively had a good classification accuracy (>60%) with classification of Alcelaphini and Tragelaphini at >75% and Reduncini at >65%. Hippotragini classified correctly 50% of the time and Neotragini 42%. The one worn Bovini tooth in the test data set was classified as that of a Hippotragini. The classification rates of teeth with medium to late wear were lower than the unworn teeth, but not so low as to suggest that this methodology for identifying bovid teeth is inappropriate for worn teeth, especially when supplementary to other methods.
SIGNIFICANCE:
•Worn teeth can be taxonomically identified by performing elliptical Fourier analysis on the occlusal surface of their teeth.
•While teeth with medium to late wear correctly classify at lower rates than relatively unworn ones, the methodology is a valuable supplement to other methods for taxonomically identifying bovid teeth.
•Worn teeth of the tribes Alcelaphini and Tragelaphini classify at the highest rates.
Keywords: Bovidae; worn teeth; elliptical Fourier analysis; machine learning
Introduction
Fossil remains from the family Bovidae, such as antelopes and buffalo, are used frequently to reconstruct past environments.1-4 Bovid teeth, in particular isolated bovid teeth, are some of the most common fossils found in the southern African record. However, taxonomic identification of bovid teeth is often difficult because of overlap in their shape and size. Biasing factors such as sex, age-at-death and degree of occlusal (chewing surface) attrition often result in considerable overlap in absolute and relative tooth size, thereby complicating taxonomic identification.4 Thus, taxonomic diagnoses of some bovid teeth often do not extend beyond broad levels such as the tribe or family.3,5-10 Furthermore, the overlap in tooth shape and/or size often leads to interobserver error as researchers cannot reliably compare the faunal lists produced by different analysts. This result is often because of differences in experience, confidence levels in identifications and access to comparative materials. Researchers can, therefore, analyse the same assemblages and report different results. For example, Vrba6, Wells and Cooke11, and Reed12 reported varying identifications and subsequently concluded different palaeoenvironmental reconstructions for Makapansgat, South Africa.
Previous research has provided a method for reducing the subjectivity in bovid identification by quantifying the occlusal surface of bovid teeth and suggests that teeth from closely related species can be reliably distinguished from each other based on an analysis of the form (shape and size) of their occlusal outline.4,13-15 These previous studies involved the digitisation and extraction of the outlines of the occlusal surface of molars (upper three and lower three) from 20 species. Elliptical Fourier analysis (EFA) was used to approximate each shape as a series of ellipses. The coefficients generated by EFA were then used as features in machine learning algorithms to classify specimens into taxonomic groups. By applying machine learning techniques to the amplitudes generated by EFA for modern bovids, >80% of specimens in each of the six teeth groups classified correctly to tribe.14 The ultimate purpose of those studies was to develop a methodology to identify fossil representatives of modern counterparts in the fossil record of southern Africa with greater accuracy. These results can then be utilised as a supplement to other defining characteristics of bovid teeth, such as degree of hypsodonty, enamel thickness and rugosity.
Importantly, the teeth used in the aforementioned studies were not separated into different wear stages, although the samples did include worn teeth. The purpose of these studies was solely to produce a reliable, replicable and practical tool for classifying modern and fossil bovid teeth. The goal was thus to avoid separating the teeth in the reference data set into wear stages in order to encompass more natural, biological variation within the data set and to make the program more accessible to a wider range of teeth. While the teeth were not separated into wear stages, the authors of each paper did state that at least 85% of the occlusal surface of the tooth, as determined by someone familiar with bovid teeth, was required to enable use of this method (Figure 1).
Brophy13 and Brophy et al.4 specifically investigated whether the average shape of the occlusal surface of a tooth remained consistent over the lifespan of a bovid, regardless of age and attrition. Their research assessed intra-tooth variation by means of computed tomography (CT) using teeth from different bovid tribes and applying EFA to specific increments of CT scan slices from the same tooth. The Fourier-derived amplitudes were considered 'unknowns' and compared with those of known bovid teeth, using linear discriminant analyses. The 'unknown' digitised CT scans were correctly classified to the known species ≥85% of the time.4
The teeth utilised in Brophy13 and Brophy et al.4 were not separated into wear stages but were digitised by one of the authors who is familiar with the shape of bovid teeth. The teeth employed in Matthews et al.15 were also not separated into wear stages but were digitised as Human Intelligence Tasks by Amazon Mechanical Turk workers, or Turkers. These individuals, very likely, had no a priori knowledge of bovid teeth other than the video tutorial given to them in order to complete the task. In Matthews et al.15, the same tooth was digitised three times by Turkers. One author (J.K.B.) reviewed every Turker submission before using it in the project to ensure that the digitised tooth did not include information other than the actual tooth. Teeth were removed from the study if they included information other than the outline of the tooth, as explained by the Human Intelligence Task. Using a random forest algorithm, the classification results based on the teeth digitised by J.K.B. were compared with the Turkers' results. This process was done using the average shape and generalised Procrustes analysis of the three replications of digitisations created by the Turkers. While classification of the J.K.B. outlines outperformed the Turker outlines, the differences were relatively small and in both cases each tooth was classified correctly to tribe ≥80% of the time in leave-one-out cross-validation tests.15 While the purpose of that study was to determine if the digitisation process could be crowd sourced, the high correct classification results provide support for not separating the teeth into wear stages.
The results of the aforementioned studies indicate that the shape of the tooth throughout an animal's life does not change significantly enough to impede identification of that tooth using the occlusal surface. The findings also support the use of occlusal outlines as reliable indicators of bovid tribe and species. It is important to note that while the overall classification rate is high, in all of these studies some tooth types (e.g. M2) and taxonomic groups classified with higher accuracy than others. In addition, while worn teeth were included in these studies, heavily worn teeth were not targeted for the sample.
This current study expands on the previous results and specifically assesses the effect medium to late wear (<85% of their occlusal surface) has on the identification of bovid teeth when using the form of the occlusal surface and EFA to classify specimens (see Figure 1 for an example). For the purpose of this paper, medium to late wear includes teeth with (1) enamel rim ranges from thin to missing on at least one side, (2) infundibulum reduced or worn away completely, (3) a small remnant of or missing goat fold and (4) a crown height spanning from severely to completely lost. These categories were adapted from Klein's dental occlusal wear stages for bovid molars.16
Essentially, the study was designed to investigate whether a completely unworn or slightly worn (i.e. fully observed) tooth is absolutely necessary to make an accurate identification; in other words, can a tooth classify correctly when it is significantly worn? To achieve this, the classification results of teeth with ≥85% of their occlusal surface (as determined by J.K.B.) were compared with the results of teeth with <85% of their occlusal surface intact, as a result of wear (Figure 1). The expectation was that the classification rates of the worn teeth would be lower than those of the fully observed ones, but not so low as to suggest that this methodology for identifying bovid teeth is not applicable to worn teeth.
Materials and methods
Bovid tooth images were obtained from the University of the Witwatersrand (UW), Ditsong National Museum of Natural History (TM and AZ), and National Museum Bloemfontein (NMB). The teeth were systematically oriented in the same manner with the cervical plane perpendicular to a digital camera so that the occlusal surface could be seen clearly. A levelled scale bar was placed directly next to the tooth at the height of the occlusal surface. Photographs were taken of the three upper and three lower molars, which included 20 species from seven tribes (Table 1). The outlines of the occlusal surfaces for the training data set (≥85% of their occlusal surface intact) were obtained using MLmetrics (see Brophy et al.4 for protocol). Briefly, this process involved creating outlines of the same tooth type (e.g. M2) from a given species by manually placing 60 points around the enamel edge according to a template. MLmetrics generated x-y coordinates for each of the 60 points that were used in the training data set.
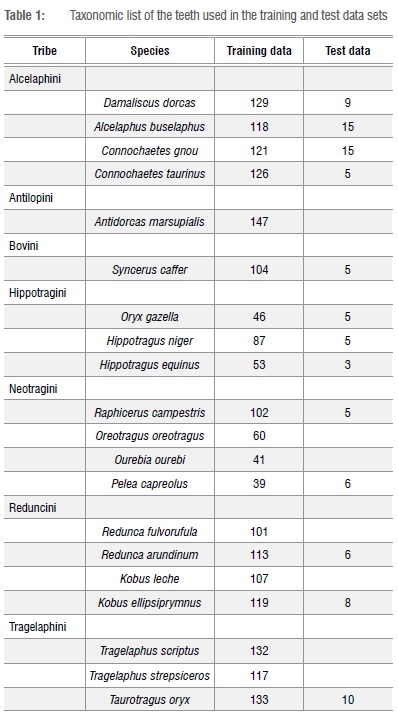
A sample of 92 teeth with medium to late wear were chosen for the test sample of this study from six different tribes and 13 species (Table 1). It is important to note that the majority of the test data set was from Alcelaphini species and that the distribution across tooth types is highly heterogeneous; some tribes number more individuals than others. Alcelaphini is typically one of the most common tribes at sites in southern Africa e.g. Elandsfontein17, Equus Cave18, Gladysvale19, Sterkfontein20 and Plovers Lake3; therefore this tribe was targeted in this study.
Each tooth in the test data set for this study was digitised using the freeware GIMP (the GNU Image Manipulation Program; www.gimp.org) according to the same method outline in Matthews et al.15, briefly described hereafter. The image of a bovid tooth was downloaded in GIMP and an outline of the tooth was created using the lasso function. Next, the outline was converted to a black and white image. The black and white images were then imported into R using the Momocs package.21 This function extracted x- and y-coordinates along the edge between the black and white fields in the images. A model was then trained using a data set containing only teeth with ≥85% of their occlusal surface intact, to assess how accurately the worn teeth could be classified.
The shape of each tooth in both the training and test data sets was approximated using EFA with 20 harmonics. Each harmonic generates four components; 80 coefficients in total were used as features in a random forest classification model with 5000 trees to predict tribe membership (see Matthews et al.14). Additionally, 20 principal components based on the 80 coefficients were used as possible features in the model. Models were fit separately for each tooth type (M1, M2, etc.). The fitted random forest model was used to predict tribe membership for each specimen in the test data set. A confusion matrix was generated which displays which teeth classified correctly. Tables 2 and 3, respectively, reveal the results of the classification performance of the training data and test data set.
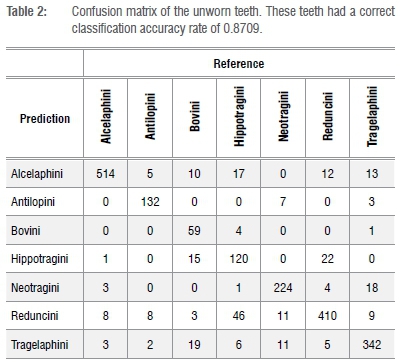
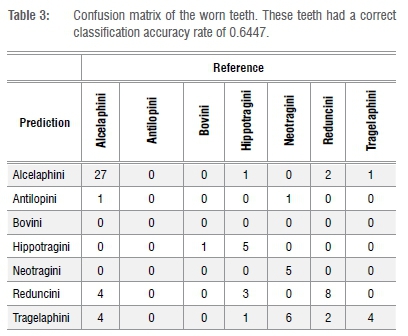
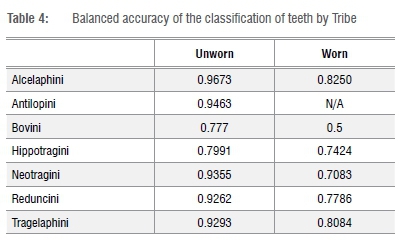
Table 4 shows the balanced accuracy of the unworn and worn teeth. Balanced accuracy is an average of sensitivity and specificity. Sensitivity is the true positive rate and explains, for example, if a tooth predicted to be an Alcelaphini and was actually an Alcelaphini. Specificity is a true negative rate, which, for example, shows when a tooth that is not an Alcelaphini did not predict to be an Alcelaphini.
Results
In the training data set, all tribes classified correctly more than 87% of the time, with both Alcelaphini and Reduncini classifying over 90% correctly (Tables 2 and 4). As expected, lower rates of classification accuracy were found for the worn teeth, but all of the tribes combined had a classification accuracy of more than 60%, with Alcelaphini (n=36) and Tragelaphini (n=5) both at more than 75% and Reduncini (n=12) at over 65% (Tables 3 and 4). Hippotragini (n=10) classified with a 50% accuracy while the tribe Neotragini (n=12) classified with only 42% accuracy. The one Bovini in the test data set classified as a Hippotragini. While these classification rates are certainly lower than those for the less worn teeth, they are still high enough to prove useful for taxonomic classification.
Discussion
The results of this study suggest that using EFA is a viable, supplemental method for classifying worn bovid teeth, even when there is significant wear. This study highlights how wear can often complicate taxonomic identification. Molars such as the maxillary and mandibular first molars (M1, M1) tend to be more difficult to identify because they erupt first and so generally have more wear than other molars. The shape and size of teeth overlap more for some tribes than for others, making identification of these tribes particularly challenging; this overlap is even more exaggerated when the teeth are heavily worn. Any methodology that can assist in narrowing the classification of the taxon of a worn tooth is useful, but the percentages reported here are particularly informative.
In this study, Alcelaphini, Tragelaphini and Reduncini were identified with the highest rates of the six tribes. These results suggest that the form of their teeth do not change significantly throughout their lifetime, regardless of wear, making this methodology particularly useful for these three tribes.
The Hippotragini, Neotragini and Bovini were not identified with as high classification rates, thereby suggesting that their teeth (specifically those teeth with medium to late wear) are more difficult to classify. Half of the Hippotragini teeth were too worn to classify correctly. As a result of attrition, these teeth lost some of the classic features of Hippotragini, resulting in the tooth appearing more similar to those of Reduncini or Alcelaphini. Neotragini teeth overlap in size and, with attrition, look more similar to those of Tragelaphus scriptus (bushbuck). Therefore, six of the Neotragini teeth were classified as belonging to Tragelaphini. Finally, although the one Bovini tooth was classified as that of a Hippotragini, there was only one tooth. A larger sample of worn Bovini teeth is needed to demonstrate whether this result is typical of all Bovini teeth. The EFA methodology can aid in identifying teeth from these tribes, but teeth with medium to heavy wear may require additional methods to ratify the identification. Such methodology may involve degree of hypsodonty, enamel thickness, rugosity, and/or associated teeth/horn cores, depending on what is present. Other specimens, in particular isolated teeth, may prove to present too little data to enable classification.
The following four examples provide more background related to the results of this study, that is, reasons for the correct or incorrect classification of teeth in the test data set. All the teeth in these cases exhibited heavy wear and, according to van Zyl22, would have been at least 37 months old.
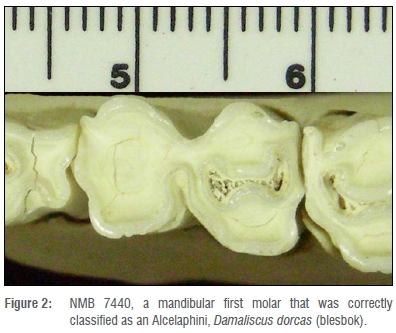
The first case is tooth NMB 7440 - a worn lower, left, first molar that was correctly classified as that of an Alcelaphini, Damaliscus dorcas (blesbok) (Figure 2). The mesial lobe has been obliterated by wear with no infundibulum, while the distal lobe has retained the infundibulum. The tooth is distorted mesially and distally as a result of interproximal wear from both the fourth premolar and the second molar. The mesial side of the tooth angles mesiobuccally to distolingually. The enamel around the buccal portion of the mesial lobe (protoconid) is also damaged. Distally, the tooth curves from the eruption of the second molar. Regardless, the program was able to correctly classify this tooth to its tribe, genus and species, based on the shape and size of the remaining tooth.
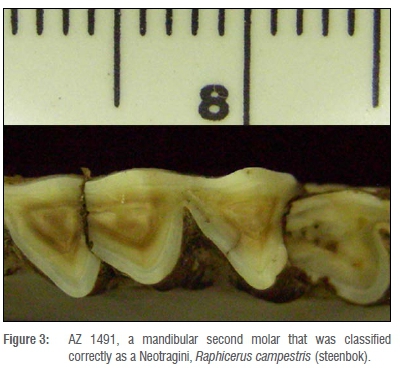
The second case, tooth AZ 1491, is a mandibular second molar that, despite its wear, was classified correctly as a Neotragini, Raphicerus campestris (steenbok) (Figure 3). The first and third molars have greatly affected the mesial and distal lobes of this second molar through interproximal wear. The mesial side of the tooth is essentially a straight line, parallel with the tooth row, because of impact with the first molar during eruption. The distal lobe curves as a result of the third molar impact. Despite the wear on the tooth, enough information was present for the program to classify the tooth correctly.
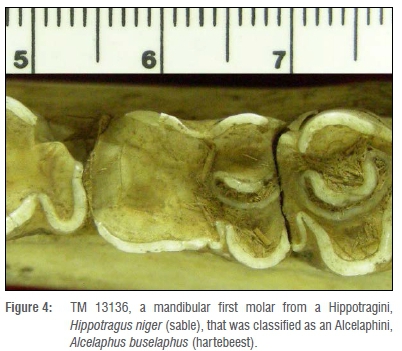
The third case is that of a mandibular first molar, TM 13136, belonging to a Hippotragini, Hippotragus niger (sable), which was classified as Alcelaphus buselaphus (hartebeest), an Alcelaphini (Figure 4). The mesial lobe of this tooth has been obliterated by wear and is missing the majority of its mesial and lingual enamel. The third molar has impacted the tooth, damaging the enamel, dentine and infundibulum of the distal lobe. Because of this wear, the tooth lacks the characteristics typical of Hippotragus, including a strong goat fold and pinching of the lateral/buccal lobes and strong ectostylid.23 A tooth that has lost this much enamel, together with the change in shape, was difficult for the program to identify. However, without the presence of the surrounding mandible and/or other associated material, this tooth would, arguably, have been difficult to classify by any means.
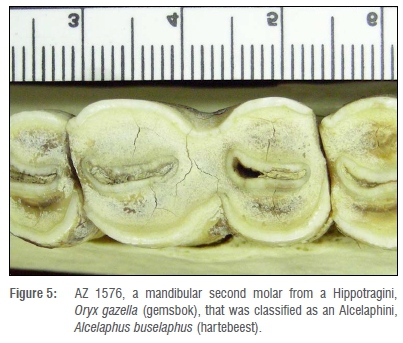
The last example of an incorrectly classified tooth is AZ 1576, a mandibular second molar of a Hippotragini, Oryx gazella (gemsbok), which was classified as an Alcelaphini, Alcelaphus buselaphus (Figure 5). This tooth exhibits late wear. The mesial enamel is worn away, leaving dentine that angles mesiolingually to distobucally. No goat fold is present; only a faint remnant of the parastylid remains. The distal lobe has been damaged by the third molar which has worn away the distolingual (entoconid) enamel and part of the distobuccal (hypoconid) enamel. A weak, barely visible ectostylid remains on the tooth. Because of wear, the tooth has lost some of the classic characteristics of hippotragine teeth and exhibits characteristics of alcelaphine teeth, including rounded lobes and the absence of a goat fold and ectostylid.23 This tooth was, therefore, difficult to classify using the EFA method.
Taking into consideration that wear remains an issue, the method presented here could assist in mitigating the challenge of taxonomically identifying worn bovid teeth. Valuable information - such as the number of individual specimens, minimum number of individuals and relative abundance of taxa - is lost when teeth in an assemblage are grouped into broad taxonomic levels due to their poor preservation. It is important to keep in mind that most bovid teeth in the fossil record are isolated. The classifications of the teeth in this study are known because they are complete dentitions in the maxilla/mandible, associated with horn cores and housed in museum collections. In the absence of such a context, teeth are especially difficult to identify. This program, however, provides a supplement to the identification of bovid teeth where ambiguity exists.
Conclusions
Worn teeth are more challenging to classify than unworn or less worn teeth. Elliptical Fourier analysis performed on the form of the occlusal surface of teeth with medium to late wear is a viable method for aiding in taxonomic identification; this finding is despite the fact that worn teeth do not classify at the same high probabilities as relatively unworn teeth. Although some tooth types (e.g. M1) tend to be more difficult to classify than others, and the teeth of some tribes overlap more in form when they are worn, this method aids in taxonomic classification where ambiguity in their identification exists. This research is particularly important when limited data exist on a specimen, such as an isolated tooth. By identifying more bovid teeth with higher confidence at a site, more individual specimens can be used in faunal lists. These data, in turn, are valuable for reconstructing past environments.
Acknowledgements
We thank the reviewers for taking the time to comment on our paper; Ditsong Museum, Stephanie Potze and Lazarus Kgasi for access to the collections and facilitating research; Bernhard Ziphel, Curator at the University of the Witwatersrand; and James Brink at the National Museum Bloemfontein.
Authors' contributions
J.K.B.: Conceptualisation, methodology, data collection, data analysis, data curation, writing - the initial draft. G.J.M.: Conceptualisation, methodology, data analysis, validation, data curation, writing - revisions. G.K.T.: Conceptualisation, methodology, writing - revisions.
References
1.Bobe R, Eck GG. Responses of African bovids to Pliocene climatic change. Paleobiology. 2001;27(2):1-48. https://doi.org/10.1666/0094-8373(2001)027<0001:ROABTP>2.0.CO;2 [ Links ]
2.Alemseged Z. An integrated approach to taphonomy and faunal change in the Shungura Formation (Ethiopia) and its implications for hominid evolution. J Hum Evol. 2003;44:451-478. https://doi.org/10.1016/S0047-2484(03)00012-5 [ Links ]
3.De Ruiter DJ, Brophy JK, Lewis PJ, Churchill SE, Berger LR. Faunal assemblage composition and paleoenvironment of Plovers Lake, a Middle Stone Age locality in Gauteng Province, South Africa. J Hum Evol. 2008;55:1102-1117. https://doi.org/10.1016/j.jhevol.2008.07.011 [ Links ]
4.Brophy JK, De Ruiter DJ, Athreya S, DeWitt TJ. Quantitative morphological analysis of bovid teeth and its implications for paleoenvironmental reconstructions in South Africa. J Arch Sci. 2014;41:376-388. https://doi.org/10.1016/j.jas.2013.08.005 [ Links ]
5.Vrba ES. Some evidence of the chronology and paleoecology of Sterkfontein, Swartkrans and Kromdraai from the fossil Bovidae. Nature. 1975;254:301-304. https://doi.org/10.1038/254301a0 [ Links ]
6.Vrba ES. The fossil record of African antelopes (Mammalia, Bovidae) in relation to human evolution and paleoclimate. In: Vrba ES, Denton GH, Partridge TC, Burckle LH, editors. Paleoclimate and evolution, with emphasis on human origins. New Haven, CT: Yale University Press; 1995. p. 385-424. https://doi.org/10.1017/s0016756897247710 [ Links ]
7.Brain CK. The hunters or the hunted. An introduction to African cave taphonomy. Chicago, IL: University of Chicago Press; 1981. https://doi.org/10.1002/ajpa.1330590320 [ Links ]
8.Brain CK. Temperature-induced environmental changes in Africa as evolutionary stimuli. In: Vrba ES, editor. Species and speciation. Transvaal Museum Monograph 4. Pretoria: Transvaal Museum; 1985. p. 45-52. [ Links ]
9.Watson V. Composition of the Swartkrans bone accumulations, in terms of skeletal parts and animals represented. In: Brain CK, editor. Swartkrans: A cave's chronicle of early man. Transvaal Museum Monograph 8. Pretoria: Transvaal Museum; 1993. p. 35-74. [ Links ]
10.Klein RG, Avery G, Cruz-Uribe K, Steele TE. The mammalian fauna associated with an archaic hominin skullcap and later Acheulean artifacts at Elandsfontein, Western Cape Province, South Africa. J Hum Evol. 2007;52:164-186. https://doi.org/10.1016/j.jhevol.2006.08.006 [ Links ]
11.Wells LH, Cooke HBS. Fossil Bovidae from the limeworks quarry, Makapansgat, Potgietersrus. Paleont Afr. 1956;4:1-55. [ Links ]
12.Reed KE. Using large mammal communities to examine ecological and taxonomic organization and predict vegetation in extant and extinct assemblages. Paleobiology. 1998;32:384-408. https://doi.org/10.1666/0094-8373(1998)024[0384:ULMCTE]2.3.CO;2 [ Links ]
13.Brophy JK. Reconstructing the habitat mosaic associated with Australopithecus robustus: Evidence from quantitative morphological analysis of bovid teeth [dissertation]. College Station, TX: Texas A&M University; 2011. [ Links ]
14.Matthews GJ, Brophy JK, Luetkemeier MP, Gu H, Thiruvathukal GK. Classification of bovid teeth: A comparison of machine learning techniques for taxonomic classification of teeth from the Family Bovidae. J App Stats. 2018;45(15):2773-2787. https://doi.org/10.1080/02664763.2018.1441381 [ Links ]
15.Matthews GJ, Thiruvathukal GK, Luetkemeier MP, Brophy JK. Examining the use of Amazon's Mechanical Turk for edge extraction of the occlusal surface of fossilized bovid teeth. PLoS ONE. 2017;12(7), e0179757, 18 pages. https://doi.org/10.1371/journal.pone.0179757 [ Links ]
16.Klein RG. On the taxonomic status, distribution and ecology of the blue antelope, Hippotragus leucophaeus (Pallas, 1766). Ann S Afr Mus. 1974;65:99-143. [ Links ]
17.Klein RG, Cruz-Uribe K. The bovids from Elandsfontein, South Africa, and their implications for the age, palaeoenvironment, and origins of the site. Afr Archaeol Rev. 1991;9:21-79. [ Links ]
18.Klein RG, Cruz-Uribe K, Beaumont PB. Environmental, ecological, and paleoanthropological implications of the late Pleistocene mammalian fauna from Equus Cave, Northern Cape Province, South Africa. Quat Res. 1991;36:94-119. [ Links ]
19.Lacruz RS, Brink JS, Hancox PJ, Skinner AR, Herries A, Schmid P, Berger LR. Palaeontology and geological context of a Middle Pleistocene faunal assemblage from the Gladysvale Cave, South Africa. Palaeont Afr. 2002;38:99-114. [ Links ]
20.Reynolds SC, Kibii JM. Sterkfontein at 75: Review of palaeoenvironments, fauna and archaeology from the hominin site of Sterkfontein (Gauteng Province, South Africa). Palaeont Afr. 2011;46:59-88. [ Links ]
21.Bonhomme V, Picq S, Gaucherel C, Claude J. Momocs: Outline analysis using R. J Stat Soft. 2014;56(13):1-24. https://doi.org/10.18637/jss.v056.i13 [ Links ]
22.Van Zyl LJ. Ageing criteria for some of the Bovidae in southern Africa [thesis]. Stellenbosch: Stellenbosch University; 1972. [ Links ]
23.Gentry AW. Evolution of African mammals. Cambridge, MA: Harvard University Press; 1978. https://doi.org/10.4159/harvard.9780674431263.c28 [ Links ]
 Correspondence:
Correspondence:
Juliet Brophy
Email: jbrophy@lsu.edu
Received: 21 Aug. 2018
Revised: 10 Oct. 2018
Accepted: 05 Apr. 2019
Published: 30 July 2019
EDITOR: Maryna Steyn
FUNDING: Louisiana State University
Supplementary Material
The open data set is available here: [Open data set]


{kind=link}