










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.114 n.9-10 Pretoria Sep./Oct. 2018
http://dx.doi.org/10.17159/sajs.2018/4235
RESEARCH ARTICLE
First report of bacterial endophytes from the leaves of Pellaea calomelanos in South Africa
Siphiwe G. Mahlangu; Mahloro H. Serepa-Dlamini
Department of Biotechnology and Food Technology, University of Johannesburg, Johannesburg, South Africa
ABSTRACT
Bacteria have an endosymbiotic association with plants. Previous studies have identified endophytic bacteria and their importance in biocontrol and drug development. However, most medicinal plants identified have not been assayed for bacterial endophytes. In this study, we characterised and identified bacterial endophytes from surface-sterilised leaves of Pellaea calomelanos, a common fern in the Limpopo and Gauteng Provinces, South Africa. Using morphological data and 16S rRNA gene sequencing, we differentiated and identified six putative endophytic bacteria, with Pantoea as the dominant genus; the other two identified bacteria belong to genera Arthrobacter and Bacillus. Data from this study are an addition to the previously less studied phylloplane bacteria. This study is a pilot in cataloguing bacterial endophytes from Pellaea calomelanos.
Significance:
• This study provides the first report of six putative bacterial endophytes from Pellaea calomelanos.
• Our results will pave the way for exploring the antimicrobial activity of P. calomelanos bacterial endophytes and whole genome comparisons between plant bacterial endophytes and plant bacterial pathogens.
Keywords: Pantoea; medicinal plant; phylogenetic analysis; biolog micro-station; 16S rRNA gene
Introduction
Plants have a mutualistic relationship with varied endophytes.1 Endophytes are endosymbiotic; they are often bacterial or fungal species which colonise the plants without causing harm or pathogenic infection3 and can spend part or their entire life cycle within the plant host2-4.
Most plant species are known to host one or more endophytic microorganisms.5 Endophytes form a symbiotic relationship with the plants by providing a biological defence mechanism for the plant host against pathogens,2-5 through the production of secondary metabolites. These metabolites halt the growth of or attack invading antagonists or lyse-infected plant cells; furthermore, the metabolites can induce plant host defence mechanisms and promote plant growth.6
The present study was designed to isolate, identify and characterise endophytic bacteria from Pellaea calomelanos obtained in South Africa. Pteridaceae is a family of fern plants with over 45 genera and more than 1000 species.7 One such genus is Pellaea with over 35 described species found growing in arid rocky regions and within narrow open pockets in the soil.7,8P. calomelanos is a common fern species in the Limpopo and Gauteng Provinces of South Africa, but also grows throughout the rest of the country.9 Common names of P. calomelanos in South Africa include inkomankomo (Zulu), lehorometso (Sotho), legogoana (Tswana), phalatjane (Sepedi) and hard fern (English).9
P. calomelanos is a multipurpose medicinal plant used for the treatment of headaches, chest colds, asthma, head colds and mouth and nasal ulcers.9 Like other plants, Pellaea species have a mutualistic relationship with endophytes, although, to date, endophytes have been reported in only P. concolor and P. viridis.10,11 We thus report here on the occurrence of bacterial endophytes within the leaves of P. calomelanos. Ours is the first study to describe the isolation, identification and characterisation of bacterial endophytes from P. calomelanos using morphological and phenotypic characteristics and the sequencing of the 16S rRNA gene.
Materials and methods
Collection of plant material
Aerial portions of the plant material were collected from Botlokwa (23°29'34.8'S, 29°42'11.2'E) in the Limpopo Province of South Africa. Whole plants were placed in sterile polyethylene bags and transported to the laboratory under 4 °C. The plant material was collected in March 2017 from a site with sandy loam soil.
Identification of the plant material
The identification of the plant material was carried out at the University of Johannesburg Herbarium (JRAU). A sample specimen of the plant material was deposited in the University of Johannesburg Herbarium (JRAU) with voucher specimen number Serepa-Dlamini 201 and species name Pellaea calomelanos. The remaining collected plant material was immediately processed in the laboratory.
Isolation of endophytic bacteria
Immediately after collection of plant material in the lab, plant leaves were washed with running tap water followed by a sequential sterilisation with the following solutions: sterile distilled water for 1 min, 70% ethanol for 1.5 min, 1% sodium hypochlorite for 3 min and finally washed three times in sterile distilled water. The final washing water was then plated as control. The surface-sterilised leaves were ground in 2 mL of saline using a pestle and mortar. Under sterile conditions, the homogenate was streaked onto nutrient agar plates. The plates were incubated for 24-48 h at 28 °C and inspected daily for bacterial colony growth. Isolated colonies were re-cultured on sterile nutrient agar plates until pure colonies with uniform morphology were obtained. For each endophyte bacterial isolate, 35% glycerol (glycerol diluted in sterile distilled water) stock cultures were prepared and stored at -80 °C for future use.
Preliminary identification of endophytic bacterial isolates
Pure colonies were subjected to a Gram reaction test.12 The Schaeffer-Fulton stain method13 was conducted to determine if the bacterial endophytes produced endospores. The hanging-drop method adapted from MacFaddin14 was used to determine motility of the bacterial isolates. All the prepared slides were examined using a bright-field compound light microscope (CX21FS1, Olympus Corporation, Tokyo, Japan) with 400x and 1000x magnification.
Identification of bacterial endophytes by the Biolog MicroPlate system
The following procedure was utilised to identify the putative endophyte bacterial isolates. Under sterile conditions, overnight single colonies of the isolates were sub-cultured in six separate 5 mL of 0.85% saline solution. The Biolog turbidimeter was used to monitor and measure the turbidity of the suspension until a 90-98% transmittance was reached. The suspension (150 µL) was aliquoted into each well of the Biolog MicroPlate (with GEN III MicroPlate™; Biolog Inc., Hayward, CA, USA) and incubated at 26 °C for 24 h. The plates were scanned using the Biolog automatic system and samples identified using the Biolog software.15
Scanning electron microscopy
A scanning electron microscope was used to observe and study the shape of the endophyte bacterial isolates. Each glycerol stock of the isolated bacterial endophytes was inoculated into 5 mL nutrient broth and incubated for 48 h at 28 °C. The bacterial suspension was centrifuged at 2935 rcf for 10 min, the supernatant was discarded and bacterial cells were rinsed with sterile distilled water three times with a 5-min interval between each rinse. The bacterial cells were then fixed in 8% glutaraldehyde overnight (glutaraldehyde 25% EM grade diluted in Ringers' solution). Sterile distilled water was used to rinse the cells twice, followed by a series of dehydration with 30%, 50%, 70%, 90%, 95% and 100% ethanol at 10-min intervals and a centrifugation step at 2935 rcf for 10 min. The bacterial pellets were left to dry overnight in open Eppendorf tubes placed in a refrigerator at 4 °C. The cells were then mounted on scanning electron microscope stubs and coated with gold and viewed using the TESCAN VEGA 3 LMH (AVG9731276ZA, Warrendale, PA, USA) scanning electron microscope fitted with a digital camera.
Identification of bacterial endophytes using 16S rRNA
Extraction of genomic DNA
Genomic DNA of each bacterial isolate was extracted from nutrient agar pure colonies using a Quick-DNATM ZR fungal/bacterial DNA MiniPrep kit (Zymo Research, Irvine, CA, USA) according to the manufacturer's protocol. The extracted DNA was quantified using the NanoDrop ND-2000 UV-Vis spectrophotometer (ThermoFisher Scientific, USA); the DNA concentration was 40 ng/µL prior to cleaning with ZR fungal/bacterial DNA clean and concentrator-5 (Zymo Research).
Polymerase chain reaction amplification and sequencing
The 16S rRNA gene of each bacterial isolate was amplified by polymerase chain reaction (PCR) in a 12.5-µL reaction containing 1.5 µL of template DNA, 1.5 µL of each primer 5`-AGAGTTTGATCCTGGCTCAG-3`f and 5`-AAGGAGGTGATCCAAGCCGCA-3`r, 6 µL of One Taq® 2X PCR master mix with standard buffer (20 mM Tris-HCI, 1.8 mM MgCl2, 22 mM NH4Cl, 22 mM KCl, 0.2 mM dNTPs, 5% glycerol, 0.06% IGEPAL® CA-630, 0.05% Tween® 20, 25 units/mL One Taq® DNA polymerase) and final volume filled up to 12.5 µL with nuclease free water. The PCR cycle was performed using MyCyclerTM Thermal Cycler (catalogue number 580BR 08389, BioRad, Hercules, CA, USA) with the following conditions: initial denaturation at 94 °C for 3 min, followed by 35 cycles of amplification: denaturation at 94 °C for 1 min, 55 °C for 1 min (annealing), 72 °C for 2 min (extension), followed by a final extension at 72 °C for 10 min.16 The PCR products were cleaned with ExoSAP-it™ (ThermoFisher Scientific) following the manufacturer's recommendations and sent for sequencing with primers to Inqaba Biotechnical Industries (Pty) Ltd, Pretoria, South Africa.
Phylogenetic analysis
The 16S rRNA gene sequences of the bacterial isolates were subjected to BLAST (v.2.6.0) analysis against the rRNA sequence database (Bacteria and Archaea) at the National Center for Biotechnology Information (NCBI) to identify the closest related bacterial species. Only bacterial species with 95-100% identity similarity were selected for phylogenetic analysis. The aligned sequences, containing the isolate and closest related bacterial species, were determined by MUSCLE17 and phylogenetic analysis carried out using the maximum likelihood method based on the Tamura-Nei model17. Positions overlapping with gaps and missing nucleotide data were eliminated. All evolutionary analyses were conducted in MEGA 7.18Escherichia coli ATCC 11775T with GenBank accession number X80725 was used as the outgroup. The phylogenetic trees were reported with bootstrap percentages. The 16S rRNA gene sequences of bacterial isolates identified in the study were deposited in GenBank (https://www.ncbi.nlm.nih.gov/genbank/) with the following accession numbers and names: MF613647 (Arthrobacter sp. strain MHSD1), MF613648 (Pantoea sp. strain MHSD2), MF613649 (Bacillus infantis strain MHSD3), MF613650 (Pantoea sp. strain MHSD4), MF613651 (Pantoea ananatis strain MHSD5) and MF613652 (Pantoea sp. strain MHSD6). The assigned names of the bacterial isolates were based on the BLAST homology percentages as well as phylogenetic results.
Results and discussion
Isolation and identification of endophytic bacteria
Morphological identification
The isolation and enumeration of endophytes from surface-sterilised plant material is the recommended method.3,4,19-23 In the current study, six different putative bacterial endophytes were isolated from the surface-sterilised leaves of P. calomelanos obtained from Botlokwa, Limpopo Province, South Africa. Previous studies on bacterial endophytes have focused on rhizosphere bacterial endophytes because of their major role in nutrient uptake and high diversity in soils.19-21
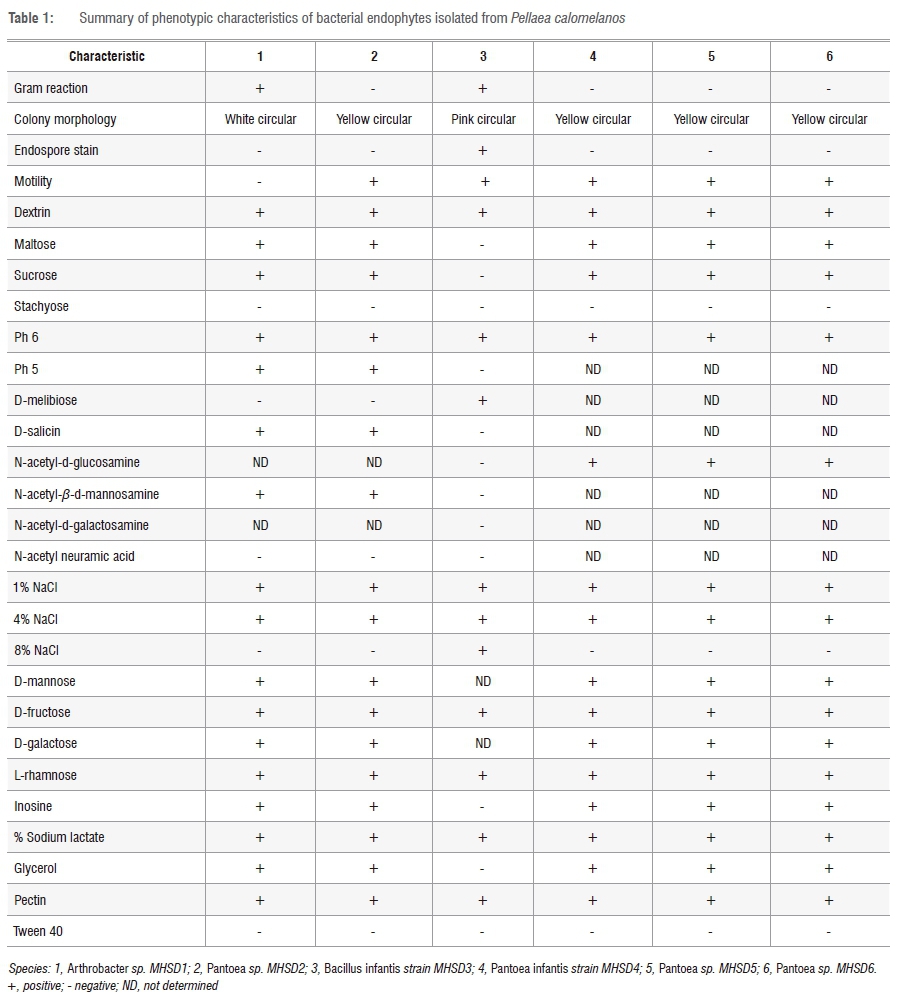
Preliminary morphological observations of the six isolated colonies that included the production of endospores, motility and Gram staining has enabled the grouping of the bacterial cultures into various groups, some with similar characteristics as shown in Table 1. The Gram stain results indicated that all bacteria were rod shaped and two out of the six isolates were Gram positive and the remaining four isolates were Gram negative. Only one of the Gram-positive isolates was an endospore former. All the isolates, except one Gram-positive isolate, exhibited motility. The morphological shapes were confirmed by the scanning electron microscope results (not shown) which showed uniform cells depicting pure cultures of the bacterial isolates.
Biolog MicroPlates
The phenotypic assays were performed on the isolates using the Biolog MicroPlates (GEN III MicroPlate) and the results are presented in Table 1. The bacterial isolates can be differentiated based on the utilisation of some of the carbon sources such as dextrin, maltose, sucrose, stachyose and pectin. Most of the isolates appear to hydrolyse these carbon sources, although variations were observed. Some of the results were not determined (ND), such as utilisation of N-acetyl-d-glucosamine, N-acetyl-β-d-mannosamine and N-acetyl neuraminic acid, and thus we cannot rely solely on the phenotypic tests for variation of the bacterial isolates.
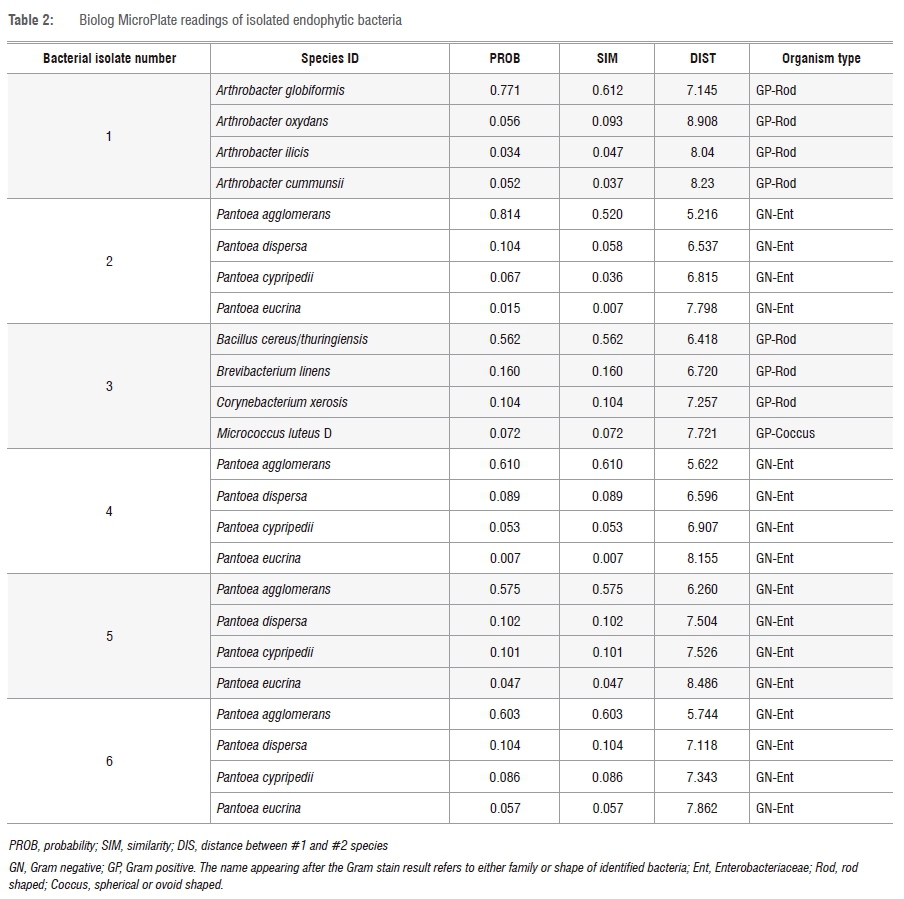
The Biolog system can identify bacterial species both at genus and species levels, as well as through Gram stain reaction. The system does so by providing four top-ranked species for identification; Table 2 indicates that isolates 2, 4, 5 and 6 were Gram negative and belonged to the Enterobacteriaceae family. Although the species ID levels for these isolates had different species names for each (of the four top-ranked ID species), they all were identified to be of the genus Pantoea. The Gram reaction results obtained here correlate with those performed initially in the study. Isolates 1 and 3 were also predicted to be Gram positive; however, the family names as well as the genus and species names differed among each of the four predicted species names. This difference could be because we utilised a different growth medium from the one recommended in the MicroStation™ System/MicroLog User Guide.15
The MicroStation™ System/MicroLog User Guide15 states that if the top four identified species belong to the same or closely related genera, then the identification can be concluded as a positive result. The other parameters that can be considered from the identified results are: the probability (PROB), similarity (SIM) and distance (DIST). These parameters indicate the approximate degree of matching between the MicroPlate results and the corresponding database. SIM ≥ 0.5, DIST ≤ 5.0 and PROB close to 1 indicate reliability of the test results. Although the SIM and DIST results of the top ranked ID species for each isolate were greater than 0.5 and less than 5.0, respectively, the PROB for all isolates was at 0.7 or above 0.7, which is relatively close to 1, thus indicating reliable results.
Although the variation of phenotypic tests could not be concluded from the Biolog MicroPlates (GEN III MicroPlate), the system has shown to be reliable when identifying bacterial species to genus level; further identification can be supported by Gram stain reaction and colony morphology. We do, however, recommend that the use of Biolog MicroPlates (GEN III MicroPlate) be complemented with other strain identification methods, despite the current study showing the reliability of identifying bacterial species by use of the Biolog MicroPlates (GEN III MicroPlate). Furthermore, previous studies have identified and examined phenotypic characteristics of bacterial species utilising this system.24-27
Phylogenetic analysis
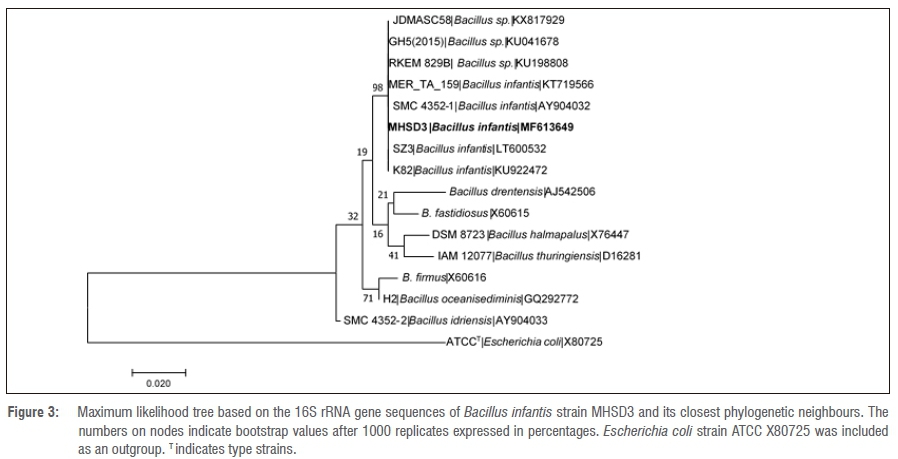
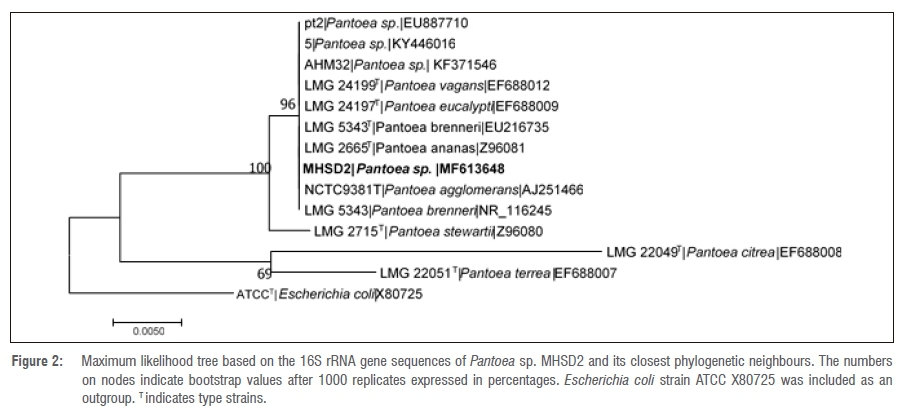
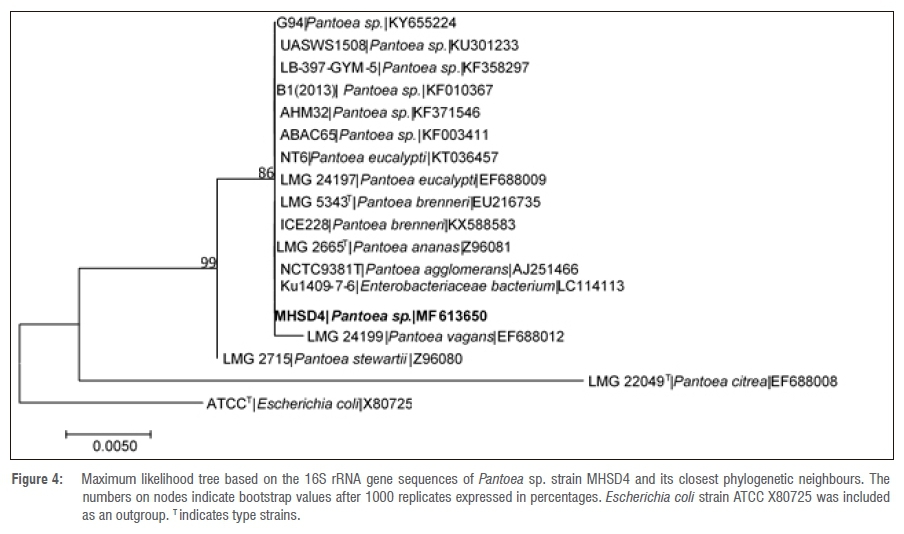
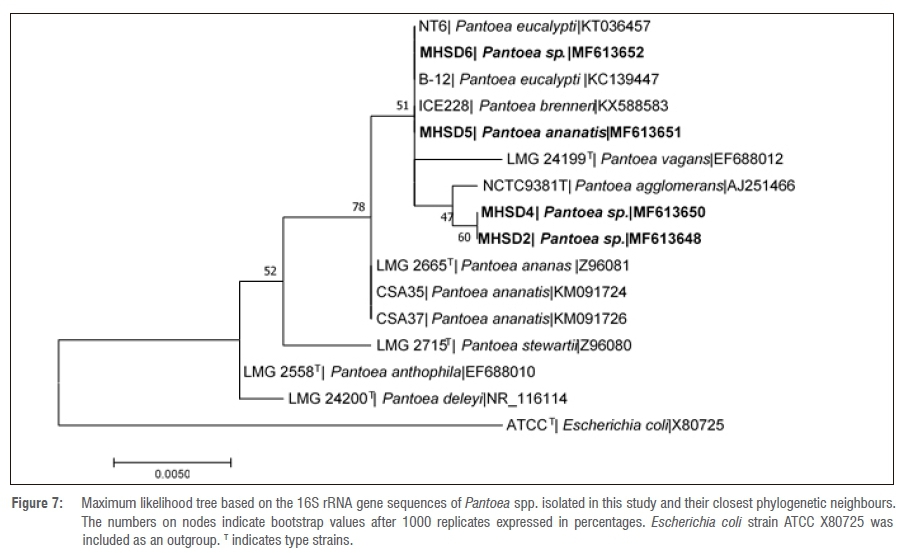
The 16S rRNA gene sequence lengths (in base pairs) are shown in Table 3. The NCBI BLAST search results indicated that bacterial endophyte isolate number 1 had a 100% identity similarity with Arthrobacter spp., bacterial endophyte isolate 3 had 100% identity similarity with Bacillus spp. and bacterial endophyte isolates 2, 4, 5 and 6 had 99% identity similarities with Pantoea spp. These results indicate that the dominating endophytic bacteria from Pellaea calomelanos belong to the genus Pantoea. The homology percentage identity was performed among all the bacterial isolates (results not shown). The lowest homology similarity percentage was observed between Arthrobacter vs Bacillus, Arthrobacter vs Pantoea and Bacillus vs Pantoea isolates, indicating that these are different species. There was, however, a 99% homology similarity between all the Pantoea isolates. These isolates were further resolved with phylogenetic analysis (Figures 1-6).
All the delineated phylogenetic trees for the isolates had polytomy relations with closely related species. Isolate 1 had a polytomy relationship with undescribed Arthrobacter species and Arthrobacter polychromogenes (Figure 1) with a 47% bootstrap value. Isolate 3 had a polytomy relationship supported by a 98% bootstrap value with undescribed Bacillus species and other strains of Bacillus infantis (Figure 3). Isolates 2, 4, 5 and 6 had polytomy relationships with closely related Pantoea species and all showed varying bootstrap values - the phylogenetic relationships were different for each isolate indicating that these are different Pantoea species (Figures 2, 4, 5 and 6). A delineated tree including all the isolated Pantoea species showed that Pantoea sp. MHSD2 and Pantoea sp. MHSD4 had a monophyletic relation supported by a 60% bootstrap value, whereas Pantoea sp. MHSD5 and Pantoea sp. MHSD6 had a polytomy relationship with Pantoea eucalypti and P. brenneri species (Figure 7). The 16S rRNA gene sequence is the foremost molecular marker used to identify bacterial species; however, from the results obtained, it is evident that 16S rRNA does not resolve the phylogenetic and evolutionary relationships between closely related bacterial strains. The phylogeny of these closely related bacterial strains can be resolved with the use of multilocus sequence analysis.28
We isolated bacterial endophytes from only the leaves and we believe the results obtained here will be additional to the minimally reported number of phylloplane bacterial endophytes.4 Braithwaite et al.29 have reported on the antimicrobial activity of P. calomelanos against bacterial pathogens and yeast. Thus, in exploring the antimicrobial activity and other potential applications of bacterial endophytes from this plant, we had to identify and understand its associated bacterial endophytes.
Pantoea spp., Bacillus spp. and Arthrobacter spp. are common soil bacteria. Furthermore, these bacterial species have been previously isolated from maize, rice and medicinal plants as bacterial endophytes.30-33 Although they are considered to be plant endophytes, bacteria from the same genera as the reported bacterial endophytes have also been isolated from diseased plants and clinical samples.27,34 From these reports and observations, there is a need to understand the evolutionary relationships of these bacterial species, and also the occurrences and differences between bacterial endophytes as plant endophytes and as phytopathogens. Therefore, we recommend that whole genome sequencing and comparisons can provide more comprehensive insights to resolve the evolutionary relationships and identify the bacterial groups to strain level, as well as to identify genetic components that prompt the occurrence of bacterial species as bacterial endophytes.
Soil type35, season36,37 and host type38,39 have been reported to affect the diversity and seasonal fluctuations of bacterial endophytes. These factors could explain the low number of bacterial endophytes isolated from the current study. Furthermore, the Pantoea genus was dominant, with four identified Pantoea species, and we strongly believe that more bacterial endophytes are likely associated with P. calomelanos. Further studies are underway to isolate and identify bacterial endophytes from P. calomelanos collected in different seasons and from different soil types. Methods which are culture independent can also be employed in identifying bacterial endophytes.
Bacterial endophytes produce the same or similar metabolites as their hosts. Therefore, because P. calomelanos has antibacterial and antifungal activities, the ability of bacterial endophytes from this plant to produce metabolites which (1) have therapeutic activity, (2) are similar to those produced by P. calomelanos and (3) have other possible potential applications, needs to be studied.
Conclusion
Bacterial endophytes from P. calomelanos are poorly studied. The current study provides information on the isolation and diversity of bacterial endophytes from P. calomelanos. This study is a pilot to ongoing research on P. calomelanos obtained in South Africa, on its secondary metabolites and bacterial endophytes and potential application of the secondary metabolites and bacterial endophytes. Furthermore, whole genomic studies are underway to understand the evolutionary relationships between bacteria that occur as plant endophytes and plant pathogens. The genomic components that drive the symbiosis between P. calomelanos and its bacterial endophytes will also form part of future studies.
Acknowledgements
We thank the Faculty Research Committee and University Research Committee of the University of Johannesburg for funding this research. We are thankful to Mr Rudzani Manafe and Mr Witness Qaku for their technical support. Mr Siphiwe G. Mahlangu received the NRF Scarce Skills Master's scholarship (grant number SFH160628174791).
Authors' contributions
M.H.S-D. conceptualised the research; S.G.M. conducted the experiments; both authors analysed the data and contributed to the writing and editing of the manuscript.
References
1. Guo LD, Hyde KD, Liew ECY. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000;147:617-630. https://doi.org/10.1046/j.1469-8137.2000.00716.x [ Links ]
2. Chojniak J, Jalowiecki L, Dorgeloh E, Hegedusova B, Ejhed H, Magner J, et al. Application of the BIOLOG system for characterization of Serratia marcescens ss marcescens isolated from onsite wastewater technology (OSWT)*. Acta Biochim Pol. 2015;62(4):799-805. https://doi.org/10.18388/abp.2015_1138 [ Links ]
3. Toghueo RMK, Zabalgogeazcoa I, Vazquez de Aldana BR, Boyom FF. Enzymatic activity of endophytic fungi from medicinal plants Terminalia catappa, Terminalia mantaly and Cananga odorata. S Afr J Bot. 2017;109:146-153. https://doi.org/10.1016/j.sajb.2016.12.021 [ Links ]
4. Gouda S, Das G, Sen SK, Shin H, Patra JK. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front Microbiol. 2016;7, Art. #1538, 8 pages. https://doi.org/10.3389/fmicb.2016.01538 [ Links ]
5. Strobel GA. Endophytes as sources of bioactive products. Microb Infect. 2003;5(6):535-544. https://doi.org/10.1016/S1286-4579(03)00073-X [ Links ]
6. Ding T, Melcher U. Influences of plant species, season and location of leaf endophytic bacterial communities of non-cultivated plants. PLoS ONE. 2016;11(3), e0150895, 13 pages. https://doi.org/10.1371/journal.pone.0150895 [ Links ]
7. Schuettplez E, Schneider H, Huiet L, Windham MD, Pryer KM. A molecular phylogeny of the fern family Pteridaceae: Assessing overall relationships and the affinities of previously unsampled genera. Mol Phylogenet. Evol. 2007;44(3):1172-1185. https://doi.org/10.1016/j.ympev.2007.04.011 [ Links ]
8. Alan RS, Kathleen MP, Eric S, Peter K, Harald S, Paul GW. A classification for extant ferns. Taxon. 2007;55(3):705-731. [ Links ]
9. South African National Biodiversity Institute (SANBI). Pellaea calomelanos [homepage on the Internet]. c2017 [cited 2017 Jul 30]. Available from: http://pza.sanbi.org/pellaea-calomelanos [ Links ]
10. Muthukumar T, Prabha K. Aubuscular mycorrhizal and septate endophyte fungal associations in lycophytes and ferns of south India. Symbiosis. 2013;59(1):15-33. https://doi.org/10.1007/s13199-012-0185-z [ Links ]
11. Tarnau K, Anielska T, Jurkiewicz A. Mycothallic/mycorrhizal symbiosis of chlorophyllous gametophytes and sporophytes of a fern, Pellaea viridis (Forsk.) Prantl (Pellaeaceae, Pteridales). Mycorrhiza. 2005;15(2):121-128. https://doi.org/10.1007/s00572-004-0306-5 [ Links ]
12. Cruikshank R, Duguid JP, Marmoin BP, Swain RH. Medical microbiology. 12th ed. New York: Longman Group Limited; 1975. [ Links ]
13. Prescott LM, Harley JP, Klein DA. Laboratory exercises in microbiology. 5th ed. New York: McGraw-Hill; 2002. [ Links ]
14. MacFaddin J. Biochemical tests for identification of medical bacteria. 3rd ed. Philadelphia, PA: Lippincott Williams & Wilkins; 2000. [ Links ]
15. MicroStation™ System/MicroLog™ user guide. A powerful automated system for the rapid identification of bacteria, yeasts and filamentous fungi. Hayward, CA; Biolog; 2010. [ Links ]
16. Yeates C, Gillings MR, Davison AD, Altavilla N, Veal DA. PCR amplification of crude microbial DNA extracted from soil. Lett Appl Microbiol.1997;25:303-307. https://doi.org/10.1046/j.1472-765X.1997.00232.x [ Links ]
17. Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993;10:512-526. [ Links ]
18. Kumar S, Stecher G, Tamura K. MEGA 7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870-1874. https://doi.org/10.1093/molbev/msw054 [ Links ]
19. Jasim B, Joseph AA, John CJ, Mathew J. Isolation and characterization of plant growth promoting endophytic bacteria from the rhizome of Zingiber officinale. 3 Biotech. 2014;4(2):197-204. https://doi.org/10.1007/s13205-013-0143-3 [ Links ]
20. Long HH, Schmidt DD, Baldwin IT. Native bacterial endophytes promote host growth in a species-specific manner; phytohormone manipulations do not result in common growth responses. PLoS One. 2008;3(7), e2702, 10 pages. https://doi.org/10.1371/journal.pone.0002702 [ Links ]
21. Ashkan MF, Bleakly B. Isolation, characterization and identification of putative bacterial endophytes from some plants in Hot Springs, South Dakota. Int J Curr Microbiol App Sci. 2017;6(6):756-767. https://doi.org/10.20546/ijcmas.2017.606.089 [ Links ]
22. Fouda AH, Hassan SE, Eid AM, Ewais EE. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Ann Agric Sci. 2015;50(1):95-104. https://doi.org/10.1016/j.aoas.2015.04.001 [ Links ]
23. Azevedo JL, Quecine MC. Diversity and benefits of microorganisms from the tropics. Zug, Switzerland: Springer International Publishing AG; 2017. https://doi.org/10.1007/978-3-319-55804-2 [ Links ]
24. Wu W, Yang F, Bai L. Influence of substrate on electricity generation of Shewanella loihica PV-4 in microbial fuel cells. Microb Cel Fact. 2014;13, Art. #69, 6 pages. https://doi.org/10.1186/1475-2859-13-69 [ Links ]
25. Pyar H, Peh K. Characterization and identification of Lactobacillus acidophilus using Biolog rapid identification system. Int J Pharm Sci. 2014;6(1):189-193. [ Links ]
26. Stoyanova M, Vancheva T, Moncheva P, Bogatzevska N. Differentiation of Xanthomonas spp. causing bacterial spot in Bulgaria based on Biolog system. Int J Microbiol. 2014;2014, Art. #495476, 7 pages. https://doi.org/10.1155/2014/495476 [ Links ]
27. Brady CL, Venter SN, Cleenwerck I, Engelbeen K, Vancanneyt M, Swings J, et al. Pantoea vagans sp. Nov., Pantoea eucalypti sp. Nov., Pantoea deleyi sp. Nov and Pantoea antophila sp. Nov. Int J Syst Evol Microbiol. 2009;59:2339-2345. https://doi.org/10.1099/ijs.0.009241-0 [ Links ]
28. Grevers D, Cohan FM, Lawrence JG, Spratt BG, Coenye T, Feil EJ, et al. Re-evaluating prokaryotic species. Nat Rev Microbiol. 2005;3:733-739. https://doi.org/10.1038/nrmicro1236 [ Links ]
29. Braithwaite M, Van Vuuren SF, Viljoen AM. Validation of smoke inhalation therapy to treat microbial infections. J Ethnopharmacol. 2008;119(3):501-506. https://doi.org/10.1016/j.jep.2008.07.050 [ Links ]
30. Xiong XQ, Liao HD, Liu XM, Zhang LY, Shi XW, Yang XL, et al. Isolation of a rice endophytic bacterium, Pantoea sp. Sd-1 with ligninolytic activity and characterization of its rice straw degradation ability. Lett Appl Microbiol. 2013;58:123-129. https://doi.org/10.1111/lam.12163 [ Links ]
31. Sheibani-Tezerji R, Naveed M, Jehl MA, Sessitsch A, Rattei T, Mitter B. The genomes of closely related Pantoea ananatis maize seed endophytes having different effects on the host plant differ in secretion system genes and mobile genetic elements. Front Microbiol. 2015;6, Art. #440, 16 pages. https://doi.org/10.3389/fmicb.2015.00440 [ Links ]
32. Egamberdieva D, Wirth S, Behrendt U, Ahmad P, Berg G. Antimicrobial activity of medicinal plants correlates with the proportion of antagonistic endophytes. Front Microbiol. 2017;8, Art. #199, 11 pages. https://doi.org/10.3389/fmicb.2017.00199 [ Links ]
33. Loh CY, Tan Y, Rohani R, Weber JF, Bhore SJ. Diversity of endophytic bacteria in Malaysian plants as revealed by 16S rRNA encoding gene sequence based method of bacterial identification. J Young Pharm. 2013;5:95-97. https://doi.org/10.1016/j.jyp.2013.07.001 [ Links ]
34. Brady C, Cleenwerck I, Venter S, Vancanneyt M, Swings J, Coutinho T. Phylogeny and identification of Pantoea species associated with plants, humans and the natural environment based on multilocus sequence analysis (MLSA). Syt Appl Microbiol. 2008;31:447-460. https://doi.org/10.1016/j.syapm.2008.09.004 [ Links ]
35. Hardoim PR, Hardoim CC, Van Overbeek LS, Van Elsas JD. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS One. 2012;7(2), e30438, 13 pages. https://doi.org/10.1371/journal.pone.0030438 [ Links ]
36. Forchetti G, Masciarelli O, Izaguirre MJ, Alemano S, Alvarez D, Abdala G. Endophytic bacteria improve seedling growth of sunflower under water stress, produce salicylic acid, and inhibit growth of pathogenic fungi. Curr Microbiol. 2010;61:485-493. https://doi.org/10.1007/s00284-010-9642-1 [ Links ]
37. Gao XX, Zhou H, Xu DY, Yu CH, Chen YQ, Qu LH. High diversity of endophytic fungi from the pharmaceutical plant, Heterosmilax japonica Kunth revealed by cultivation-independent approach. FEMS Microbiol Lett. 2005;249:255-266. https://doi.org/10.1016/j.femsle.2005.06.017 [ Links ]
38. Ding T, Palmer MW, Melcher U. Community terminal restriction fragment length polymorphisms reveal insights into the diversity and dynamics of leaf endophytic bacteria. BMC Microbiol. 2013;13, Art. #1, 11 pages. https://doi.org/10.1186/1471-2180-13-1 [ Links ]
39. Hunter PJ, Hand P, Pink D, Whipps JM, Bending GD. Both leaf properties and microbe-microbe interactions influence within-species variation in bacterial population diversity and structure in the lettuce (Lactuca species) phyllosphere. Appl Env Microbiol. 2010;76(24):8117-8125. https://doi.org/10.1128/AEM.01321-10 [ Links ]
 Correspondence:
Correspondence:
Mahloro Serepa-Dlamini
Email: hopes@uj.ac.za
Received: 16 Oct. 2017
Revised: 18 Jan. 2018
Accepted: 21 May 2018
Published: 11 Sep. 2018
FUNDING: University of Johannesburg


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}