










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.114 no.5-6 Pretoria may./jun. 2018
http://dx.doi.org/10.17159/sajs.2018/20170288
RESEARCH ARTICLE
Arable agriculture changes soil microbial communities in the South African Grassland Biome
Gilbert Kamgan NkuekamI; Don A. CowanI; Angel ValverdeI, II
ICentre for Microbial Ecology and Genomics, Department of Genetics, University of Pretoria, Pretoria, South Africa
IIDepartment of Microbial, Biochemical and Food Biotechnology, University of the Free State, Bloemfontein, South Africa
ABSTRACT
Many studies, mostly in temperate regions of the northern hemisphere, have demonstrated that agricultural practices affect the composition and diversity of soil microbial communities. However, very little is known about the impact of agriculture on the microbial communities in other regions of the world, most particularly on the African continent. In this study, we used MiSeq amplicon sequencing of bacterial 16S rRNA genes and fungal ITS regions to characterise microbial communities in agricultural and natural grassland soils located in the Mpumalanga Province of South Africa. Nine soil chemical parameters were also measured to evaluate the effects of edaphic factors on microbial community diversity. Bacterial and fungal communities were significantly richer and more diverse in natural grassland than in agricultural soils. Microbial taxonomic composition was also significantly different between the two habitat types. The phylum Acidobacteria was significantly more abundant in natural grassland than in agricultural soils, while Actinobacteria and the family Nectriaceae showed the opposite pattern. Soil pH and phosphorus significantly influenced bacterial communities, whereas phosphorus and calcium influenced fungal communities. These findings may be interpreted as a negative impact of land-use change on soil microbial diversity and composition.
SIGNIFICANCE:
• This report is the first of the effect of land-use changes on the diversity of the soil microbial communities in African grassland soils.
• Land-use changes influence the diversity and structure of soil microbial communities in the Grassland Biome of South Africa.
• This study serves as a baseline for future studies on South African soil microbial diversity.
Keywords: land use; grassland; agriculture; microbial diversity; next-generation sequencing
Introduction
Soils represent a reservoir for a wide diversity of microorganisms such as bacteria, fungi and viruses.1 Bacteria and bacteriophages are typically the most abundant microorganisms present in soils, although their prevalence is highly variable and affected by edaphic factors such as soil mineral content and pH1,2 and by local plant biodiversity3. Soil microbial communities are important drivers of ecosystem functioning and climate change mitigation through the fixation, immobilisation and cycling of greenhouse gases.4 Plants synthesise organic matter via photosynthetic activities and provide energy to soil microbes through root exudates.5,6 In return, soil microorganisms provide the plants with critical 'services', including decomposition of organic matter, mineral cycling and biocontrol of soil-borne pathogens.4,7
Land use, such as the modification of a natural ecosystem for agriculture purposes, has been proven to have significant effects on soil microbial communities by changing the physical and chemical properties of the soil. Such effects on the microbial community include changes in microbial species abundance, richness and diversity.8-11The effect of changing land use on the diversity of soil microbes has also been observed in grassland ecosystems. For example, Acosta-Martinez et al.12 studied soil bacteria diversity in a single soil type in Texas, USA. They found much higher bacteria diversity in soils under agriculture than in natural grassland soils, providing evidence of the positive influence of land use on soil bacterial diversity. The overall bacterial community diversity and composition in different grassland soils from across South and North America were significantly influenced with soil pH.13Similar results were reported in a study of bacterial communities in German grassland and forest soils.14,15 In a more recent study, both bacterial and fungal communities exhibited contrasting beta diversity among two types of European subalpine/alpine grasslands, and both bacterial and fungal communities were influenced by grassland type.16 However, most of these studies have been conducted in temperate northern hemisphere grasslands and none is directly relevant to the unique and defined biomes of South Africa.17 We argue that more geographically diverse comparative analyses are required in order to better understand how microbial communities are altered by land use.
South Africa is one of the world's biodiversity hotspots, and is ranked third in the global list; it is composed of nine biomes which together contain between 250 000 and 1 000 000 species including animals and plants, many of which are endemic to the country.17 However, while higher eukaryotes have been intensively studied in southern Africa, there has been only very limited focus on the biodiversity of soil microbial communities in any of South Africa's major biomes. Mpumalanga is a province in the northeast of South Africa, much of which is designated as Grassland Biome.17 The Grassland Biome is the cornerstone of commercial maize cropping, and many grassland areas have been converted to arable production. Sorghum, wheat and sunflowers are also farmed in the region but on a smaller scale.18 Agriculture is, therefore, one of the important economic sectors in Mpumalanga, with significant contributions toward national economic growth19, but with major modifications to much of the original natural grassland habitat.
In this study, we evaluated the differences in bacterial and fungal communities across a series of paired agricultural and natural grassland soils in the Mpumalanga Province, South Africa. We used an amplicon sequencing based approach to characterise the soil bacterial and fungal communities and to quantify their differences across 15 sites, and related these differences to measured soil chemical parameters. Inventories of the composition and differences in the bacterial and fungal communities in Mpumalanga's agricultural and natural grassland soils will contribute to the body of knowledge, and can provide a description of the core soil microbial communities in the unmodified grassland.
Materials and methods
Sample collection
The study sites are situated on the Highveld escarpment in the Mpumalanga Province, which makes up 6.5% of South Africa's land area, and form part of the Grassland Biome.20 The climatic conditions of the Grassland Biome of South Africa span a rainfall gradient from about 400 mm to >1200 mm per year, a temperature gradient from frost-free to snow-bound in winter, and an altitude from sea level to >3300 m; a spectrum of soil types occurs in the Grassland Biome, from humic clays to poorly structured sands.21 In April 2016, 15 paired agricultural and natural sites in the Grassland Biome were localised using a geographic information system (GIS) technology (Supplementary table 1). The distance between sampling sites spanned 18-240 km, and at each sampling site, the sampling points between the two habitat types (agricultural vs natural grassland soil) were approximately 200 m apart. At each GPS-located sampling site, five pseudo-replicate soil samples (50-200 mm depth) were recovered from within a 10 x 10 m quadrat. All pseudo-replicate samples from each habitat type (agricultural vs natural grassland soil) were bulked into a single sample. This process was repeated for all 15 sampling sites, resulting in a total of 30 soil samples (15 agricultural and 15 grassland soils). The samples were transported to the laboratory at 5 °C and stored at -20 °C for subsequent DNA extraction.
Analysis of the soil chemical parameters
Nine soil chemical parameters were analysed to evaluate their influence on the structure and composition of the soil bacterial and fungal communities. Soil pH was measured using a Crison pH meter (Crison Instruments, Barcelona, Spain) from the supernatants of 1:2.5 (g/g) soil/ deionised water suspensions, after soil particles had settled after 2 min of vigorous shaking.22 Total inorganic phosphate was measured from the supernatant of 4 g of soil in 39 mL of the Bray-1 solution after 1 min of vigorous shaking and filtration through Whatman paper.23,24 Extractable cations (K, Ca, Na, Mg) were measured from the supernatants of 1:10 (g/g) soil/ammonium acetate suspensions.24 Total carbon content was determined by oxidation with potassium dichromate and sulfuric acid as described previously.24 Total ammonium (NH4) and nitrate (NO3) contents were analysed by Bemlab (Pty) Ltd (Strand, Western Cape, South Africa) using standard protocols.
Genomic DNA extraction, amplification and high-throughput sequencing
Genomic DNA was extracted from 0.25 g soil (dry weight) using the PowerSoil DNA Isolation Kit (Mo Bio Laboratories Inc., Carlsbad, CA, USA) within 48 h of sampling, following the manufacturer's instructions. Soil samples were ground with an electric Powerlyser (Mo Bio Laboratories Inc.) before further processing. Aliquots of 5 of genomic DNA were premixed with GelRedTM Nucleic Acid Gel Stain (Biotium, Hayward, CA, USA), separated on 1% agarose gels and visualised under UV light to determine the success of the extraction.
DNA amplification was done at MRDNA (www.mrdnalab.com, Shallo-water, TX, USA) sequencing facility in a 30-cycle PCR, using the HotStarTaq Plus Master Mix Kit (Qiagen, Germantown, MD, USA). For bacteria, 16S rRNA gene V4 variable region PCR primers 515/80625 were used. For fungi, internal transcribed spacer region (ITS) primers ITS1f and ITS426 were used. The thermal cycling conditions for amplification of the two gene regions were as follows: 94 °C for 3 min, followed by 28 cycles of 94 °C for 30 s, 53 °C for 40 s and 72 °C for 1 min, with a final elongation step at 72 °C for 5 min. After amplification, PCR products were checked on 2% agarose gels to determine the success of amplification and the relative intensity of bands. Multiple samples were pooled together (e.g. 30 samples) in equal proportions based on their molecular weight and DNA concentrations. Pooled samples were purified using calibrated Ampure XP beads. Pooled and purified PCR products were used to prepare DNA libraries following the Illumina TruSeq DNA library preparation protocol. High throughput sequencing was performed at MRDNA on a MiSeq platform following the manufacturer's guidelines.
Sequence analysis
Sequences were processed using Qiime V1.9.1.27 Bacterial and fungal sequences were analysed independently. Firstly, the mapping file was checked to ensure that it is formatted correctly. Barcodes were extracted from the sequence reads. The split libraries command was used to demultiplex the fastq file and assign sequence reads to their respective sample according to their corresponding barcodes, using sample mapping information (Supplementary tables 2 and 3) at the default parameter implemented in Qiime. Chimeric sequences were screened and then removed using the USEARCH software28 and the Greengenes-derived 16S reference databa se implemented in Qiime29 for archaea and bacteria, respectively, and the UNITE-INSD (release 7) derived ITS reference database for fungi. High-quality reads were clustered into operational taxonomic units (OTUs) at 97% sequence similarity and assigned taxonomy using blast search against the Greengenes database (for archaea and bacteria) and the UNITE-INSD database (for fungi), respectively. Sequences identified as singletons were removed before sub-sampling to a depth of 3001 (for bacteria) and 12 358 (for fungi) sequences per sample to ensure equal sampling effort across samples.
Data access
The DNA sequences from this study were deposited in the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) database, with accession numbers SRR5341506 and SRR5341505 for bacterial 16S and fungal ITS, respectively.
Statistical analyses
All statistical analyses were conducted in R version 3.2.1.30 Differences in community composition were visualised using non-metric multidimensional scaling with Hellinger-transformed Bray-Curtis dissimilarities. Abiotic data were standardised (mean=0, SD=1) and the resulting data matrices were used to perform principal component analysis to evaluate differences in soil chemistry between agricultural soils and natural grassland soils. An analysis of similarity (ANOSIM) was performed to determine whether differences between habitats (agricultural vs natural grassland soils) were statistically significant (999 permutations, α<0.05). The number of shared OTUs between communities or samples was visualised using Venn diagrams. Differences in means for bacterial and fungal diversities metrics (species richness, Shannon-Wiener index, Simpson index and Pielou's evenness), phyla abundance and abiotic data were compared using paired two-tailed Student's t-tests. The compositions of major bacterial and fungal phyla were visualised using a heatmap with unweighted pair group method with arithmetic mean (UPGMA) clustering on Bray-Curtis distances after Hellinger transformation. Distance-based redundancy analysis was used to evaluate the effects of the environment on microbial community composition.31
Results and discussion
We evaluated and compared the compositions of the bacterial and fungal communities in agricultural and natural grassland soils from 15 sites in the Mpumalanga Province in the Grassland Biome of South Africa using deep amplicon sequencing. We also evaluated the effect of edaphic factors on the patterns of the bacterial and fungal communities in the two habitats.
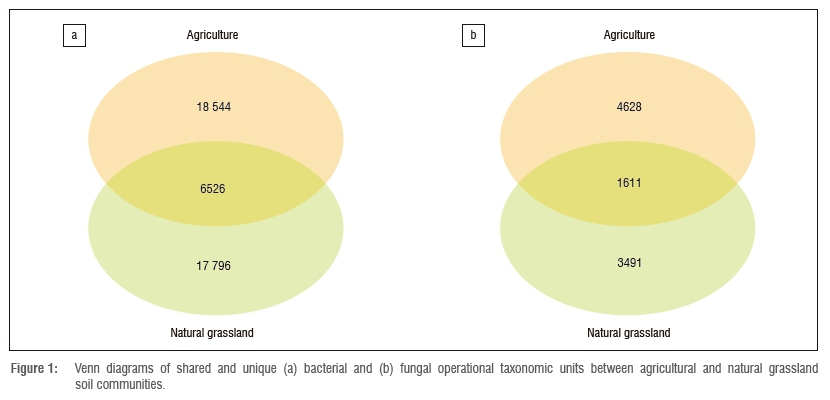
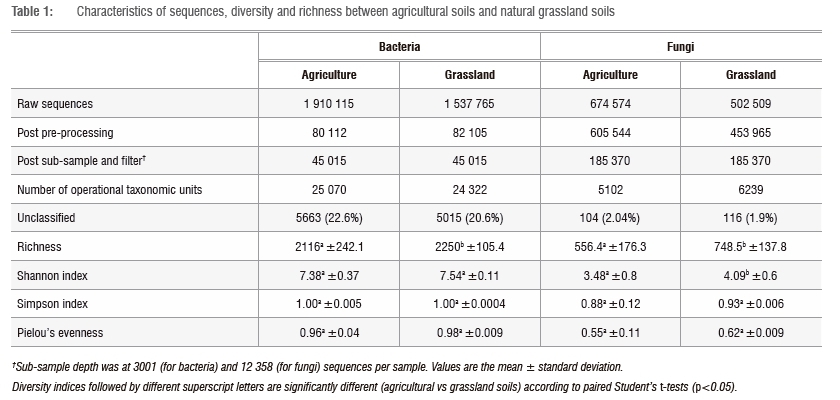
After quality filtering, removal of chimeras and singletons and sub-sampling, 90 030 bacterial and 370 740 fungal sequences remained, clustering into 42 866 bacterial and 9730 fungal OTUs (Figure 1, Table 1), respectively. Although the relative numbers of bacterial versus fungal taxa cannot be quantified from our data, our results suggest that natural grassland soils are richer in bacterial taxa than in fungal taxa, which is in agreement with previous studies showing that soil microbial biomass in natural grasslands is usually strongly dominated by bacteria.32,33
Bacterial alpha diversity metrics (Shannon and Simpson diversity indices and Pielou's evenness) revealed no significant difference (paired f-test, p>0.05) between agricultural and natural grassland soils (Table 1). However, natural grassland soils were significantly richer (paired f-test, p<0.05) in bacterial OTUs than were agricultural soils (Table 1), confirming the effects of land-use perturbation in decreasing soil microbial diversity.34,35 However, this finding contradicts previous studies on grassland soils, in which higher bacterial diversity was associated with land-use changes.11,36,37 The high level of bacterial genotypic richness detected in natural grassland may be explained by the influence of the high diversity of plant species in the grassland ecosystem, compared to the monoculture of agricultural crops.8,38 Plant species differ in the quantity and variety of their root exudates, as well as in the range of organic compounds in plant litter detritus: both factors are potential drivers of the higher microbial diversity often associated with more diverse plant communities.34,39-41 Our results are therefore consistent with the view that plant diversity promotes soil microbial diversity in grassland ecosystems.3
Fungal alpha diversity metrics (species richness, Shannon and Simpson diversity indices, and Pielou's evenness) were significantly higher (paired f-test, p<0.05) in natural grassland soils than in agricultural soils (Figure 1, Table 1), which is also consistent with previous studies on grasslands.11,36,37 The relatively low fungal diversity detected in agricultural soils is probably as a result of land-use management processes such as tillage. Tillage has been shown to damage fungal mycelia, leading to a reduction in fungal abundance and diversity in the soil.42,43 However, these results should be treated with caution as rarefaction curves obtained for both bacteria and fungi (Supplementary figures 1 and 2) did not plateau, indicating that the diversity of bacteria and fungi in both agricultural and natural grassland soils had not been fully sampled.
Differences in microbial diversity in managed agricultural soils, as compared to natural grassland soils, have been disputed in several studies. Some studies reported high bacterial diversity in conventionally managed arable soils as well as grassland soils.32,33,36,37 Others have shown that extensively managed grassland soils are richer in fungi than bacteria.37,44
The contrasting pattern of bacterial and fungal diversity in agricultural compared to natural grassland soils has been explained by the difference in nutrient availability in the two ecosystems.45,46 In extensively managed agricultural soils, high fertility and nutrient availability derived from the input of fertiliser favour bacterial communities, while the low soil fertility characteristics of natural grassland soils favour the fungal communities.45,46In comparison, the factors that shape the differences in bacterial and fungal diversity in managed soils are still largely unknown. The bacterial and fungal taxa unique to agricultural soils and natural grassland soils accounted for the majority of the raw sequences (Figure 1, Table 1). In addition, more than 20% of the bacterial sequences and about 2% of the fungal sequences could not be classified (Table 1), suggesting that these soils remain understudied.
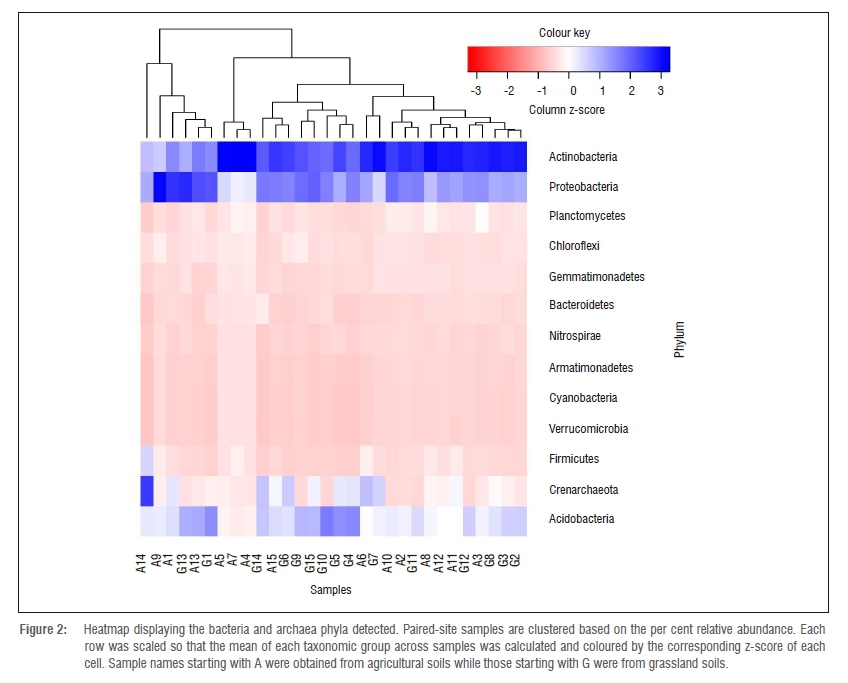
A total of 13 prokaryotic phyla (12 bacterial phyla and 1 archaeal phylum) were detected in the two habitats (Figure 2), of which Actinobacteria, Proteobacteria and Acidobacteria were the most abundant (Supplementary table 4). These three bacterial phyla are the most dominant taxa found in soils worldwide and make up the large proportion of bacterial 16S rRNA sequences available in public databases.47,48 Seven fungal phyla were detected in the two habitats (Figure 3), of which Ascomycota accounted for the majority of the sequences (71.6% and 63.4%) in agricultural and natural grassland soil samples, respectively (Supplementary table 4). Both the bacterial and fungal phyla identified are typically ubiquitous in soils and are thought to be important role players in soil geochemical cycling processes.3,14
Species of the phylum Acidobacteria were significantly more abundant (paired f-test, p<0.05) in natural grassland soils than in agricultural soils (Figure 2, Supplementary table 4). Acidobacteria abundance is commonly reduced in soils under conventional tillage11,49, which may explain this pattern. This phylum includes many environmentally important species, which provide a wide range of functions including the biodegradation of cellulose, hemicelluloses and chitin, nitrate and nitrite reduction, and the production of antimicrobial compounds.50
The most abundant bacterial and fungal taxa accounted for at least 10% of the total number of sequences in both agricultural and natural grassland soil samples. Analysis of these sequences at class, order, family and genus levels is shown in Supplementary table 4. At the class and order levels, Actinobacteria were significantly more abundant (paired f-test, p<0.05) in agricultural soils than in natural grassland soils (Supplementary table 4). These results suggest that land-use changes favour Actinobacteria abundance, which contradicts the results of previous related studies on non-African grassland soils, which suggested that Actinobacteria are more abundant in non-disturbed grassland soils than in agricultural managed soils.11,12 Actinobacteria are major producers of extracellular enzymes and secondary metabolites, and are thought to play significant roles in carbon cycling, plant disease suppression and enhancement of plant growth.51,52
At family level, Nectriaceae (Hypocreales) was the most abundant fungal family with 17.4% and 8.7% of all the fungal raw sequences in agricultural soils and natural grassland soils, respectively (Supplementary table 4). However, species of the family were significantly more abundant (paired f-test, p<0.05) in agricultural soils than in natural grassland soils (Supplementary table 4). The family Nectriaceae includes species of both economic and health importance. Species of the genus Fusarium produce mycotoxin and are both crop and opportunistic human pathogens.53,54
Non-metric multidimensional scaling analysis showed that bacterial and fungal communities in agricultural soils were distinct from those of the natural grassland soils (Supplementary figures 3 and 4). These results were confirmed by an analysis of similarity (ANOSIM: Rbacteria=0.46, p=0.001; Rfungi=0.16, p=0.001). A principal component analysis showed a clear discrimination of soil chemistry between agricultural soils and natural grassland soils. Soil pH and Ca concentrations tended to be higher in natural grassland soils, while NH4, NO3, K, C, P and Na concentrations were higher in agricultural soils (Supplementary figure 5). However, only three of the chemical parameters evaluated (Ca, P pH) were significantly different (paired f-test, p<0.05) between the two habitats (Supplementary table 5). The high concentration of phosphorus observed in agricultural soils may be explained by mineral fertilisation commonly used in agriculture practices to improve plant performance.55,56 Members of the soil microbial community, particularly bacteria, are capable of solubilising soil phosphate minerals into a usable form for plant uptake, which may influence the mineral concentration in the soil.57 The differences observed in the composition and pattern of the bacterial and fungal communities (according to ANOSIM analyses) in agricultural soils compared with those in natural grassland soils, can potentially be explained by the differences in the soil chemistry of the two habitats. In previous studies on grassland soils, soil fertilisation and land-use intensification have both been linked to shifts in richness and diversity of soil microbial communities.55,56
Using distance-based redundancy analysis, we found that the composition and pattern of the bacterial communities in agricultural soils compared to natural grassland soils were influenced significantly by both soil pH (R=0.008, p=0.002) and phosphorus concentrations (R=0.013, p=0.002) (Supplementary figure 6). Our results are in agreement with previous studies which have shown that soil pH and nutrient availability shaped soil bacterial diversity.2,9,13-15 In contrast, the differences found in the composition and pattern of the fungal communities between the two habitats were better explained by calcium (R=0.019, p=0.01) and phosphorus (R=0.01, p=0.004) concentrations (Supplementary figure 7), and corroborated previous studies.9,58
In summary, our results have shown that bacterial and fungal communities were significantly influenced by land-use changes, with agricultural soils containing distinct bacterial and fungal communities compared with natural grassland soils. Natural grassland soils consistently exhibited higher numbers of OTUs than did agricultural soils. The differences in microbial communities between the two habitat types were influenced by differences in soil chemical properties, supporting the concept that both soil chemical properties and microbial community compositions and diversities are altered after a conversion from a natural to an agricultural ecosystem.
This study was focused on a single economically important biome and represents the first demonstration of the effect of land-use changes on the diversity and structure of the soil microbial communities in African grassland soils. The study therefore serves as the benchmark for future studies on South African soil microbial diversity, and for monitoring future changes in soil microbial communities resulting from changing land use and climate.
Acknowledgements
We thank Dr Surendra Vikram and Mr Riegardt Johnson for providing bioinformatics support.
Authors' contributions
All authors conceived and designed the study and approved the final version; G.K.N. and A.V. collected the samples; G.K.N. performed research and analysed the data; G.K.N. wrote the manuscript; and D.A.C. and A.V. critically revised the manuscript.
References
1. Miransari M. Soil microbes and the availability of soil nutrients. Acta Physiol Plant. 2013;35:3075-3084. https://doi.org/10.1007/s11738-013-1338-2 [ Links ]
2. Fierer N, Jackson RB. The diversity and biogeography of soil bacterial communities. Proc Natl Acad Sci USA. 2006;103:626-631. https://doi.org/10.1073/pnas.0507535103 [ Links ]
3. Prober SM, Leff JW, Bates ST, Borer ET, Firn J, Harpole WS, et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol Lett. 2015;18:85-95. https://doi.org/10.1111/ele.12381 [ Links ]
4. Singh BK, Bardgett RD, Smith P Reay DS. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat Rev Microbiol. 2010;8:779-790. https://doi.org/10.1038/nrmicro2439 [ Links ]
5. Schenck zu Schweinsberg-Mickan M, Jörgensen RG, Müller T. Rhizodeposition: Its contribution to microbial growth and carbon and nitrogen turnover within the rhizosphere. J Plant Nutr Soil Sci. 2012;175:750-760. https://doi.org/10.1002/jpln.201100300 [ Links ]
6. Singh K, Trivedi P Singh G, Singh B, Patra DD. Effect of different leaf litters on carbon, nitrogen and microbial activities of sodic soils. Land Degrad Dev. 2016;27:1215-1226. https://doi.org/10.1002/ldr.2313 [ Links ]
7. Lehman RM, Cambardella CA, Stott DE, Acosta-Martinez V Manter DK, Buyer JS, et al. Understanding and enhancing soil biological health: The solution for reversing soil degradation. Sustainability. 2015;7:988-1027. https://doi.org/10.3390/su7010988 [ Links ]
8. Johnson MJ, Lee KY Scow KM. DNA fingerprinting reveals links among agricultural crops, soil properties, and the composition of soil microbial communities. Geoderma. 2003;114:279-303. https://doi.org/10.1016/S0016-7061(03)00045-4 [ Links ]
9. Lauber CL, Strickland MS, Bradford MA, Fierer N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol Biochem. 2008;40:2407-2415. https://doi.org/10.1016/j.soilbio.2008.05.021 [ Links ]
10. García-Orenes F, Morugán-Coronado A, Zornoza R, Scow K. Changes in soil microbial community structure influenced by agricultural management practices in a Mediterranean Agro-Ecosystem. PLoS ONE. 2013;8, e80522, 9 pages. https://doi.org/10.1371/journal.pone.0080522 [ Links ]
11. Lienhard P Terrat S, Prévost-Bouré NC, Nowak V Régnier T, Sayphoummie S, et al. Pyrosequencing evidences the impact of cropping on soil bacterial and fungal diversity in Laos tropical grassland. Agron Sustain Dev. 2014;34:525-533. https://doi.org/10.1007/s13593-013-0162-9 [ Links ]
12. Acosta-Martínez V Dowd S, Sun Y Allen V. Tag-pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol Biochem. 2008;40:2762-2770. https://doi.org/10.1016/j.soilbio.2008.07.022 [ Links ]
13. Lauber CL, Hamady M, Knight R, Fierer N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl Environ Microbiol. 2009;75:5111-5120. https://doi.org/10.1128/AEM.00335-09 [ Links ]
14. Nacke H, Thürmer A, Wollherr A, Will C, Hodac L, Herold N, et al. Pyrosequencing-based assessment of bacterial community structure along different management types in German Forest and grassland soils. PLoS ONE. 2011;6, e17000, 12 pages. https://doi.org/10.1371/journal.pone.0017000 [ Links ]
15. Kaiser K, Wemheuer B, Korolkow V Wemheuer F, Nacke H, Schöning I, et al. Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci Rep. 2016;6:33696. https://doi.org/10.1038/srep33696 [ Links ]
16. Geremia RA, Puscas M, Zinger L, Bonneville JM, Choler P. Contrasting microbial biogeographical patterns between anthropogenic subalpine grasslands and natural alpine grasslands. New Phytol. 2016;209:1196-1207. https://doi.org/10.1111/nph.13690 [ Links ]
17. Rutherford MC, Mucina L, Powrie LW. Biomes and bioregions of southern Africa. Strelitzia. 2006;19:32-50. [ Links ]
18. Bredenkamp G, Granger JE, Van Rooyen N. Moist Sandy Highveld Grassland. In: Low AB, Robelo AG, editors. Vegetation of South Africa, Lesotho and Swaziland. Pretoria: Department of Environmental Affairs and Tourism, 1996. [ Links ]
19. Abstract of Agricultural Statistic. Pretoria: Directorate: Statistics and Economic Analysis of the Department of Agriculture, Forestry and Fisheries; 2015. [ Links ]
20. Ferrar AA, Lötter MC. Mpumalanga biodiversity conservation plan handbook. Nelspruit: Mpumalanga Tourism and Parks Agency; 2007. [ Links ]
21. O'Connor TG, Bredenkamp GJ. Grassland. In: Cowling RM, Richardson DM, Pierce SM, editors. Vegetation of South Africa. Cambridge: Cambridge University Press; 1997. [ Links ]
22. Schofield RK, Taylor AW. The measurement of soil pH. Soil Sci Soc Am J. 1955;19:164-167. https://doi.org/10.2136/sssaj1955.03615995001900020013x [ Links ]
23. Bray RH, Kurtz LT. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945;59:39-45. https://doi.org/10.1097/00010694-194501000-00006 [ Links ]
24. Pansu M, Gautheyrou J. Handbook of soil analysis - Mineralogical, organic and inorganic methods. Berlin: Springer-Verlag; 2006. https://doi.org/10.1007/978-3-540-31211-6 [ Links ]
25. Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc Natl Acad Sci USA. 2011;108:4516-4522. https://doi.org/10.1073/pnas.1000080107 [ Links ]
26. White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninisky JJ, White TJ, editors. PCR protocols: A guide to methods and applications. New York: Academic Press; 1990. p. 315-322. https://doi.org/10.1016/B978-0-12-372180-8.50042-1 [ Links ]
27. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335-336. https://doi.org/10.1038/nmeth.f.303 [ Links ]
28. Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 2010;26:2460-2461. https://doi.org/10.1093/bioinformatics/btq461 [ Links ]
29. DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol. 2006;72:5069-5072. https://doi.org/10.1128/AEM.03006-05 [ Links ]
30. R Core Team. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2013. Available from: http://www.R-project.org [ Links ]
31. Ramette A. Multivariate analyses in microbial ecology. FEMS Microbiol Ecol. 2007;62:142-160. https://doi.org/10.1111/j.1574-6941.2007.00375.x [ Links ]
32. Hassink J, Bouwman LA, Zwart KB, Brussaard L. Relationships between habitable pore space, soil biota and mineralization rates in grassland soils. Soil Biol Biochem. 1993;25:47-55. https://doi.org/10.1016/0038-0717(93)90240-C [ Links ]
33. Velvis H. Evaluation of the selective respiratory inhibition method for measuring the ratio of fungal: Bacterial activity in acid agricultural soils. Biol Fert Soils. 1997;25:354-360. https://doi.org/10.1007/s003740050325 [ Links ]
34. Millard P Singh BK. Does grassland vegetation drive soil microbial diversity? Nutr Cycl Agroecosys. 2010;88:147-158. https://doi.org/10.1007/s10705-009-9314-3 [ Links ]
35. Mendes LW, Tsai SM, Navarrete AA, De Hollander M, Van Veen JA, Kuramae EE. Soil-borne microbiome: Linking diversity to function. Microbiol Ecol. 2015;70:255-265. https://doi.org/10.1007/s00248-014-0559-2 [ Links ]
36. Frey SD, Elliott ET, Paustian K. Bacterial and fungal abundance and biomass in conventional and no-tillage agroecosystems along two climatic gradients. Soil Biol Biochem. 1999;31:573-585. https://doi.org/10.1016/S0038-0717(98)00161-8 [ Links ]
37. De Vries FT, Hoffland E, Van Eekeren N, De Hollander M, Van Veen JA, Kuramae EE. Fungal/bacterial ratios in grasslands with contrasting nitrogen management. Soil Biol Biochem. 2006;38:2092-2103. https://doi.org/10.1016/j.soilbio.2006.01.008 [ Links ]
38. Osanai Y Bougoure DS, Hayden HL, Hovenden MJ. Co-occurring grass species differ in their associated microbial community composition in a temperate native grassland. Plant Soil. 2013;368:419-431. https://doi.org/10.1007/s11104-012-1529-4 [ Links ]
39. Kowalchuk GA, Buma DS, De Boer W, Klinkhamer PGL, Van Veen JA. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. A Van Leeuw J Microb. 2002;81:509-520. https://doi.org/10.1023/A:1020565523615 [ Links ]
40. Zak DR, Holmes WE, White DC, Peacock AD, Tilman D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology. 2003;84:2042-2050. https://doi.org/10.1890/02-0433 [ Links ]
41. Berg G, Smalla K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol. 2009;68:1-13. https://doi.org/10.1111/j.1574-6941.2009.00654.x [ Links ]
42. Balesdent J, Chenu C, Balabane M. Relationship of soil organic matter dynamics to physical protection and tillage. Soil Till Res. 2000;53:215-230. https://doi.org/10.1016/S0167-1987(99)00107-5 [ Links ]
43. Six J, Feller C, Denef K, Ogle SM, De Morae Sa JC, Albrecht A. Soil organic matter, biota and aggregation in temperate and tropical soils-effects of no-tillage. Agronomie. 2002;22:755-775. https://doi.org/10.1051/agro:2002043 [ Links ]
44. Bloem J, Schouten T, Didden W, Jagers op Akkerhuis G, Keidel H, Rutgers M, et al. Measuring soil biodiversity: Experiences, impediments and research needs. In: Francaviglia R, editor. Proceedings of the OECD expert meeting on soil erosion and soil biodiversity indicators; 2004 March 25-28; Rome, Italy. Paris: OECD; 2004. p. 109-129. [ Links ]
45. Bardgett RD, Frankland JC, Whittaker JB. The effects of agricultural management on the soil biota of some upland grasslands. Agr Ecosyst Environ. 1993;45:25-15. https://doi.org/10.1016/0167-8809(93)90057-V [ Links ]
46. Bardgett RD, Hobbs PJ, Frostegard A. Changes in soil fungal: Bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol Fert Soils. 1996;22:261-264. https://doi.org/10.1007/BF00382522 [ Links ]
47. Janssen PH. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl Environ Microbiol. 2006;72:1719-1728. https://doi.org/10.1128/AEM.72.3.1719-1728.2006 [ Links ]
48. Spain AM, Krumholz LR, Elshahed MS. Abundance, composition, diversity and novelty of soil Proteobacteria. ISME J. 2009;3:992-1000. https://doi.org/10.1038/ismej.2009.43 [ Links ]
49. Nemergut DR, Costello EK, Hamady M, Lozupone C, Jiang L, Schmidt SK, et al. Global patterns in the biogeography of bacterial taxa. Environ Microbiol. 2011;13:135-144. https://doi.org/10.1111/j.1462-2920.2010.02315.x [ Links ]
50. Ward NL, Challacombe JF, Janssen PH, Henrissat B, Coutinho PM, Wu M, et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl Environ Microb. 2009;75:2046-2056. https://doi.org/10.1128/AEM.02294-08 [ Links ]
51. Schlatter D, Fubuh A, Xiao K, Hernandez D, Hobbie S, Kinkel L. Resource amendments influence density and competitive phenotypes of Streptomyces in soil. Microbial Ecol. 2009;57:413-120. https://doi.org/10.1007/s00248-008-9433-4 [ Links ]
52. Dharumadurai D, Jiang Y Actinobacteria - Basics and biotechnological applications. Rijeka: InTech; 2016. [ Links ]
53. Marasas WFO, Nelson PE, Toussoun TA. Toxigenic Fusarium species: Identity and mycotoxicology. University Park, PA: Pennsylvania State University Press; 1984. [ Links ]
54. Rossman A, Seifert K. Phylogenetic revision of taxonomic concepts in the Hypocreales and other Ascomycota: A tribute to Gary J. Samuels. Stud Mycol. 2011;68:1-256. [ Links ]
55. Jangid K, Williams MA, Franzluebbers AJ, Sanderlin JS, Reeves JH, Jenkins MB, et al. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol Biochem. 2008;40:2843-2853. https://doi.org/10.1016/j.soilbio.2008.07.030 [ Links ]
56. Cassman NA, Leite MFA, Pan Y, De Hollander M, Van Veen JA, Kuramae EE. Plant and soil fungal but not soil bacterial communities are linked in long-term fertilized grassland. Sci Rep. 2016;6:23680. https://doi.org/10.1038/srep23680 [ Links ]
57. Rodriguez H, Fraga R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol Adv. 1999;17:319-339. https://doi.org/10.1016/S0734-9750(99)00014-2 [ Links ]
58. Pec GJ, Karst J, Taylor DL, Cigan PW, Erbilgin N, Cooke JEK, et al. Change in soil fungal community structure driven by a decline in ectomycorrhizal fungi following a mountain pine beetle (Dendroctonus ponderosae) outbreak. New Phytol. 2017;213:864-873. https://doi.org/10.1111/nph.14195 [ Links ]
 Correspondence:
Correspondence:
Gilbert Kamgan Nkuekam
Email: kamgan2000@yahoo.com
Received: 21 Aug. 2017
Revised: 21 Nov. 2017
Accepted: 16 Mar. 2018
Published: 30 May 2018
FUNDING: National Research Foundation (South Africa); University of Pretoria


{kind=link}
{kind=link}
{kind=link}
{kind=link}