










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.114 no.1-2 Pretoria ene./feb. 2018
http://dx.doi.org/10.17159/sajs.2018/20170266
RESEARCH ARTICLE
Palaeoecology of giraffe tracks in Late Pleistocene aeolianites on the Cape south coast
Charles HelmI, II; Hayley CawthraII, III; Richard CowlingII; Jan De VynckII; Curtis MareanII, IV; Richard McCreaI; Renee RustV
IPeace Region Palaeontology Research Centre, Tumbler Ridge, British Columbia, Canada
IICentre for Coastal Palaeoscience, Nelson Mandela University, Port Elizabeth, South Africa
IIIGeophysics Competency, Council for Geoscience, Bellville, South Africa
IVInstitute of Human Origins, School of Human Evolution and Social Change, Arizona State University, Tempe, Arizona, USA
VEvolutionary Studies Institute, Archaeology and Environmental Studies, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Until now there have been no reliable historical or skeletal fossil records for the giraffe (Giraffa camelopardalis) south of the Orange River or northern Namaqualand. The recent discovery of fossil giraffe tracks in coastal aeolianites east of Still Bay, South Africa, significantly increases the geographical range for this species, and has implications for Late Pleistocene climate and vegetation in the southern Cape. Giraffe populations have specialised needs, and require a savanna ecosystem. Marine geophysical and geological evidence suggests that the broad, currently submerged floodplains of the Gouritz and Breede Rivers likely supported a productive savanna of Vachellia karroo during Pleistocene glacial conditions, which would have provided a suitable habitat for this species. We show evidence for the hypothesis that the opening of the submerged shelf during glacial periods acted as a pathway for mammals to migrate along the southern coastal plain.
SIGNIFICANCE:
• The identification of fossil giraffe tracks on the Cape south coast, far from the area in which giraffe have previously been known to occur, is unexpected; conclusions about prehistoric conditions and vegetation can be drawn from this discovery.
Keywords: fossil tracks; Still Bay; savanna; South Africa
Introduction
Pleistocene aeolianites, the lithified remains of ancient dune systems, extend intermittently along much of the South African coastline, where the conditions were favourable for the deposition and subsequent preservation of these features. They are sensitive barometers of fluctuations in palaeoenvironments, as archived in their orientation, geometry, palaeontology and archaeological content.1 Exposed bedding planes characterise these sequences, and may contain ichnofossils.2,3 Aeolianites outcrop along much of the Cape south coast. In this area, a broad continental shelf is currently submerged but various authors have postulated the role of this shelf and its link to submerged landscapes4-6 - the opening of a landscape over which mammals could migrate without the impact of the Cape Fold Belt7, supporting populations that were hunted by ancient people and carnivores and fossilised in caves and rock shelters8,9.
A ground survey by the lead author (C.H.) between 2007 and 2016 of a 275-km stretch of coastline along the Cape south coast, from Witsand in the west to Robberg in the east (Figure 1), identified over a hundred Late Pleistocene trackway sites. These sites provide a rich source of palaeoecological information independent of the fossil remains that traditionally form the main data set for palaeoecological reconstruction. Fossil remains in caves and rock shelters are normally the remains of hunted prey and thus represent a biased sub-set of the animals on the landscape. In contrast, trackways have no such predation selection but rather are a direct reflection of the animals' locomotion and their frequency of walking across ancient land surfaces, although there is often a preservational bias towards larger, heavier trackmakers, which tend to leave deeper tracks. The bones of larger mammals are less likely to be transported to caves and rock shelters, so body fossil sites and track sites provide complementary sources of palaeoecological information. While many such track sites in the southern Cape occur in situ, others have been eroded out and lie as loose blocks at the base of cliffs. Some tracks can be attributed to extinct species or subspecies, while others indicate range differences compared with historical records or the skeletal fossil record. The age of the aeolianites can be obtained with optically stimulated luminescence (OSL) dating, which now allows the trackways to be much more accurately dated and placed on a geological timescale.
The giraffe track site described here occurs east of Still Bay, within a rare zone of concentration of fossil track sites. Some 120 m to the east of the site there is a significant accumulation of elephant tracks, which beyond this point remain evident in profile and as casts in a 300 m laterally persistent, low-angled, cross-laminated facies. Roberts et al.2 provided an analysis of these elephant and associated tracks, including geochronological control by the application of OSL and amino-acid racemisation dating, confirming their Late Pleistocene age. Some 300 m to the west of the giraffe track site lies a large displaced block, which contains probable tracks of four species of the Late Pleistocene megafauna. Numerous other fossil tracks have been identified between these sites.
Geological context
The ~800-km Cape south coast represents a tract of relatively low-relief coastal plain bordered at its landward geographic limit by the Cape Fold Belt and at the seaward margin by the Indian Ocean. The basement geology in the area investigated consists of Ordovician to Silurian quartzites of the Table Mountain Group and Devonian age Bokkeveld Group shales with interbedded sandstone packages.10
The south coast offshore area is distinguished by its wide, planed continental shelf forming the Agulhas Bank (~270 km wide south of Cape Agulhas) with a shelf break shoaling from a depth of -200 m off Cape Agulhas to -140 m south of Port Elizabeth.11 In general, the south coast is characterised by a wide, flat outer shelf, a middle shelf rise at a water depth of about 60-80 m, a relatively flat inner shelf and a narrow (-2-14 km), sediment-free rocky nearshore zone.12 A series of bathymetric nick points and erosional terraces across the shelf at 105-100 m, 80-75 m, 55-50 m and -40 m water depths are evidence of previous sea level stillstands.6,11
The rapid and globally extensive changes of Quaternary climates resulted in glacio-eustatic sea-level oscillations with maximum regressions exposing the entire Agulhas Bank. The Holocene sediment wedge is a quasi-continuous unconsolidated sediment feature stretching approximately 1500 km from the east to the south coast along the inner to mid-shelf.11 The major source of terrigenous sediment supply is by fluvial discharge from major rivers, which would have been substantial contributors to the sediments blanketing this ancient plain.
The Late Pleistocene coastal trackways, including the giraffe track site, occur in the Waenhuiskrans Formation of the Bredasdorp Group.13 Numerous subsequent geochronological investigations have been performed in rocks of the Waenhuiskrans Formation through OSL, thermally transferred OSL and amino-acid racemisation dating.2,1416 The great majority of these are from MIS (Marine Isotope Stage) 6 to MIS 5b, with limited MIS 11 deposits dated at ~400 ka.17 The dating studies performed at the elephant track sites and palaeosols east of the giraffe site yielded an age range from MIS 6 to MIS 5b (140±8.3 ka to 91±4.6 ka).2
Track preservation may have been promoted by the favourable substrate constituted by moist sand, swift burial as a result of high sedimentation rates, and the high bioclastic carbonate content of coastal dunes and inter-dune areas.18 In the region of the giraffe track site, aeolianite rocks form cliffs up to 50 m in height along a rugged 6 km stretch of coastline. Storm surges and high spring tides cause cliff collapse, which results in a high rate of removal of exposed tracks, with the likelihood of new trackways being exposed.
Methods
Nine visits were conducted to the coastline east of Still Bay by the first author (C.H.) between 2007 and 2011. A further visit was conducted in 2016 in order to assess the fate of the main elephant site first described by Roberts et al.2 During this visit, the giraffe track site was identified. Global Positioning System (GPS) readings were obtained for the track site using a handheld GPS device. Measurements and photographs were taken of the track-bearing surface and the better-preserved tracks. Photogrammetry was performed using a Canon PowerShot D30 camera. Point clouds and digital terrain models were compiled using Agisoft Photoscan Professional (v.1.0.4) and colour topographic profiles were created with CloudCompare (v.2.6.3.beta).
Geological outcrops were investigated in the field by means of correlation to known and documented deposits for which stratigraphic links could be demonstrated. At the site described here, standard field techniques were applied in understanding the context of the ichonofossils. Vertical logs were compiled from the coastal cliffs, the thicknesses of beds and foresets were measured, dip and strike readings were taken on strata to determine palaeowind direction, and geological samples were obtained for thin sections and petrography (Figure 2).
Results
The giraffe track site was not noted during any of the visits between 2007 and 2011, but was readily identified during the 2016 visit. Locality information for this site is reposited at the African Centre for Coastal Palaeoscience. The exposed ichnofossil surface is nestled between a prominent cliff to the north and the Indian Ocean to the south, on a wave-cut platform (Figure 3). The track-bearing surface cannot be extended through excavation.
The tracks are aligned in an approximate north-south, downslope direction on an in-situ surface that measures 3.9 m from north to south and 7.9 m from east to west. They are located in the basal geological unit of the sequence of deposits on the coastal cliffs, which are dominated by calcarenite-composition aeolianite. Variable cementation of the aeolianites has resulted in positive weathering of select surfaces and the layer containing the giraffe ichnofossils is competent. Beds are shallowly dipping (10-15°) and were laid down by southwesterly winds, likely the prevailing conditions at the time. Based on correlations to the reported stratigraphy2, we suggest that these deposits date to MIS 5e as the published ages2 for this unit obtained less than 1 km to the east are 126±7.1 ka and 121±6.5 ka. Dating studies above and below the giraffe track horizon would allow this site to be placed with greater certainty within the southern African Pleistocene track record.
There is considerable encrustation and boring of the surface with a modern gastropod species, Nodilittorina africana knysnaensis, which inhabits the highest intertidal zone, the littorina.19 The littorina is partially submerged during high tides and is impacted by ocean wave action. The tracks are subject to degradation both by wave action and the lithophagic effects of the gastropods in establishing their domicile burrows (Domichnia) on the hardgrounds supplied by the consolidated Pleistocene aeolianites.
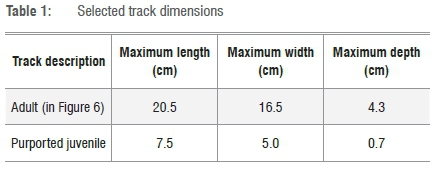
A total of 12 tracks are currently visible; the best preserved tracks are found at the southern end of the site, where the largest and the smallest tracks occur (Figures 4-6 and Table 1). Although individual trackways are probably present, they cannot be identified with certainty, and pace length and stride length cannot be confidently determined. Two tracks display partial over-printing. Minimal displacement rims are evident.
Identity of the trackmaker
The tracks appear morphologically consistent with those of the giraffe (Artiodactyla, Giraffidae, Giraffa camelopardalis). Other than giraffe, there are no extant mammals in southern Africa that leave tracks with these dimensions.20,21 Many of the coastal Late Pleistocene trackways that have been identified were made by bovids (Bovidae). In most cases these even-toed ungulate tracks cannot be identified to species level, and the terms 'large bovid', 'medium bovid' and 'small bovid' have to suffice. Exceptions include trackmakers with specialised hoof structure, e.g. the klipspringer (Oreotragus oreotragus) or adult tracks with distinctive size associated with differences in shape, such as the buffalo species Syncerus caffer and Syncerus antiquus. However, in comparing adult tracks of extant Giraffidae and Bovidae, the distinction is simpler, as giraffe tracks are uniquely large and long, relatively narrow compared with the largest bovids, and with a distinctive shape to the interdigital sulcus in well-preserved tracks. The better-preserved tracks at this site exhibit these characteristics. The presence of smaller tracks with similar morphology suggests that a family group may have made the tracks.
Sivatheres (short-necked giraffe) need to be excluded as potential trackmakers. We contend that sivatheres are unlikely on two grounds. Firstly, while the extinction date for Sivatherium maurusium, the African representative of the giant short-necked giraffes, cannot be determined with precision22, the most recent documented date of its occurrence in the western or southern Cape is 400 ka at Elandsfontein23,24. Possible Late Pleistocene sivathere teeth were recovered at Florisbad (north of Bloemfontein) in 1926, but have been lost.25
Secondly, while globally there appear to be no records of sivathere trackways, the massive sivatheres had very robust limb bones, along with substantially longer ossicones than extant giraffe. These factors, considered in the context of Lockley's observation that 'hooves follow horns'26, and Harris' comparative measurements of sivathere and giraffe22, suggest that sivathere tracks would not only be larger, but would also be relatively wider than the tracks of G. camelopardalis. The fact that the tracks east of Still Bay appear morphologically identical to those of G. camelopardalis supports the giraffe as the trackmaker.
Amongst the known Pleistocene megafauna in the southern Cape, the giant hartebeest (Megalotragus) merits consideration as a trackmaker. Although significantly larger than the extant red hartebeest {Alcelaphus buselaphus) and other alcelaphines, and although fossil tracks of this species have not been documented, the track morphology of this species would likely resemble that of other alcelaphines, and a track length of over 20 cm appears implausible (the track length of the red hartebeest has been recorded as 8.5-9 cm by Liebenberg20 and 8-10 cm by Stuart and Stuart21). The giant hartebeest is documented on the south coast in MIS 2 deposits at Nelson Bay Cave and MIS 5 and MIS 4 deposits at Klasies River.27
The final possibility is that the trackmaker was a hitherto undescribed member of the Pleistocene megafauna, with tracks that resemble those of the modern giraffe. While such a possibility cannot be completely excluded, it appears to be remote.
Discussion
Pliocene giraffid tracks recorded at Laetoli (3660 ka)28 had dimensions of 17 cm x 12.5 cm29. Holocene giraffe tracks, thought to be hundreds of years old, were recorded from the Kuiseb Delta in Namibia.30 We contend that the Still Bay trackways are the only Pleistocene giraffe tracks recorded thus far in southern Africa.
Historical records
Historically, giraffe in the Cape Province were mainly confined to 29°S (in the Orange River valley) and further north, associated with riverine woodland and arid savanna, respectively. There is no record of giraffe occurring south of 30°S. The southernmost record was at the Spoeg River, Namaqualand, as noted by Van Meerhoff, 28 November 1663.31 Here giraffe may have persisted in the Vachellia karroo woodlands of the Namaqualand river valleys as well as in isolated patches of V. erioloba on deep, sandy soils.32 Sclater33 noted that by 1761 'there does not seem to be any evidence of the occurrence of this animal south of the Orange River'. Mitchell and Skinner34 indicate that 'modem African giraffes were well known to the Khoisan as far south as Graaff Reinet', but provide no reference for this statement.
Skead35 noted that: '...Namaqualand in the north-west Cape is the most southerly district in which the giraffe has been known in historical times, and probably for long before that'. Skead35 also noted an 1837 report by Herschel of four giraffe near Cape Town36. However, this report appears to refer to giraffe in captivity.
Rock art records
A rock art site from the Hex River Valley includes compelling fine line images of at least two giraffe (Site no. Hx BF 3, slide numbered 13) in Buffelshoekkloof on the farm Kanetvlei, latitude 33°31' S. The images are yellow-orange in colour and were recorded as being 170 mm in height and in poor condition. Beside them antelope-shaped images were recorded, superimposed with finger dots. Figure 7 presents a tracing of these images.37
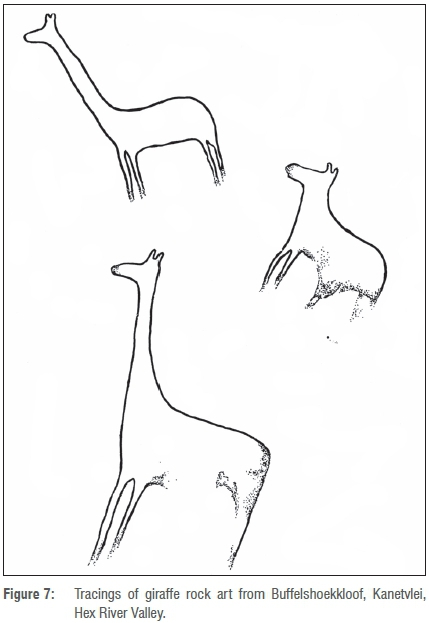
The rock art of the Hex River Valley includes entoptic images, finger dots and handprints, and the depiction of sheep (which are not known in the archaeological record of southern Africa before approximately 2.0-1.6 ka). It was probably executed in the final phase of the rock art tradition in the southwestern Cape by peoples who occupied the area within the last 2 ka. Evidence suggests that handprints, finger dots and strokes in the Western Cape are later in the dating sequence than fine line paintings.38 Paintings of handprints and finger dots are superimposed on antelope in the same shelter as the images of giraffe, suggesting that the inanimate images were executed later in the chronological sequence.
However, rock art images of giraffe cannot be regarded as a proxy for the historical distribution of the species, as they may simply reflect awareness by the artists that these distinctive mammals occurred further north. The Hex River Valley does provide access to areas further north from which historical records are known.
The only other possible San rock art of giraffe in the Cape has been recorded from the Eastern Cape near the Swart Kei River and near Whittlesea in the Queenstown District.39 It is also contended that these images may represent mountain reedbuck. This species, although relatively long-necked in comparison to most other ungulates, has forward-pointing horns which are distinct from the ossicones of the giraffe. The Hex River images appear consistent with the depiction of giraffe ossicones rather than reedbuck horns.
Fossil records
Skead35 noted 'signs that in prehistoric times a giraffe or giraffe-like animal lived as far south as Darling' (~33.3°S), mentioning a report by Cooke40 of fossil specimens from this area in the South African Museum in Cape Town. However, Skead35 noted further that Hendey, the palaeontologist at the museum, knew of no other fossils having been found at that site, and had cast doubt on the locality. Harris in his description of Giraffoidea attributed mid-Pleistocene age to this possible site.41 Harris described a sivathere species and a giraffine species in Pliocene deposits at Langebaanweg (~33.0°S).42
The Cape south coast has a diversity of fossil palaeoarchives that have samples of fossil animal remains dating from the Pleistocene and Holocene. These represent the predation and collection activities of primarily modern humans, hyenas, jackals, leopards and porcupines. Giraffe are regularly predated by humans with advanced projectile weapons and/or poison technologies, e.g. the Hadza.43 Giraffe are not regularly predated by hyenas, leopards and jackals, although hyenas and jackals will scavenge them when possible. It is important to note that the bones of very large mammals like giraffe are not as readily transported by predators as the bones of smaller prey, so there is a taphonomic filter that biases their presence in palaeoarchives such as caves and rock shelters.44
Giraffe fossils are not reported from palaeoarchives on the south coast, despite there being some very large samples collected by a diversity of bone-collecting agents. Several sites have sediments that date to the likely time of the giraffe track site. Pinnacle Point (PP) 13B and PP30 have MIS 6 sediments and giraffe are not reported.45 Blombos, PP13B and Klasies River have MIS 5 sediments and no reported giraffe remains.46,47 There is a large sample of fauna dating from MIS 2-1 at Nelson Bay Cave, and giraffe are not reported.48 It would seem that the giraffe track site provides a unique record, perhaps because giraffe were not predated or their remains were not regularly transported to these caves. This finding highlights the fact that ichnofossils can provide an independent record of past animal occurrences that expands on the records provided by traditional fossil archives.
Past distribution of giraffe in KwaZulu-Natal
Cramer and Mazel49 found no reliable historical records of giraffe in KwaZulu-Natal, nor any rock art of giraffe. However, they noted reports of giraffe bones at three archaeological sites, the southernmost of which was Sibudu Cave, latitude 29°31'S. The most recent of these sites was dated at 1 ka.
Implications of the palaeoecology of the Palaeo-Agulhas Plain
Recent reviews of the palaeoecology of the Palaeo-Agulhas Plain can be found in Marean et al.8 and Copeland et al.9 The faunal composition of the Palaeo-Agulhas Plain was first revealed by publications on the fossil faunal remains from Nelson Bay Cave and Klasies River.46,48 More recent publications of faunal assemblages from Blombos47 and Pinnacle Point45 have added to this body of data. It is important to note that these assemblages were accumulated primarily by humans, and modern hunter-gatherers sample with a ~10 km daily foraging radius50, so the assemblages represent both interior and Palaeo-Agulhas Plain faunas. The Pleistocene samples all include both browsing and grazing species, with the grazers being more abundant, and the browsing species being typical of the Cape fauna today such as grysbok, grey duiker and bushbuck. The species absent from the Cape today and present in these Pleistocene assemblages include large-bodied grazers typical of African savanna ecosystems. The large-bodied grazing faunal suite includes many extinct species. Marean51 hypothesised that when sea levels retreated, this exposed plain was the location of a seasonal migration ecosystem with animals wintering in the west to exploit green forage in the winter rainfall regime, and then moving east in the summer to forage the green summer rainfall grasses. A strontium isotope analysis showed that these grazing species lived almost exclusively on the Palaeo-Agulhas plain and did not venture inland in any significant manner.9 An analysis of age at death data of fossil blue antelope is consistent with a seasonal migratory movement.52
The emergence of the Palaeo-Agulhas Plain is strictly controlled by sea level and the position of resultant shorelines.5,53 Interpretation of marine geophysical data in the Mossel Bay area suggests a low-relief 'plains' landscape offshore of the southern coastal plain16,6, and this region was dominated by shallowly incised rivers with vast floodplains and wetlands. Cemented seafloor deposits, as well as a mobile wedge of unconsolidated sediment available for dune construction through times of sea-level regression, provide evidence that the aeolianite deposits on the present day coastal plain are the fringing remnants of a dominance of comparable deposits on the adjacent shelf.16
Contemporary giraffe populations in southern Africa are invariably associated with subtropical savanna, mostly dominated by deciduous acacias (Vachellia spp.) with a field layer of summer-growing, C4 grasses.35 Their presence in the southern Cape fossil record, far south of their current range, has important implications for the Late Pleistocene climate and vegetation of the region. Most noteworthy is the requirement for extensive subtropical savanna, a habitat that is rare in the present landscape, being confined to the alluvial soils of the incised valleys of the Gouritz, Breede, Great Brak and Little Brak Rivers. This is the 'doringveld' of Vlok et al.54, comprising an open and species-poor stratum of relatively low (3-5 m) trees, dominated by deciduous V. karroo, and a field layer of largely ephemeral grasses and karroid shrubs. Although several introductions of this species to this habitat have occurred in the recent past, it appears unlikely that this habitat can sustainably support populations of giraffe. However, when complemented with evergreen thicket, a habitat that would likely have been widespread on the calcareous substrata that dominated the Palaeo-Agulhas Plain, the sustenance of a giraffe population appears more realistic.
The giraffe populations of the Late Pleistocene southern Cape likely occurred on the wide, alluvial valleys of the aforementioned rivers, which meandered across the low-gradient Palaeo-Agulhas Plain6, when it was exposed during glacial conditions52. While appropriate edaphic conditions, namely fertile alluvial soils55, certainly existed to support giraffe habitat at this time, there is less certainty regarding the climate. Isotopic evidence derived from speleothems from Pinnacle Point shows shifts from C3 vegetation and winter rain to C4 grasses and summer rain at several times during the Middle and Late Pleistocene, notably at the transition between MIS 5 and MIS 4 (about 74 ka).56,57 The invasion of the usually temperate Southern Cape by subtropical biomes (C4 grassland and savanna) during these transitional intervals would have provided habitat suitable for giraffe and many other ungulate species observed in the Pleistocene fossil record, including several species now extinct or occurring many hundreds of kilometres to the east and north.
Conservation
The likelihood is that the track surface became exposed at some point since 2011, as it would likely have been noted during searches for track sites during visits between 2007 and 2011. Visits in 2017 revealed interim partial degradation of the track surface caused by erosion. Given its precarious perch, the likelihood of it falling into the ocean is high. The slab is probably too heavy for helicopter retrieval to be considered. Preserving it with a substance such as ParaloidTM would help prevent erosion by weather and gastropods on the surface, but would not prevent its collapse into the Indian Ocean. The photogrammetry that has been performed allows for the reproduction of the track surface. Alternatively, the creation of a replica using latex or silicone would be feasible. The nearby museum in Still Bay would potentially form a suitable repository for such specimens, and for their interpretation. The richness of the ichnofauna along the coastline east of Still Bay, and the ephemeral nature of exposed track sites, makes the need for regular ichnological surveys desirable.
Conclusions
The discovery of a Pleistocene giraffe track site on the Cape south coast is unexpected, given the lack of known skeletal material in the region and the recorded historical distribution of the species. Our description of the earliest known evidence for giraffes in this region has significant palaeoecological implications because of the specialised feeding niche of giraffes which require a savanna ecosystem. The likely location for this ecosystem is on the broad floodplain of the southern Cape river valleys which could have supported a savanna of Vachellia karroo and C4 grasses. The presence of giraffe tracks at the margin of this submerged plain is likely evidence for such vegetation.
Acknowledgements
The assistance and support of Stephen Townley Bassett, Chris Heese, Linda Helm and Thalassa Matthews are acknowledged. Guy Gardner, and Peter Todd and family, generously provided access across their land to the coastline. We thank Dr John Rowan and Dr Jan Venter for their helpful review and comments, and Izak Venter and Tertius Haak for field assistance. Dr David Roberts made the initial elephant trackway discoveries; his mentorship and presence are sorely missed.
Authors' contributions
C.H. is the senior and corresponding author and co-discoverer of the site; he was responsible for the conceptualisation, site analysis, data collection, data analysis, write-up of the manuscript and project leadership. H.C. was responsible for the data analysis and field stratigraphy, contributed regarding the geological context, and reviewed drafts and revisions. R.C. was responsible for the palaeobotany contributions; contributed to the conceptualisation and data analysis, and reviewed drafts and revisions. J.D.V. is the co-discoverer of the site and was responsible for site analysis, data collection and photography for photogrammetry, and reviewed drafts and revisions. C.M. was responsible for the palaeoecology, contributed to the methodology, and reviewed drafts and revisions. R.M. contributed to the data analysis and photogrammetry, and reviewed drafts and revisions. R.R. was responsible for the rock art contributions and reviewed drafts and revisions.
References
1. Roberts DL, Cawthra HC, Musekiwa C. Dynamics of Late Cenozoic aeolian deposition along the South African coast: A record of evolving climate and ecosystems. In: Martini IP Wanless HR, editors. Sedimentary coastal zones from high to low latitudes: Similarities and differences. Special Publication 388. London: Geological Society of London; 2014. p. 353-387. http://dx.doi.org/10.1144/SP388.11 [ Links ]
2. Roberts DL, Bateman MD, Murray-Wallace CV Carr AS, Holmes PJ. Last interglacial fossil elephant trackways dated by OSL/AAR in coastal aeolianites, Still Bay, South Africa. Palaeogeogr Palaeoclimatol Palaeoecol. 2008;257(3):261-279. http://dx.doi.org/10.1016/j.palaeo.2007.08.005 [ Links ]
3. Helm C, McCrea R, Helm D. A South African Pleistocene avian and mammal track site with purported prints of a shod hominid. The Digging Stick. 2012;29(3):17-20. [ Links ]
4. Dingle RV, Rogers J. Pleistocene palaeogeography of the Agulhas Bank. T Roy Soc S Afr.1972;40:155-465. http://dx.doi.org/10.1080/00359197209519415 [ Links ]
5. Van Andel TH. Late Pleistocene sea levels and human exploitation of the shore and shelf of southern South Africa. J Field Archaeol. 1989;16:133-155. http://dx.doi.org/10.1179/jfa.1989.16.2.133 [ Links ]
6. Cawthra HC, Compton JS, Fisher EC, MacHutchon MR, Marean CW. Submerged terrestrial landscape features off the South African south coast. In: Harff J, Bailey G, Lüth F, editors. Geology and archaeology: Submerged landscapes of the continental shelf. Special Publication 411. London: Geological Society of London; 2015. p. 219-233. http://dx.doi.org/10.1144/SP411.11 [ Links ]
7. Compton JS. Pleistocene sea level fluctuations and human evolution on the southern coastal plain of South Africa. Quat Sci Rev. 2011;30(5-6):506-527. http://dx.doi.org/10.1016/j.quascirev.2010.12.012 [ Links ]
8. Marean CW, Cawthra HC, Cowling RM, Esler KJ, Fisher E., Milewski A, et al. Stone Age people in a changing South African Greater Cape Floristic Region. In: Allsopp N, Colville JF, Verboom GA, editors. Fynbos: Ecology, evolution, and conservation of a megadiverse region. Oxford: Oxford University Press; 2014. p. 164-199. http://dx.doi.org/10.1093/acprof:oso/9780199679584.001.0001 [ Links ]
9. Copeland SR, Cawthra HC, Fisher EC, Lee-Thorp JA, Cowling RM, Le Roux PJ, et al. Strontium isotope investigation of ungulate movement patterns on the Pleistocene Paleo-Agulhas Plain of the Greater Cape Floristic Region, South Africa. Quat Sci Rev. 2016;141:65-84. http://dx.doi.org/10.1016/j.quascirev.2016.04.002 [ Links ]
10. Newton AR, Shone RW, Booth PWK. The Cape Fold Belt. In: Johnson MR, Annhauser CR, Thomas RJ, editors. The geology of South Africa. Johannesburg: Geological Society of South Africa/Council for Geoscience; 2006. p. 521-531. [ Links ]
11. Martin AK, Flemming BW. The Holocene shelf sediment wedge off the south and east coast of South Africa. In: Knight RJ, McLean JR, editors. Shelf sands and sandstones. Memoir II. Calgary: Canadian Society of Petroleum Geologists; 1986. p. 27-44. [ Links ]
12. Birch GF. Nearshore Quaternary sedimentation off the south coast of South Africa: (Cape Town to Port Elizabeth) - Geological Survey Bulletin 67. Pretoria: Government Printing Works; 1980. [ Links ]
13. Malan JA. Lithostratigraphy of the Waenhuiskrans Formation (Bredasdorp Group) - South African Committee for Stratigraphy Lithostratigraphic Series 8. Pretoria: Department of Mineral and Energy Affairs;1989. [ Links ]
14. Bateman MD, Carr AS, Dunajko AC, Holmes PJ, Roberts DL, McLaren SJ, et al. The evolution of coastal barrier systems: A case study of the Middle-Late Pleistocene Wilderness barriers, South Africa. Quat Sci Rev. 2011;30:63-81. http://dx.doi.org/10.1016/j.quascirev.2010.10.003 [ Links ]
15. Jacobs Z, Roberts RG, Lachlan TJ, Karkanas P Marean CW, Roberts DL. Development of the SAR TT-OSL procedure for dating Middle Pleistocene dune and shallow marine deposits along the southern Cape coast of South Africa. Quat Geochronol. 2011;5(5):491-513. http://dx.doi.org/10.1016/j.quageo.2011.04.003 [ Links ]
16. Cawthra HC, Bateman MD, Carr AS, Compton JS, Holmes PJ. Understanding Late Quaternary change at the land-ocean interface: A synthesis of the evolution of the Wilderness coastline, South Africa. Quat Sci Rev. 2014;99(1):210-223. https://doi.org/10.1016/j.quascirev.2014.06.029 [ Links ]
17. Roberts DL, Karkanas P Jacobs Z, Marean CW, Roberts RG. Melting ice sheets 400,000 yr ago raised sea level by 13 m: Past analogue for future trends. Earth Planet Sci Lett. 2012;357-358:226-237. http://dx.doi.org/10.1016/j.epsl.2012.09.006 [ Links ]
18. Roberts DL. Last Interglacial hominid and associated vertebrate fossil trackways in coastal eolianites, South Africa. Ichnos. 2008;15(3):190-207. http://dx.doi.org/10.1080/10420940802470482 [ Links ]
19. Branch GM, Griffiths CL, Branch ML, Beckley IE. Two oceans: A guide to the marine life of southern Africa. Cape Town: David Philip; 1994. [ Links ]
20. Liebenberg L. A photographic guide to tracks and tracking in southern Africa. Cape Town; Struik Publishers; 2000. [ Links ]
21. Stuart C, Stuart T. A field guide to the tracks and signs of southern and east African wildlife. Cape Town: Struik Nature; 2000. [ Links ]
22. Harris JM. Family Giraffidae. In: Harris JM, editor. Koobi Fora research project: Volume 3. The fossil ungulates: Geology, fossil artiodactyls, and palaeoenvironments. Oxford: Clarendon Press; 1991. p. 93-138. [ Links ]
23. Klein RG, Avery G, Cruz-Uribe K, Steele TE. The mammalian fauna associated with an archaic hominin skullcap and later Acheulean artifacts at Elandsfontein, Western Cape Province, South Africa. J Hum Evol. 2007;52:164-186. http://dx.doi.org/10.1016/j.jhevol.2006.08.006 [ Links ]
24. Kingdon J, Happold D, Butynski T, Hoffmann M, Happold M, Kalina J. Mammals of Africa. vol. VI. London: A & C Black Publishers Ltd; 2013. [ Links ]
25. Singer R, Boné EL. Modern giraffes and the fossil giraffids of Africa. Ann S Afr Mus.1960;45:375-603. [ Links ]
26. Lockley M. The eternal trail - A tracker looks at evolution. New York: Perseus Publishing; 1999. [ Links ]
27. Klein RG. Palaeoenvironmental implications of Quaternary large mammals in the Fynbos Region. In: Deacon HJ, Hendey QB, Lambrechts JJN, editors. Fynbos palaeoecology: A preliminary synthesis. Pretoria: Cooperative Scientific Programmes; 1983. p. 116-138. [ Links ]
28. Lockley M, Roberts G, Kim JY In the footprints of our ancestors: An overview of the hominid track record. Ichnos. 2008;15(3-4):106-125. http://dx.doi.org/10.1080/10420940802467835 [ Links ]
29. Masao FT, Ichumbaki EB, Cherin M, Barili A, Boschian G, Iurino DA, et al. New footprints from Laetoli (Tanzania) provide evidence for marked body size variation in early hominins. eLife. 2016;5, e19568, 29 pages. http://dx.doi.org/10.7554/eLife.19568 [ Links ]
30. Bennett MR, Liutkus CM, Thackeray F, Morse SA, McClymont J, Stratford D. Footprints of the Kuiseb Delta, Namibia. The Digging Stick. 2010;27:3. [ Links ]
31. Godée Molsbergen IE. Reizen in Zuid-Afrika in de Hollandse tijd. Deel 1.Tochten naar het Noorden, 1652-1686's [Travels in South Africa during the Dutch period. Part 1. Excursions to the North, 1652-1686]. Gravenhage: Martinus Nijhoff for Lichthoten Society; 1916 [Dutch]. In: Skead CJ. Historical incidence of the larger land mammals in the broader Western and Northern Cape. In: Boshoff AF, Kerley GIH, Lloyd PH, editors. Port Elizabeth: Centre for African Conservation Ecology, Nelson Mandela Metropolitan University; 2011. p. 288. [ Links ]
32. Cowling RM, Pierce SM. Namaqualand, a succulent desert. Cape Town: Fernwood Press; 1999. [ Links ]
33. Sclater WL. The mammals of South Africa. Volume I. London: Porter; 1900. [ Links ]
34. Mitchell G, Skinner JD. On the origin, evolution and phylogeny of giraffes Giraffa camelopardalis. Trans R Soc S Afr. 2003;58(10):51-73. http://dx.doi.org/10.1080/00359190309519935 [ Links ]
35. Skead CJ. Historical mammal incidence in the Cape Province. Vol. 1: The Western and Northern Cape. Cape Town: Department of Nature and Environmental Conservation of the Provincial Administration of the Cape of Good Hope; 1980. [ Links ]
36. Evans DS, Deeming TJ, Evans BH, Goldfarb S. Herschel at the Cape: Diaries and correspondence of Sir John Herschel, 1834-1838. Austin, TX: University of Texas Press; 1969. [ Links ]
37. Rust R. A micro-regional survey of rock art in the Hex River Valley, Western Cape, South Africa [BA honours thesis]. Stellenbosch: Stellenbosch University; 1995. [ Links ]
38. Yates RJ, Manhire AH, Parkington JE. Rock painting and history in the southwestern Cape. In: Dowson TA, Lewis-Williams JD, editors. Contested images: Diversity in southern African rock art research. Johannesburg: Witwatersrand University Press; 1994. p. 29-60. [ Links ]
39. Stow GW. The native races of South Africa. A history of the intrusion of the Hottentots and Bantu into the hunting grounds of the Bushmen, the Aborigines of the country. London: Swan Sonnenschein & Co. Limited; 1905. [ Links ]
40. Cooke HBS. Some fossil mammals in the South African Museum collections. Ann S Afr Mus. 1955;42:161-168. [ Links ]
41. Harris JM, Solounias N, Geraads D. Giraffoidea. In: Werdelin L, Sanders WJ, editors. Cenozoic mammals of Africa. Berkeley, CA: University of California Press; 2010. p. 797-811. http://dx.doi.org/10.1525/california/9780520257214.003.0039 [ Links ]
42. Harris JM. Pliocene Giraffoidea (Mammalia, Artiodactyla) from the Cape Province. Ann S Afr Mus. 1976:69;325-353. [ Links ]
43. Marlowe F. The Hadza: hunter-gatherers of Tanzania. Berkeley: University of California Press; 2010. [ Links ]
44. Monahan CM. The Hadza carcass transport debate revisited and its archaeological implications. J Archaeol Sci. 1998;25:405-424. https://doi.org/10.1006/jasc.1997.0241 [ Links ]
45. Rector AL, Reed KE. Middle and Late Pleistocene faunas of Pinnacle Point and their paleoecological implications. J Hum Evol. 2010;59(3-4):340-357. https://doi.org/10.1016/j.jhevol.2010.07.002 [ Links ]
46. Klein RG. The mammalian fauna of the Klasies River Mouth sites, southern Cape Province, South Africa. S Afr Archaeol Bull. 1976;31:75-98. https://doi.org/10.2307/3887730 [ Links ]
47. Henshilwood CS, Sealy JC, Yates RJ, Cruz-Uribe K, Goldberg P, Grine FE, et al. Blombos Cave, southern Cape, South Africa: Preliminary report on the 1992-1999 excavations of the Middle Stone Age levels. J Archaeol Sci. 2001;28:421-448. https://doi.org/10.1006/jasc.2000.0638 [ Links ]
48. Klein RG. The Late Quaternary mammalian fauna of Nelson Bay Cave (Cape Province, South Africa): Its implications for megafaunal extinctions and environmental and cultural change. Quatern Res. 1972;2:135-142. https://doi.org/10.1016/0033-5894(72)90034-8 [ Links ]
49. Cramer MD, Mazel AD. The past distribution of giraffe in Kwazulu-Natal. S Afr J Wildl Res. 2007;37:198-201. [ Links ]
50. Marlowe FW. Hunter-gatherers and human evolution. Evol Anthropol. 2005;14(2):54-67. https://doi.org/10.1002/evan.20046 [ Links ]
51. Marean CW. Pinnacle Point Cave 13B (Western Cape Province, South Africa) in context: The Cape Floral Kingdom, shellfish, and modern human origins. J Hum Evol. 2010;59:425-443. https://.org/10.1016/j.jhevol.2010.07.011 [ Links ]
52. Faith TJ, Thompson JC. Fossil evidence for seasonal calving and migration of extinct blue antelope (Hippotragus leucophaeus) in southern Africa. J Biogeogr. 2013;40(11):2108-2118. https://doi.org/10.1111/jbi.12154 [ Links ]
53. Fisher EC, Bar-Matthews M, Jerardino A, Marean CW. Middle and Late Pleistocene paleoscape modeling along the southern coast of South Africa. Quat Sci Rev. 2010;29:1382-1398. http://dx.doi.org/10.1016/j.quascirev.2010.01.015 [ Links ]
54. Vlok JHJ, Euston-Brown DIW, Cowling RM. Acocks' Valley Bushveld 50 years on: New perspectives on the delimitation, characterisation and origin of thicket vegetation. S Afr J Bot. 2003;69:27-51. http://dx.doi.org/10.1016/S0254-6299(15)30358-6 [ Links ]
55. Cawthra HC. The marine geology of Mossel Bay [PhD thesis]. Cape Town: University of Cape Town; 2014. [ Links ]
56. Bar-Matthews M, Marean CW, Jacobs Z, Karkanas IP Fisher EC, Herries AI, et al. A high resolution and continuous isotopic speleothem record of paleoclimate and paleoenvironment from 90 to 53 ka from Pinnacle Point on the south coast of South Africa. Quat Sci Rev. 2010;29(17):2131-2145. http://dx.doi.org/10.1016/j.quascirev.2010.05.009 [ Links ]
57. Braun K. Influence of the Agulhas Current on the terrestrial climate of South Africa as derived from speleothems [PhD thesis]. Jerusalem: Hebrew University of Jerusalem; 2014. [ Links ]
 Correspondence:
Correspondence:
Charles Helm
Email: helm.c.w@gmail.com
Received: 11 Aug. 2017
Revised: 30 Sep. 2017
Accepted: 18 Oct. 2017
FUNDING: None


{kind=link}
{kind=link}
{kind=link}