










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.112 n.9-10 Pretoria Sep./Oct. 2016
http://dx.doi.org/10.17159/sajs.2016/a0177
COMMENTARIES
http://dx.doi.org/10.17159/sajs.2016/a0177
Response to Thackeray (2016) - The possibility of lichen growth on bones of Homo naledi: Were they exposed to light?
Patrick S. Randolph-QuinneyI, II, III; Lucinda R. BackwellII; Lee R. BergerII; John HawksII, IV; Paul H.G.M. DirksII, V; Eric M. RobertsII, V; Godwin NhauroII; Jan KramersVI
ISchool of Forensic and Applied Sciences, University of Central Lancashire, Preston, Lancashire, United Kingdom
IIEvolutionary Studies Institute, University of the Witwatersrand, Johannesburg, South Africa
IIISchool of Anatomical Sciences, University of the Witwatersrand Medical School, Johannesburg, South Africa
IVDepartment of Anthropology, University of Wisconsin-Madison, Madison, Wisconsin, USA
VDepartment of Geoscience, James Cook University, Townsville, Queensland, Australia
VIDepartment of Geology, University of Johannesburg, Johannesburg, South Africa
Thackeray1 questions the hypothesis of deliberate body disposal in the Rising Star Cave by Homo naledi, as proposed by Dirks and colleagues2. Thackeray proposes that lichens produced mineral staining on the skeletal remains of H. naledi. As lichens require some exposure to light, in Thackeray's opinion, the presence of mineral staining necessitates either a direct entrance deep into the Rising Star Cave that once admitted light into the Dinaledi Chamber, or relocation of mineral-stained bones from a location exposed to light. Here we consider multiple lines of evidence that reject Thackeray's hypothesis that lichens deposited mineral staining upon the surface of these skeletal remains. We welcome the opportunity to address the inferences presented by Thackeray, and further hope that this response may dispel misinterpretations of our research2, and of other areas of the scientific literature that bear upon site formation processes at work within the Rising Star Cave system.
Briefly, we review Thackeray's1 chain of inference: (1) lichens can be found today growing as colonies on the surface of chert, dolomite and sub-aerially exposed bones found within the Cradle of Humankind; (2) some colonies of lichen on extant rocks overlie deposits of black manganese oxy-hydroxide on the rock surfaces; (3) some manganese (Mn) mineral deposits appear as diffuse spots, which have some resemblance to the shape and surface patterning of some lichen colonies; (4) small dots or spots of Mn present on the surface of the bones of H. naledi may, therefore, have been produced as a consequence of lichen growth; (5) the growth of lichen colonies requires light, either in the open environment or within the light zone of caves; (6) by extension, the growth of lichen on the bones of H. naledi required the presence of subdued, but essential lighting; (7) the presence of such lighting indicates the existence of a second entrance into the Dinaledi Chamber; therefore (8) a re-assessment of the deliberate body disposal hypothesis is required.
We must state from the outset that we have no disagreement with conditions (1) to (3) above, which comport with our own first-hand observations from geological outcrops within the Cradle of Humankind. Lichens do indeed grow on chert, dolomite and bone; they do sometimes overlie Mn deposits; and there is often a diffuse spot pattern in their surface distribution. But available evidence is not consistent with the rest of this line of reasoning. We here combine previously published taphonomic and geological data from the Dinaledi Chamber2, together with a fuller review of the literature and re-interpretation of photographic evidence presented by Thackeray1.
Abundant actualistic research documents mechanisms of Mn deposition on bone that have nothing to do with lichens.3 Mn and iron (Fe) deposition on bone in dark, wet, alkaline cave contexts like that found in the Dinaledi Chamber is normally a result of diagenetic recrystallisation of bone, incorporating trace elements from surrounding sediment and water via a diffusion-absorption process.4 In dolomitic caves, which have relatively high pH and oxidising redox, the alkali metals sodium and potassium and the alkaline earth metals calcium and magnesium are the most abundant soluble cations, while the transition metals copper, Fe and Mn are the least abundant, and the alkaline earth metals strontium and barium, and the transition metal zinc are of intermediate availability.5 Under these cave conditions, the stable forms of Fe and of Mn are hydrates and manganese oxide compounds, respectively. Precipitation of these highly insoluble hydrates and oxides varies with pH and tends to form crusts and coatings.6 Mn is mainly located on the surface and crack edges of fossil bones, whereas Fe shows deeper penetration into the bone matrix.7,8 Mn, which naturally occurs in groundwater, enters the bone environment as mobile Mn2+. In the presence of free O2, the oxidation of Mn2+ to Mn3+ and ultimately to insoluble Mn4+ is thermodynamically favourable, but proceeds extremely slowly unless mediated by microbial action, which is chemotrophic and does not require light.9,10 Oxidised Mn precipitates as a number of manganese oxides and hydroxides.11 Similarly, under the same conditions, mobile ferrous iron (Fe2+) approaching the bone may be oxidised to ferric iron (Fe3+) which rapidly precipitates as limonite (FeOOH.nH2O), which later usually undergoes transformation to goethite (FeOOH). Alternatively, pyrite (FeS2) is formed instead of haematite via precipitation of iron sulfide as decaying collagen releases sulfide ions into solution.12 Black or dark brown colour of fossil bones is a result of high levels of pyrite and manganese and iron oxide and hydroxide coatings. To summarise, processes that do not involve lichens are sufficient to explain the presence of Mn stains on fossil bone in contexts like that found within the Dinaledi Chamber today.
Mn, Fe and other mineral deposition occurs on a large fraction of the skeletal material from the Dinaledi Chamber in a variety of depositional forms. Thackeray1 chose to illustrate the pattern of Mn staining (specifically spotting) on only a single specimen. In his Figure 6 he provides a photograph of tibial specimen U.W. 101-996 (his figure caption misidentifies this specimen as U.W. 101-1070) with the figure legend:
Tibial shaft specimen U.W. 101-1070 [sic] H. naledi from the Dinaledi Chamber, with dotted coatings of manganese oxy-hydroxide. It is suggested that the black dots result, at least in part, from the growth of lichen as a bacterial-algal-fungal symbiont that includes a photobiont. The growth of lichen on such bone surfaces, even for a limited time, may have occurred in subdued, but essential lighting. Note the distribution [our emphasis] of manganese oxy-hydroxide, extending from a continuous matt to more dotted occurrences; this pattern is potentially analogous [our emphasis] to the dispersal of lichen from a central thallus.1
However, the distribution of Mn staining is not only on this one side of the specimen, but on all anatomical sides (anterior, posterior, medial and lateral), as shown in Figure 1. Such a distribution can also be seen on many specimens (for example, in Figure 2, specimen U.W. 101-1070, correctly attributed here). The distribution of Mn staining on the Dinaledi Chamber material is generally circumferential, occurring on multiple sides of bones. That distribution is not compatible with lichen growth in the present context, and there are significant reasons to reject the hypothesis that the Dinaledi Chamber is a secondary deposit.2,13

Instead, we have abundant evidence for Mn formation which occurs at the interface between soil matrix and free air. Our published analysis of the mineral staining on the Dinaledi Chamber skeletal material demonstrates not only the presence of black spots as noted by Thackeray1, but also tide marks of both Mn and Fe minerals on many of the specimens2. An example of these tide marks is shown in Figure 3. Lichens do not create such tide marks. These patterns form at the interface of soil and free air, and are a reflection of the function of the relative position of individual bones (or conjoin fragments) in the soil profile.2 Such patterns of mineral deposition are most parsimoniously explained by bone to soil matrix contact3,14 and are inconsistent with lichen growth.
So what evidence points Thackeray1 toward lichens? Thackeray's citation for lichen involvement is his own technical note15 in which he suggested a possible link between manganese dioxide staining and lichen growth on hominin crania from Sterkfontein and Swartkrans. This note, similar to his current contribution, posits merely that 'the spotty, discontinuous distribution of young lichen thalli appears to be analogous to the spotty distribution of MnO2 on at least some hominid crania'3(p.28). To support this hypothesis, it is incumbent upon Thackeray to quantify this resemblance in some way and show that it is unlikely to occur by chance. We have no opinion about whether lichen has deposited Mn on surface rocks such as those illustrated from Kromdraai (Figures 2 and 3 in Thackeray1). However, the lichen thalli illustrated in Thackeray's photographs do not have similar sizes to the Mn spots he has chosen to illustrate together with them. Nor do they have a similar density distribution. A very simple mode of quantification is to count the number of lichen thalli per square centimetre and the number of Mn spots; in the illustrated examples, these appear to be completely dissimilar. Further, Thackeray might have attempted to quantify the overall shape of lichen colonies and the overall shape of Mn staining. From his illustrations, we do not detect any such similarity except in the most general sense, in that they both have some dendritic portions, and thus any association with lichen may be a case of visual pareidolia.16 As such, any possible causal link awaits empirical testing and validation. Thackeray admits this is the case, indicating that '...long-term experimental work on lichen on bone substrates in the Cradle of Humankind is planned for a 10-year period, in and around cave environments'1(p.5).
In addition to lichen, Thackeray suggests1(p.5) that snails and beetles could indicate the presence of leaf litter (on which they feed), which would be found near the cave entrance and therefore in a situation with diffuse light. However, as noted by Dirks and colleagues13, snails have been recognised to colonise dark caves to considerable depth17, and are far from restricted in diet to just plant matter18. Gulella sp. and Euonyma varia (Connolly, 1910) occur in large numbers in the Rising Star Cave system, including in deep chambers in the dark zone, and may well be responsible for some of the bone surface modifications on the fossils. Subulinid snails (including Euonyma) are largely omnivorous and are thus not dependent on green plant material as a food source (Herbert D 2016, personal communication, January 28), while Gullela spp. are carnivorous and feed mostly on other snails.18
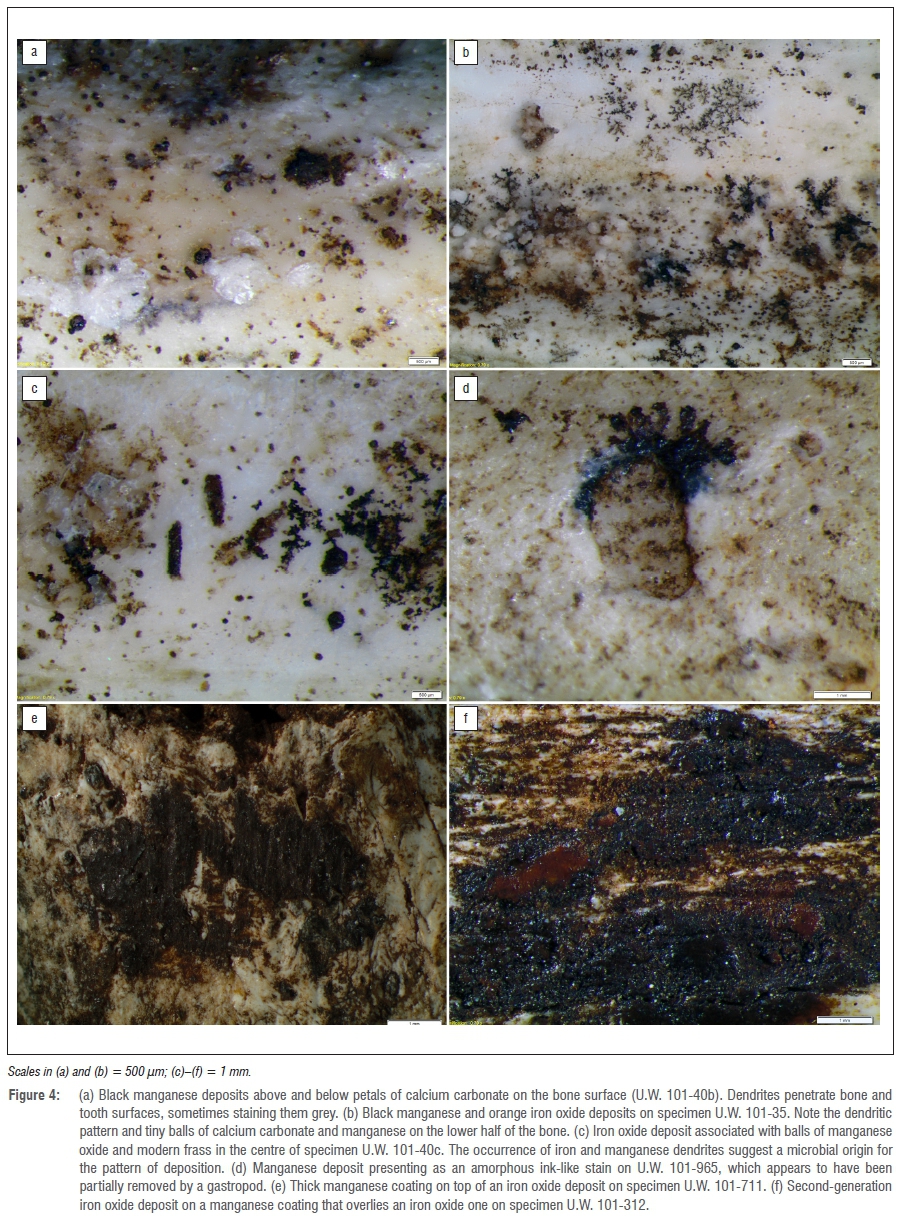
Thus the notion that snails are light-dwelling, leaf-litter feeders is incorrect, and bone surface modification by snails cannot be used to imply a close entrance to the surface or redeposition of the hominin fossil material from a location near light. Furthermore, much of the radula damage observed on the bones of H. naledi occurs after they were mineralised - therefore most invertebrate damage was probably produced inside the dark Dinaledi Chamber on bones already covered in coatings of manganese and iron oxide deposits (Figure 4).
Our work on the taphonomy and geology of the Dinaledi Chamber has been multidisciplinary, and we re-emphasise (as in Dirks et al.2) that any hypothesis must be consistent with all lines of evidence including geological, geochemical and taphonomic data. Thackeray proposes that an alternative opening to the Dinaledi Chamber, capable of transmitting diffuse light from the surface to this location more than 30 m underground, may have existed at the time the fossils were deposited. He cites the commentary of Val19, who likewise suggested that the fossils were once close to the surface in an environment with some light. Neither critic considers the strong geological or sedimentological evidence against such a scenario.13 As detailed in Dirks and colleagues2, the basic stratigraphic development of the Dinaledi Chamber comprises two facies subdivided into three stratigraphic units. Of these, hominin remains are found in Unit 2 remnants, with the bulk of the assemblage derived from Unit 3 - which accumulated along the floor of the chamber and is composed of largely unconsolidated sediment derived from weathering and erosion of Units 1 and 2.2 Sedimentological analyses indicate that the clay-rich sediments making up these units were derived from in-situ weathering, and from exogenous clays and silts, which entered the chamber through fractures that prevented passage of coarser-grained materials. Thus the infill of the Dinaledi Chamber is the end product of a series of filters or traps, which winnowed out all large-grained sediments or clastic material, en route to final deposition within the chamber. The sediment inside these fossil-bearing units of the Dinaledi Chamber is significantly different in particle size and composition2 from the neighbouring Dragon's Back Chamber, which is presently the only route from the Dinaledi Chamber toward the surface. If the fossils had been deposited at a time when there existed a larger opening into the Dinaledi Chamber, a secondary opening, or any substantially more open route to the surface, the sediment would not have these properties. Further, if the entrance to the Dinaledi Chamber had been illuminated and, therefore, accessible to surface fauna, or if there had not been special selection for only hominins, the bones of non-hominin fauna should be evident within the assemblage, in addition to other taphonomic markers of sub-aerial exposure, which are absent.2,13
In summary, we find that Thackeray's hypothesis of lichen deposition of Mn upon H. naledi fossil material is inconsistent with available evidence. Published evidence indicates that access to the Dinaledi Chamber was restricted to a single species of large-bodied animal, deposited over time, to the exclusion of all other animal forms during the period of the H. naledi depositional event.2 Any alternative model must allow fleshed and articulated remains to enter the chamber, including articulated hands and feet20,21, which are areas of anatomy that in articulation are unlikely to survive even short transport unless held together by soft tissues at the time of transport22,23. Furthermore, any other model must be restrictive enough that very limited externally derived sediments or organic material entered the chamber. Given these facts, we see nothing presented by Thackeray1, or other commentators19, that disproves the deliberate body disposal hypothesis we have put forward2.
References
1. Thackeray FJ. The possibility of lichen growth on bones of Homo naledi: Were they exposed to light? S Afr J Sci. 2016;112(7/8), Art. #a0167, 5 pages. http://dx.doi.org/10.17159/sajs.2016/a0167 [ Links ]
2. Dirks PHGM, Berger LR, Roberts EM, Kramers JD, Hawks J, Randolph-Quinney PS, et al. Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife. 2015;4, e09561, 37 pages. http://dx.doi.org/10.7554/eLife.09561 [ Links ]
3. Fernandez-Jalvo Y, Andrews P. Discolouration and staining. In: Fernandez Jalvo Y, Andrews P, editors. Atlas of taphonomic identifications. London: Springer; 2016. p. 155-68. http://dx.doi.org/10.1007/978-94-017-7432-1_5 [ Links ]
4. Millard AR, Hedges REM. A diffusion-adsorption model of uranium uptake by archaeological bone. Geochimic Cosmochim Ac. 1999;60:2139-2152. http://dx.doi.org/10.1016/0016-7037(96)00050-6 [ Links ]
5. Pate FD, Hutton JT, Norrish K. Ionic exchange between soil solution and bone: Toward a predictive model. Appl Geochem. 1989;4:303-316. http://dx.doi.org/10.1016/0883-2927(89)90034-6 [ Links ]
6. Karkanas P Bar-Yosef O, Goldberg P Weiner S. Diagenesis in prehistoric caves: The use of minerals that form in situ to assess the completeness of the archaeological record. J Archaeol Sci. 2000;27(10):915-929. http://dx.doi.org/10.1006/jasc.1999.0506 [ Links ]
7. Nhauro G. Characterization of the elemental deposits in fossils from the Cradle of Humankind in South Africa and modern bones from the same geological area. Johannesburg: University of the Witwatersrand; 2010. [ Links ]
8. López-González F, Grandal-d'Anglade A, Vidal-Romaní JR. Deciphering bone depositional sequences in caves through the study of manganese coatings. J Archaeol Sci. 2006;33(5):707-717. http://dx.doi.org/10.1016/j.jas.2005.10.006 [ Links ]
9. Diem D, Stumm W. Is dissolved Mn(II) being oxidized by O2 in absence of Mn-bacteria or surface catalysts? Geochim Cosmochim Ac. 1984;48:1571-1573. http://dx.doi.org/10.1016/0016-7037(84)90413-7 [ Links ]
10. Tebo BM, Johnson HA, McCarthy JK, Templeton AS. Geomicrobiology of manganese(II) oxidation. Trends Microbiol. 2005;13:421-128. http://dx.doi.org/10.1016/j.tim.2005.07.009 [ Links ]
11. Post JE. Manganese oxide minerals: Crystal structures and economic and environmental significance. Proc Natl Acad Sci USA. 1999;96:3447-3454. http://dx.doi.org/10.1073/pnas.96.7.3447 [ Links ]
12. Pfretzschner HU. Fossilization of Haversian bone in aquatic environments. CR Palevol. 2004;3:605-616. http://dx.doi.org/10.1016/j.crpv.2004.07.006 [ Links ]
13. Dirks PHGM, Berger LR, Hawks J, Randolph-Quinney PS, Backwell LR, Roberts EM. Comment on 'Deliberate body disposal by hominins in the Dinaledi Chamber, Cradle of Humankind, South Africa?' [J. Hum. Evol. 96(2016)145-148]. J Hum Evol. 2016;96:149-153. http://dx.doi.org/10.1016/j.jhevol.2016.04.007 [ Links ]
14. Pokines JT, Baker JE. Effects of burial environment on osseous remains. In: Pokines J, Symes S, editors. Manual of forensic taphonomy. Boca Raton, FL: CRC Press; 2013. p. 73-114. http://dx.doi.org/10.1201/b15424-6 [ Links ]
15. Thackeray JF, Sénégas F, Laden G. Manganese dioxide staining on hominid crania from Sterkfontein and Swartkrans: Possible associations with lichen. S Afr J Sci. 2005;101:28. [ Links ]
16. Liu J, Li J, Feng L, Li L, Tian J, Lee K. Seeing Jesus in toast: Neural and behavioural correlates of face pareidolia. Cortex. 2014;53:60-77. http://dx.doi.org/10.1016/j.cortex.2014.01.013 [ Links ]
17. Weigand A. New Zospeum species (Gastropoda, Ellobioidea, Carychiidae) from 980 m depth in the Lukina Jama-Trojama cave system (Velebit Mts., Croatia). Subterranean Biol. 2013;11:45-53. http://dx.doi.org/10.3897/subtbiol.11.5966 [ Links ]
18. Herbert D, Kilburn D. Field guide to the land snails and slugs of eastern South Africa. Durban: Natal Museum; 2004. [ Links ]
19. Val A. Deliberate body disposal by hominins in the Dinaledi Chamber, Cradle of Humankind, South Africa? J Hum Evol. 2016;96:145-148. http://dx.doi.org/10.1016/j.jhevol.2016.02.004 [ Links ]
20. Kivell TL, Deane AS, Tocheri MW, Orr CM, Schmid P, Hawks J, et al. The hand of Homo naledi. Nature Commun. 2015;6, Art. #8431, 9 pages. http://dx.doi.org/10.1038/ncomms9431 [ Links ]
21. Kruger A, Randolph-Quinney PS, Elliott M. Multimodal spatial mapping and visualisation of the Dinaledi Chamber and Rising Star Cave. S Afr J Sci. 2016;112(5/6), Art. #2016-0032, 11 pages. http://dx.doi.org/10.17159/sajs.2016/20160032 [ Links ]
22. Pokines JT. Faunal dispersal, reconcentration, and gnawing damage to bone in terrestrial environments. In: Pokines J, Symes S, editors. Manual of forensic taphonomy. Boca Raton, FL: CRC Press; 2013. p. 201-248. http://dx.doi.org/10.1201/b15424-10 [ Links ]
23. Galloway A. The process of decomposition: A model from the Arizona-Sonoran Desert. In: Haglund WD, Sorg MH, editors. Forensic taphonomy: The postmortem fate of human remains. London: CRC Press; 1997. p. 139-150. [ Links ]
 Correspondence:
Correspondence:
Patrick Randolph-Quinney
prandolph-quinney@uclan.ac.uk


{kind=link}
{kind=link}
{kind=link}